
Exogenously Applied GA3 Enhances Morphological Parameters of Tolerant and Sensitive Cyclamen persicum Genotypes under Ambient Temperature and Heat Stress Conditions




Abstract
:1. Introduction
2. Results
2.1. Variation in Germination Parameters of Heat-Resistant and Heat-Sensitive Cyclamen Genotypes under AT and HS
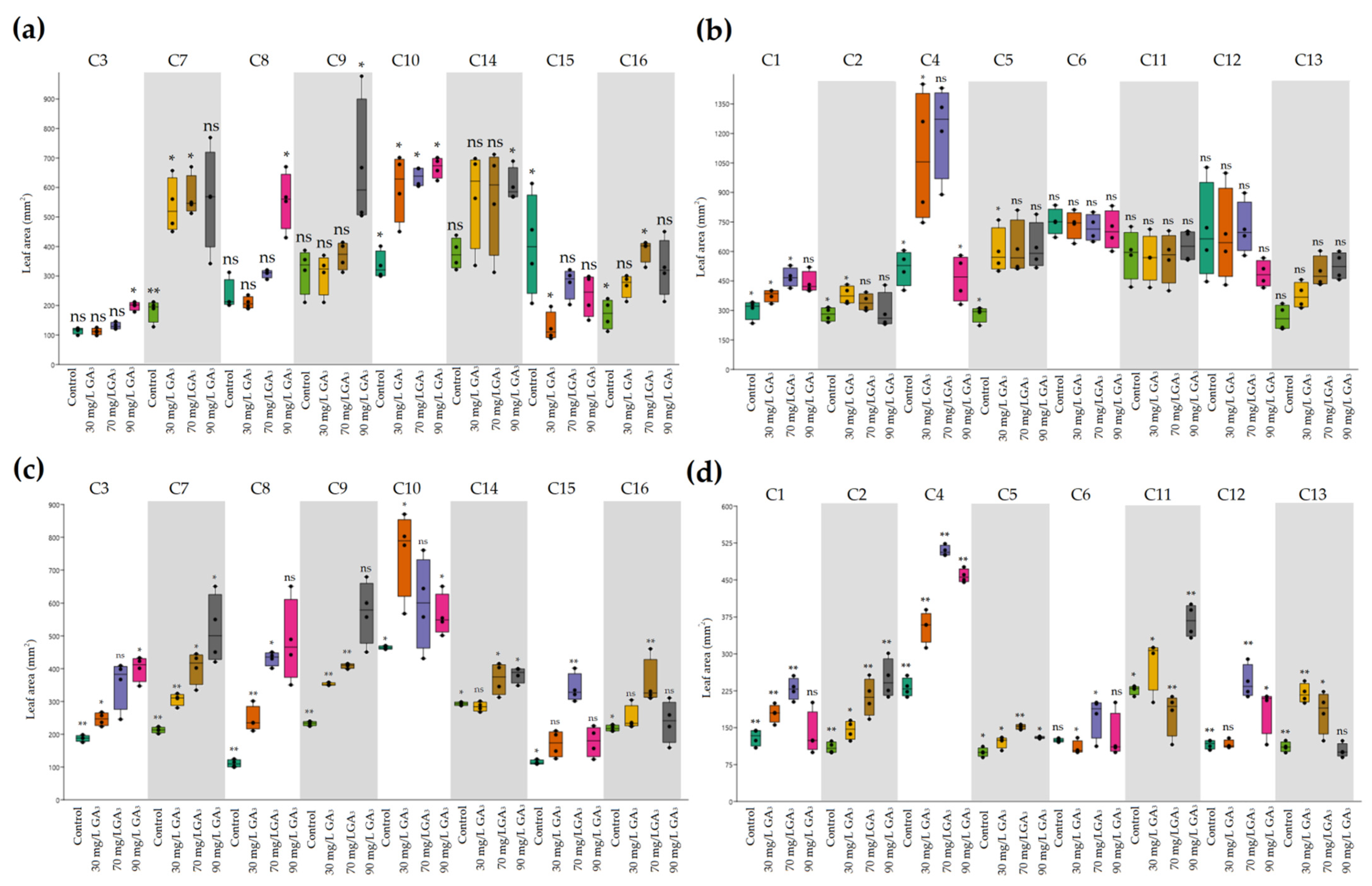
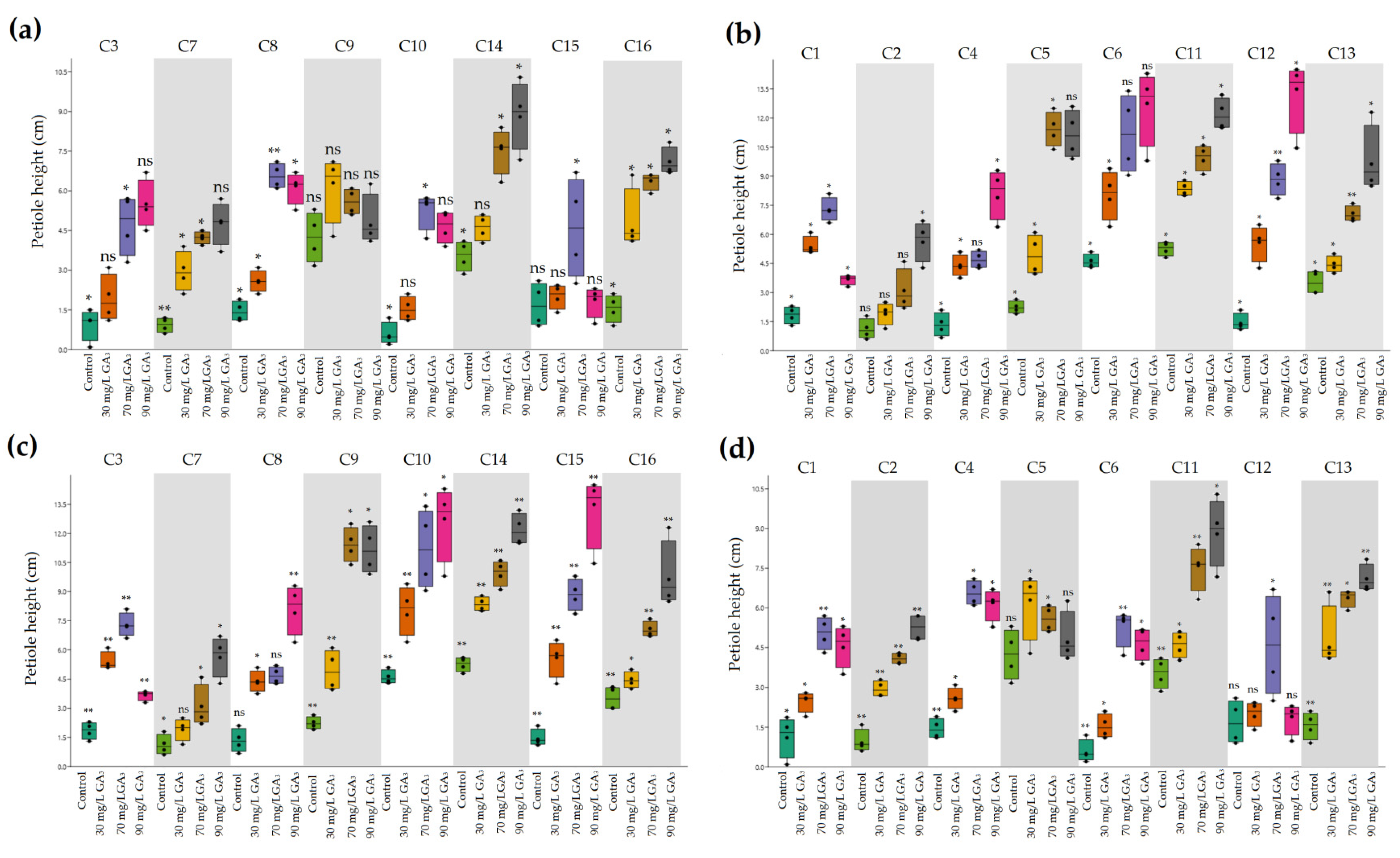
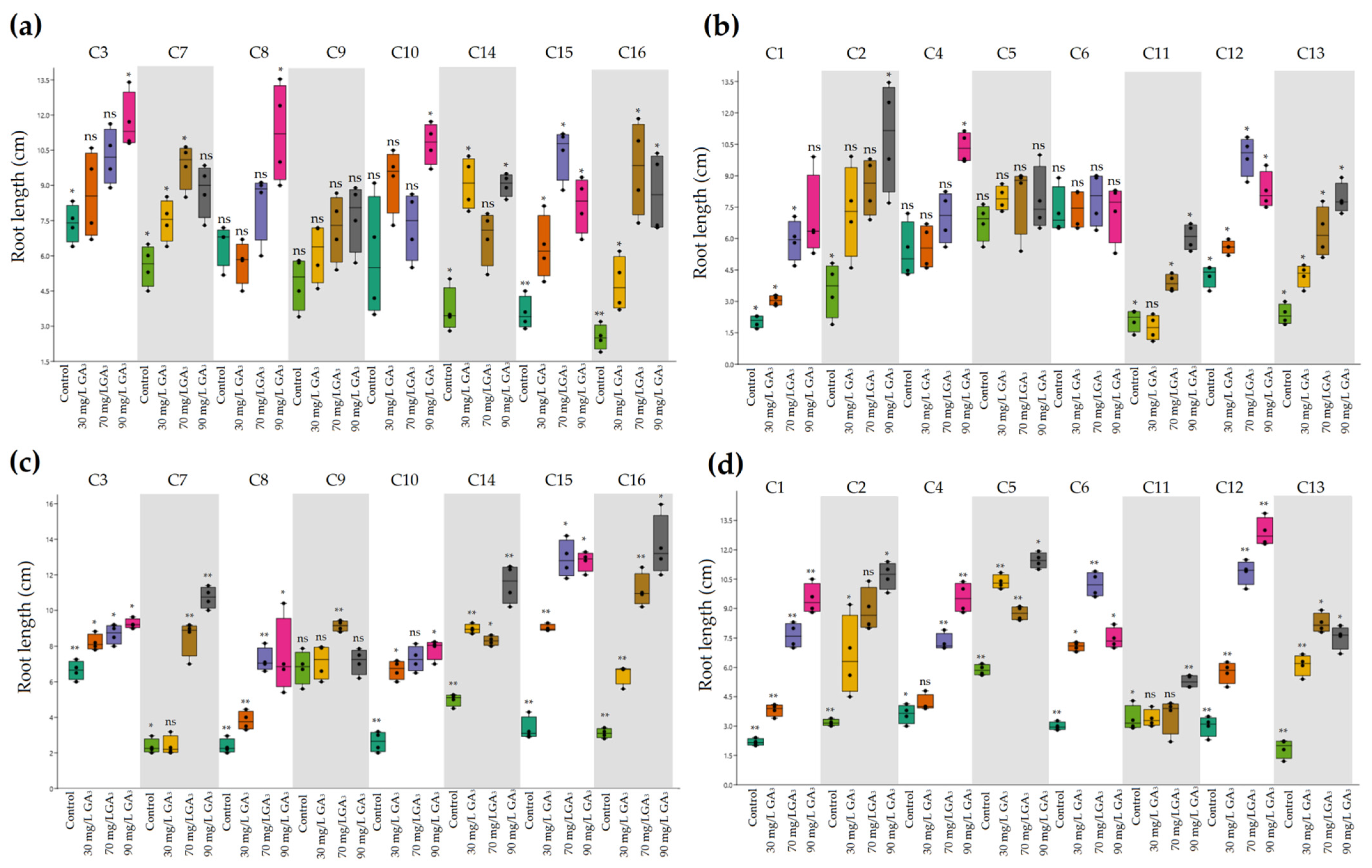
2.2. Variation in Plant Development of Heat-Resistant and Heat-Sensitive Cyclamen Genotypes under AT and HS
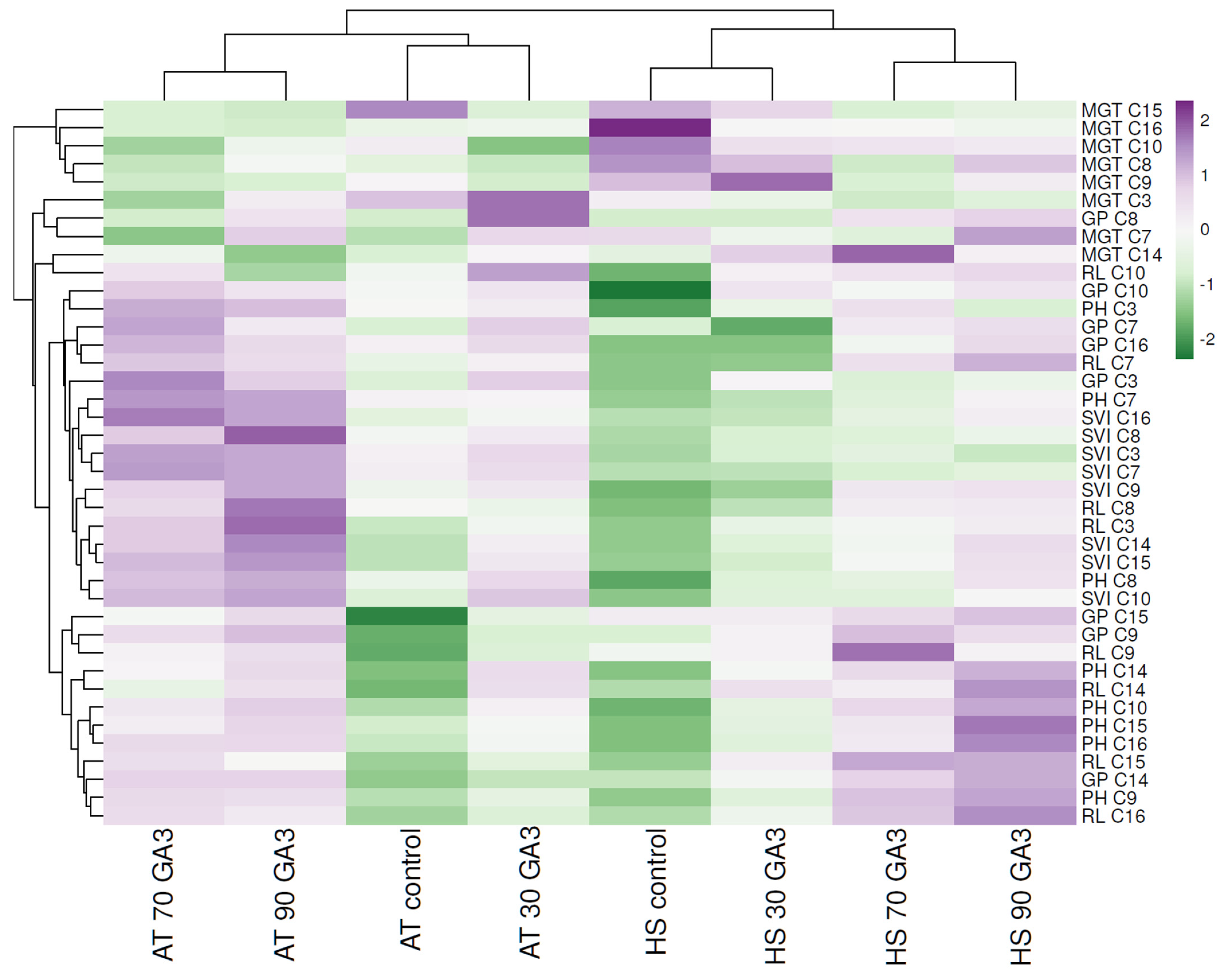
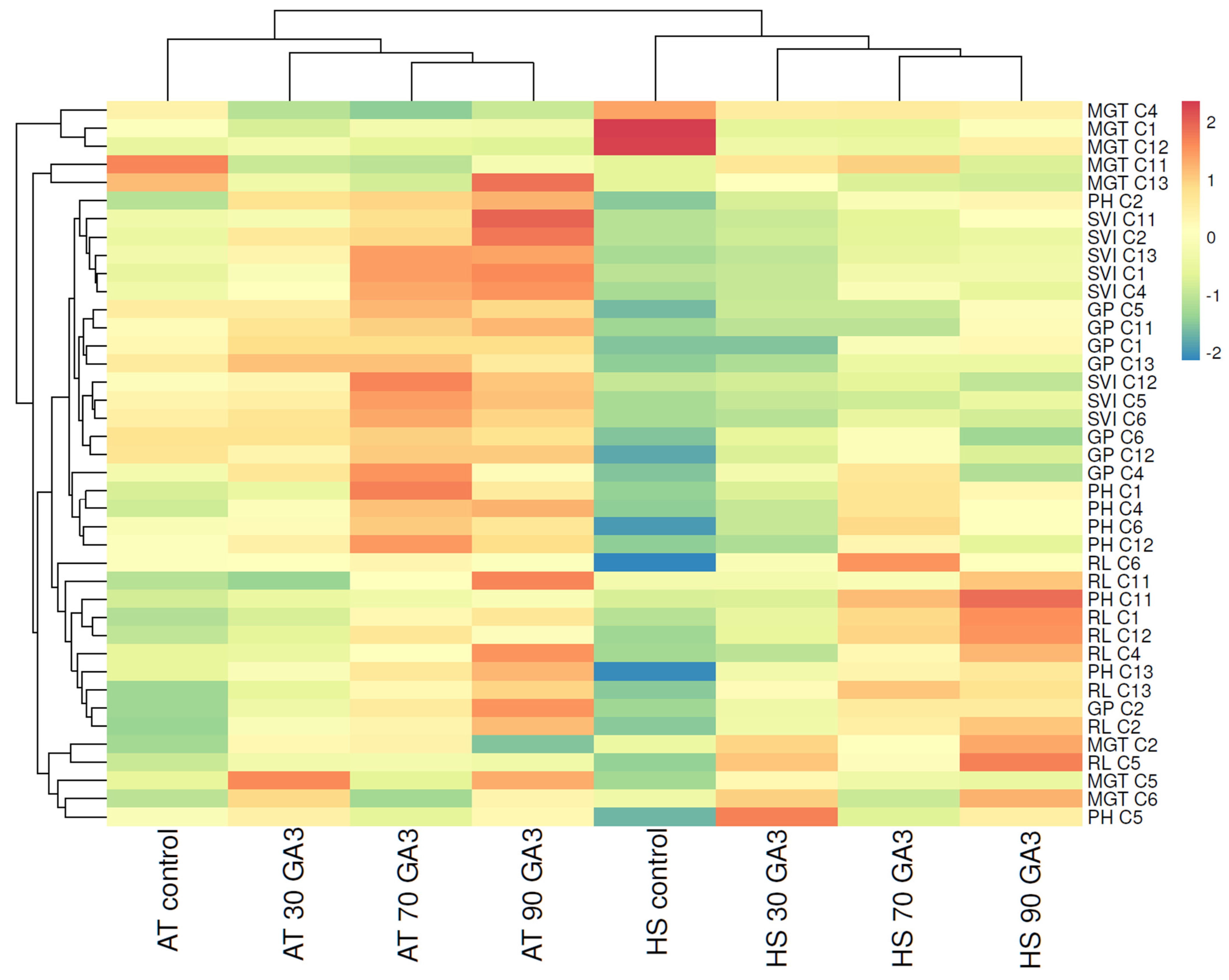
2.3. Hierarchical Clustering and Heat Mapping of Seed Germination and Plant Development Parameters of Heat-Tolerant and Heat-Sensitive Genotypes under Ambient Temperature and Heat Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Growth Regulator Treatment
4.3. Pot Experiment
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Venkateswarlu, B.; Shanker, A.K. Climate change and agriculture: Adaptation and mitigation stategies. Indian J. Agron. 2009, 54, 226–230. [Google Scholar]
- Eberius, M.; Lima-Guerra, J. High-Throughput Plant Phenotyping—Data Acquisition, Transformation, and Analysis. In Bioinformatics: Tools and Applications; Edwards, D., Stajich, J., Hansen, D., Eds.; Springer: New York, NY, USA, 2009; pp. 259–278. [Google Scholar]
- Wilson, G. Cyclamen (A Guide for Gardeners, Horticulturists and Botanists); Timber Press: London, UK, 2015. [Google Scholar]
- Cornea-Cipcigan, M.; Pamfil, D.; Sisea, C.R.; Mărgăoan, R. Gibberellic Acid Can Improve Seed Germination and Ornamental Quality of Selected Cyclamen Species Grown Under Short and Long Days. Agronomy 2020, 10, 516. [Google Scholar] [CrossRef] [Green Version]
- Karagur, E.R.; Ozay, C.; Mammadov, R.; Akca, H. Anti-invasive effect of Cyclamen pseudibericum extract on A549 non-small cell lung carcinoma cells via inhibition of ZEB1 mediated by miR-200c. J. Nat. Med. 2018, 72, 686–693. [Google Scholar] [CrossRef] [PubMed]
- El Hosry, L.; Di Giorgio, C.; Birer, C.; Habib, J.; Tueni, M.; Bun, S.-S.; Herbette, G.; De Meo, M.; Ollivier, E.; Elias, R. In vitro cytotoxic and anticlastogenic activities of saxifragifolin B and cyclamin isolated from Cyclamen persicum and Cyclamen libanoticum. Pharm. Biol. 2014, 52, 1134–1140. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G.; Mahomoodally, M.F.; Sinan, K.I.; Picot-Allain, M.C.N.; Yildiztugay, E.; Cziáky, Z.; Jekő, J.; Saleem, H.; Ahemad, N. Chemical characterization, antioxidant, enzyme inhibitory and cytotoxic properties of two geophytes: Crocus pallasii and Cyclamen cilicium. Food Res. Int. 2020, 133, 109129. [Google Scholar] [CrossRef]
- Cornea-Cipcigan, M.; Bunea, A.; Bouari, C.M.; Pamfil, D.; Páll, E.; Urcan, A.C.; Mărgăoan, R. Anthocyanins and Carotenoids Characterization in Flowers and Leaves of Cyclamen Genotypes Linked with Bioactivities Using Multivariate Analysis Techniques. Antioxidants 2022, 11, 1126. [Google Scholar] [CrossRef]
- Cordea, M.I.; Tiriplică, A. Influence of Pollen Germination Capacity on a Successful Artificial Hybridization in Cyclamen sp. Int. J. Innov. Appr. Agric. Res. 2019, 3, 53. [Google Scholar] [CrossRef]
- Cornea-Cipcigan, M.; Pamfil, D.; Sisea, C.R.; Gavriș, C.P.; da Graça Ribeiro Campos, M.; Margaoan, R. A review on Cyclamen species: Ttranscription factors vs. pharmacological effects. Acta Pol. Pharm.-Drug Res. 2019, 76, 919–938. [Google Scholar] [CrossRef]
- Breeding, S. Growing Cyclamen under High Light Levels. Available online: https://schoneveld-breeding.com/en/growing-cyclamen-under-high-light-levels/ (accessed on 16 June 2022).
- Savaedi, Z.; Parmoon, G.; Moosavi, S.A.; Bakhshande, A. The role of light and Gibberellic Acid on cardinal temperatures and thermal time required for germination of Charnushka (Nigella sativa) seed. Ind. Crops Prod. 2019, 132, 140–149. [Google Scholar] [CrossRef]
- Jha, U.C.; Nayyar, H.; Siddique, K.H.M. Role of Phytohormones in Regulating Heat Stress Acclimation in Agricultural Crops. J. Plant Growth Regul. 2022, 41, 1041–1064. [Google Scholar] [CrossRef]
- Wang, H.-Q.; Liu, P.; Zhang, J.-W.; Zhao, B.; Ren, B.-Z. Endogenous Hormones Inhibit Differentiation of Young Ears in Maize (Zea mays L.) Under Heat Stress. Front. Plant Sci. 2020, 11, 533046. [Google Scholar] [CrossRef]
- Ma, H.-Y.; Zhao, D.-D.; Ning, Q.-R.; Wei, J.-P.; Li, Y.; Wang, M.-M.; Liu, X.-L.; Jiang, C.-J.; Liang, Z.-W. A Multi-year Beneficial Effect of Seed Priming with Gibberellic Acid-3 (GA3) on Plant Growth and Production in a Perennial Grass, Leymus chinensis. Sci. Rep. 2018, 8, 13214. [Google Scholar] [CrossRef] [PubMed]
- Tsegay, B.A.; Andargie, M. Seed Priming with Gibberellic Acid (GA3) Alleviates Salinity Induced Inhibition of Germination and Seedling Growth of Zea mays L., Pisum sativum Var. abyssinicum A. Braun and Lathyrus sativus L. J. Crop. Sci. Biotechnol. 2018, 21, 261–267. [Google Scholar] [CrossRef]
- Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System. Agronomy 2019, 9, 382. [Google Scholar] [CrossRef] [Green Version]
- Muniandi, S.K.M.; Hossain, M.A.; Abdullah, M.P.; Ab Shukor, N.A. Gibberellic acid (GA3) affects growth and development of some selected kenaf (Hibiscus cannabinus L.) cultivars. Ind. Crops Prod. 2018, 118, 180–187. [Google Scholar] [CrossRef]
- Garmendia, A.; Beltrán, R.; Zornoza, C.; García-Breijo, F.J.; Reig, J.; Merle, H. Gibberellic acid in Citrus spp. flowering and fruiting: A systematic review. PLoS ONE 2019, 14, e0223147. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Deng, Y.; Wang, H.; Gao, R.; Stephen, G.u.K.; Chen, S.; Jiang, J.; Chen, F. Gibberellic Acid Signaling Is Required to Induce Flowering of Chrysanthemums Grown under Both Short and Long Days. Int. J. Mol. Sci. 2017, 18, 1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, B.A.; Dole, J.M.; McCall, I. Gibberellic Acid Shows Promise for Promoting Flower Stem Length in Four Field-grown Cut Flowers. HortTechnology Hortte 2016, 26, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Toscano, S.; Trivellini, A.; Ferrante, A.; Romano, D. Physiological mechanisms for delaying the leaf yellowing of potted geranium plants. Sci. Hortic. 2018, 242, 146–154. [Google Scholar] [CrossRef]
- Khan, M.N.; Khan, Z.; Luo, T.; Liu, J.; Rizwan, M.; Zhang, J.; Xu, Z.; Wu, H.; Hu, L. Seed priming with gibberellic acid and melatonin in rapeseed: Consequences for improving yield and seed quality under drought and non-stress conditions. Ind. Crops Prod. 2020, 156, 112850. [Google Scholar] [CrossRef]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed Priming: New Comprehensive Approaches for an Old Empirical Technique; InTech: London, UK, 2016. [Google Scholar]
- Heydariyan, M.; Basirani, N.; Sharifi-Rad, M.; Khmmari, I.; Poor, S.R. Effect of seed priming on germination and seedling growth of the caper (Capparis spinosa) under drought stress. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 2381–2389. [Google Scholar]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Aguilar-Trigueros, C.A.; Flaig, I.C.; Rillig, M.C. Root trait responses to drought are more heterogeneous than leaf trait responses. Funct. Ecol. 2020, 34, 2224–2235. [Google Scholar] [CrossRef]
- Teixeira, E.I.; Fischer, G.; van Velthuizen, H.; Walter, C.; Ewert, F. Global hot-spots of heat stress on agricultural crops due to climate change. Agric. For. Meteorol. 2013, 170, 206–215. [Google Scholar] [CrossRef]
- Hemantaranjan, A.; Bhanu, A.N.; Singh, M.N.; Yadav, D.K.; Patel, P.K.; Singh, R.; Katiyar, D. Heat stress responses and thermotolerance. Adv. Plants Agric. Res. 2014, 1, 00012. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Al-Whaibi, M.H.; Grover, A.; Ali, H.M.; Al-Wahibi, M.S. Morphological and physiological characterization of different genotypes of faba bean under heat stress. Saudi J. Biol. Sci. 2015, 22, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Pallaoro, D.S.; Avelino, A.C.D.; Camili, E.C.; Guimarães, S.C.; Albuquerque, M.C.D.F. Priming corn seeds with plant growth regulator. J. Seed Sci. 2016, 38, 227–232. [Google Scholar] [CrossRef]
- Ortega-Baes, P.; Rojas-Aréchiga, M. Seed germination of Trichocereus terscheckii (Cactaceae): Light, temperature and gibberellic acid effects. J. Arid. Environ. 2007, 69, 169–176. [Google Scholar] [CrossRef]
- Vieira, A.R.; Vieira, M.D.G.G.C.; Fraga, A.C.; Oliveira, J.A.; Santos, C.D.D. Action of gibberellic acid (GA3) on dormancy and activity of alpha-amylase in rice seeds. Rev. Bras. Sementes 2002, 24, 43–48. [Google Scholar]
- Sappalani, G.I.; Cabahug, L.M.; Valleser, V.C. Impact of gibberellic acid and organic growth media on seed germination and seedling development of rubber (Hevea brasiliensis). Int. J. Hortic. Sci. Technol. 2021, 8, 165–174. [Google Scholar]
- Szekely-Varga, Z.; Kentelky, E.; Cantor, M. Effect of Gibberellic Acid on the Seed Germination of Lavandula angustifolia Mill. RJH 2021, 2, 169–176. [Google Scholar] [CrossRef]
- Islam, M.M.; Rengel, Z.; Storer, P.; Siddique, K.H.M.; Solaiman, Z.M. Industrial Hemp (Cannabis sativa L.) Varieties and Seed Pre-Treatments Affect Seed Germination and Early Growth of Seedlings. Agronomy 2022, 12, 6. [Google Scholar] [CrossRef]
- Li, Z.; Lu, G.Y.; Zhang, X.K.; Zou, C.S.; Cheng, Y.; Zheng, P.Y. Improving drought tolerance of germinating seeds by exogenous application of gibberellic acid (GA3) in rapeseed (Brassica napus L.). Seed Sci. Technol. 2010, 38, 432–440. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Rauf, F.; Tania, S.S.; Khatun, M. Seed priming methods: Application in field crops and future perspectives. Asian J. Res. Crop. Sci. 2020, 5, 8–19. [Google Scholar] [CrossRef]
- Nimir, N.E.A.; Lu, S.; Zhou, G.; Guo, W.; Ma, B.; Wang, Y. Comparative effects of gibberellic acid, kinetin and salicylic acid on emergence, seedling growth and the antioxidant defence system of sweet sorghum (Sorghum bicolor) under salinity and temperature stresses. Crop Pasture Sci. 2015, 66, 145–157. [Google Scholar] [CrossRef]
- Khan, A.; Bilal, S.; Khan, A.L.; Imran, M.; Shahzad, R.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Azhri, M.; Mohanta, T.K.; Lee, I.-J. Silicon and Gibberellins: Synergistic Function in Harnessing ABA Signaling and Heat Stress Tolerance in Date Palm (Phoenix dactylifera L.). Plants 2020, 9, 620. [Google Scholar] [CrossRef]
- Alonso-Ramírez, A.; Rodríguez, D.; Reyes, D.; Jiménez, J.S.A.; Nicolás, G.; López-Climent, M.A.; Gómez-Cadenas, A.; Nicolás, C. Evidence for a Role of Gibberellins in Salicylic Acid-Modulated Early Plant Responses to Abiotic Stress in Arabidopsis Seeds. Plant Physiol. 2009, 150, 1335–1344. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Jannasch, A.H.; Jiang, Y. Growth and Hormone Alterations in Response to Heat Stress in Perennial Ryegrass Accessions Differing in Heat Tolerance. J. Plant Growth Regul. 2020, 39, 1022–1029. [Google Scholar] [CrossRef]
- Neveur, N.; Corbineau, F.; Côme, D. Some characteristics of Cyclamen persicum L. seed germination. J. Hortic. Sci. 1986, 61, 379–387. [Google Scholar] [CrossRef]
- Heydecker, W.; Wainwright, H. More rapid and uniform germination of Cyclamen persicum L. Sci. Hortic. 1976, 5, 183–189. [Google Scholar] [CrossRef]
- Corbineau, F.; Neveur, N.; Côme, D. Seed Germination and Seedling Development in Cyclamen persicum. Ann. Bot. 1989, 63, 87–96. [Google Scholar] [CrossRef]
- Fagge, A.A.; Manga, A.A. Effect of sowing media and gibberellic acid on the growth and seedling establishment of Bougainvillea glabra, Ixora coccinea and Rosa chinensis. 2. Root Characters. Bayero J. Pure Appl. Sci. 2011, 4, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Arteca, R.N.; Schlagnhaufer, C.D.; Arteca, J.M. Root applications of gibberellic acid enhance growth of seven Pelargonium cultivars. HortScience 1991, 26, 555–556. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Deng, H.; Zhang, X.; Yu, X.; Li, Y. Gibberellin Is Involved in Inhibition of Cucumber Growth and Nitrogen Uptake at Suboptimal Root-Zone Temperatures. PLoS ONE 2016, 11, e0156188. [Google Scholar] [CrossRef] [PubMed]
- Stavang, J.A.; Gallego-Bartolomé, J.; Gómez, M.D.; Yoshida, S.; Asami, T.; Olsen, J.E.; García-Martínez, J.L.; Alabadí, D.; Blázquez, M.A. Hormonal regulation of temperature-induced growth in Arabidopsis. Plant J. 2009, 60, 589–601. [Google Scholar] [CrossRef]
- Lu, L.; Liu, H.; Wu, Y.; Yan, G. Wheat genotypes tolerant to heat at seedling stage tend to be also tolerant at adult stage: The possibility of early selection for heat tolerance breeding. Crop J. 2022. [Google Scholar] [CrossRef]
- Kader, M.A. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proc. R. Soc. N. S. W. 2005, 138, 65–75. [Google Scholar]
- Al-Ansari, F.; Ksiksi, T. A Quantitative Assessment of Germination Parameters: The Case of Crotalaria Persica and Tephrosia apollinea. Open Ecol. J. 2016, 9, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.H.; Roberts, E.H. Towards a Rational Basis for Testing Seed Quality; Butterworths: London, UK, 1980. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Relationship Between Decarboxylation of Glutamic Acid and Vigor in Soybean Seed. Crop Sci. 1973, 13, 227–232. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Genotype | GA3 Concentrations Treatment under AT | GA3 Concentrations Treatment under HS | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 mg/L GA3 | 30 mg/L GA3 | 70 mg/L GA3 | 90 mg/L GA3 | 0 mg/L | 30 mg/L | 70 mg/L | 90 mg/L GA3 | ||
| GA3 | GA3 | GA3 | |||||||
| Germination percentage (%) | Heat-resistant genotypes | ||||||||
| C3 | 80.0 ± 8.0 b | 93.3 ± 9.1 a | 100.0 ± 10.0 a | 93.3 ± 9.4 a | 73.4 ± 3.3 c | 86.7 ± 2.1 b | 80.0 ± 2.0 c | 83.3 ± 3.4 c | |
| C7 | 73.3 ± 9.1 b | 93.3 ± 9.5 ab | 100.0 ± 10.2 a | 86.7 ± 8.8 ab | 73.3 ± 5.1 c | 60.0 ± 1.8 d | 86.7 ± 3.1 de | 90.0 ± 1.8 b | |
| C8 | 86.7 ± 8.3 a | 100.0 ± 8.9 a | 86.7 ± 8.0 a | 93.3 ± 8.8 a | 86.7 ± 4.5 a | 86.7 ± 5.7 b | 93.3 ± 6.0 ab | 95.0 ± 3.4 ab | |
| C9 | 60.0 ± 8.0 b | 73.3 ± 8.8 ab | 93.3 ± 8.3 ab | 100.0 ± 9.7 a | 73.3 ± 4.6 c | 86.7 ± 2.9 b | 100.0 ± 3.3 a | 93.3 ± 1.7 b | |
| C10 | 86.7 ± 7.9 b | 93.3 ± 9.0 ab | 100.0 ± 8.9 a | 93.3 ± 9.5 ab | 53.4 ± 6.7 d | 93.3 ± 6.6 a | 86.7 ± 2.8 d | 93.3 ± 2.5 b | |
| C14 | 53.3 ± 5.3 c | 60.0 ± 6.0 b | 86.7 ± 7.8 a | 86.7 ± 7.8 a | 60.0 ± 5.5 d | 73.3 ± 2.0 c | 86.7 ± 5.8 de | 93.3 ± 3.3 ab | |
| C15 | 40.0 ± 4.1 c | 73.3 ± 7.3 b | 80.0 ± 7.9 b | 93.3 ± 9.2 a | 86.7 ± 4.5 a | 86.7 ± 6.0 ab | 93.3 ± 5.2 ab | 100.0 ± 4.0 a | |
| C16 | 90.0 ± 8.4 b | 95.0 ± 9.5 ab | 100.0 ± 9.7 a | 95.0 ± 9.1 ab | 73.3 ± 4.4 c | 73.3 ± 3.5 c | 86.7 ± 2.7 d | 95.0 ± 1.1 b | |
| Heat-sensitive genotypes | |||||||||
| C1 | 80.0 ± 7.9 a | 93.3 ± 8.6 a | 93.3 ± 8.8 a | 93.3 ± 9.4 a | 40.0 ± 3.8 e | 40.0 ± 2.5 f | 73.3 ± 0.8 f | 80.0 ± 2.4 c | |
| C2 | 80.0 ± 7.6 b | 86.7 ± 8.8 ab | 93.3 ± 9.3 a | 100.0 ± 9.5 a | 80.0 ± 0.6 b | 86.7 ± 2.6 b | 93.3 ± 1.5 b | 93.3 ± 2.1 b | |
| C4 | 73.3 ± 7.4 b | 86.7 ± 8.3 b | 100.0 ± 9.6 a | 80.0 ± 7.8 b | 53.4 ± 2.4 d | 73.3 ± 5.3 c | 86.7 ± 1.6 d | 60.0 ± 2.8 e | |
| C5 | 80.0 ± 7.6 b | 80.0 ± 7.3 b | 93.3 ± 9.7 a | 86.7 ± 8.9 ab | 40.0 ± 5.6 e | 53.3 ± 2.3 e | 53.3 ± 1.9 h | 73.3 ± 3.9 d | |
| C6 | 93.3 ± 9.0 a | 93.3 ± 8.7 a | 100.0 ± 9.9 a | 93.3 ± 8.7 a | 33.3 ± 3.2 ef | 60.0 ± 3.3 d | 73.3 ± 2.9 f | 40.0 ± 1.7 g | |
| C11 | 73.3 ± 6.8 b | 86.7 ± 8.0 ab | 93.3 ± 8.9 a | 100.0 ± 9.9 a | 33.3 ± 5.8 ef | 40.0 ± 3.3 f | 40.0 ± 1.6 i | 73.3 ± 7.1 d | |
| C12 | 86.7 ± 8.2 a | 80.0 ± 7.9 a | 93.3 ± 8.9 a | 93.3 ± 9.0 a | 40.0 ± 7.7 e | 60.0 ± 4.9 d | 73.3 ± 3.1 f | 60.0 ± 8.0 ef | |
| C13 | 86.7 ± 9.2 b | 100.0 ± 9.9 a | 100.0 ± 10.4 a | 86.7 ± 8.4 b | 33.3 ± 7.6 f | 40.0 ± 5.6 f | 60.0 ± 3.4 g | 60.0 ± 5.6 ef | |
| MGT | Heat-resistant genotypes | ||||||||
| C3 | 13.90 ± 5.7 a | 17.64 ± 7.2 a | 3.24 ± 2.0 b | 10.49 ± 6.0 a | 10.31 ± 3.49 e | 7.50 ± 5.40 g | 5.088 ± 2.34 ef | 6.42 ± 3.84 e | |
| C7 | 4.02 ± 2.1 ab | 8.70 ± 6.1 a | 3.03 ± 1.5 b | 8.98 ± 5.9 a | 8.48 ± 0.91 f | 6.13 ± 3.06 gh | 5.35 ± 2.33 e | 10.34 ± 6.56 d | |
| C8 | 6.18 ± 3.3 a | 4.25 ± 2.2 a | 3.90 ± 2.0 a | 8.66 ± 7.7 a | 16.91 ± 2.66 d | 14.33 ± 2.86 cd | 4.62 ± 2.08 f | 13.85 ± 1.87 b | |
| C9 | 11.44 ± 5.0 a | 3.71 ± 2.0 b | 3.39 ± 1.9 b | 4.32 ± 2.2 b | 19.35 ± 5.48 c | 26.24 ± 1.90 a | 4.68 ± 2.49 f | 12.98 ± 3.71 c | |
| C10 | 11.57 ± 6.1 a | 2.34 ± 0.9 b | 3.70 ± 2.0 ab | 9.00 ± 7.9 a | 18.72 ± 4.46 cd | 12.86 ± 7.89 de | 12.28 ± 1.69 c | 11.89 ± 3.94 d | |
| C14 | 7.01 ± 3.1 a | 12.19 ± 7.3 a | 9.89 ± 6.2 a | 2.65 ± 0.3 b | 8.42 ± 3.53 f | 16.77 ± 3.52 b | 23.51 ± 3.16 a | 12.69 ± 2.90 c | |
| C15 | 19.95 ± 6.2 a | 4.82 ±1.9 b | 4.25 ± 1.6 b | 3.72 ± 1.6 b | 17.10 ± 2.08 d | 14.30 ± 1.09 cd | 4.54 ± 1.99 g | 6.47 ± 2.66 e | |
| C16 | 4.26 ± 2.1 a | 5.14 ± 2.6 a | 2.01 ± 0.7 b | 1.76 ± 0.6 b | 19.39 ± 3.95 c | 6.47 ± 3.00 gh | 6.05 ± 2.64 e | 5.12 ± 2.14 f | |
| Heat-sensitive genotypes | |||||||||
| C1 | 13.43 ± 7.4 a | 2.49 ± 0.7 a | 10.10 ± 6.0 a | 9.06 ± 4.8 a | 44.89 ± 4.94 a | 5.21 ± 1.81 h | 4.31 ± 3.01 g | 12.98 ± 5.87 c | |
| C2 | 4.77 ± 2.1 a | 10.43 ± 4.3 a | 10.91 ± 4.6 a | 3.74 ± 1.6 a | 7.98 ± 4.88 g | 13.069 ± 3.81 de | 9.944 ± 3.30 d | 14.61 ± 2.48 a | |
| C4 | 11.55 ± 6.4 a | 3.69 ± 1.5 b | 1.64 ± 0.4 c | 4.54 ± 1.7 b | 17.05 ± 2.62 d | 12.33 ± 2.67 e | 12.63 ± 4.72 c | 11.80 ± 1.70 d | |
| C5 | 10.37 ± 4.8 ab | 22.73 ± 12.5 a | 9.63 ± 4.4 b | 21.09 ± 12.7 a | 6.01 ± 3.46 h | 14.95 ± 1.87 c | 11.35 ± 3.74 d | 10.82 ± 3.57 d | |
| C6 | 3.86 ± 2.3 ab | 11.01 ± 8.0 a | 2.90 ± 1.9 b | 9.01 ± 5.7 a | 6.27 ± 3.02 h | 11.34 ± 4.68 e | 4.33 ± 2.19 g | 12.46 ± 1.67 c | |
| C11 | 20.54 ± 12.1 a | 3.94 ± 1.6 b | 3.18 ± 0.6 b | 8.78 ± 3.1 b | 6.08 ± 3.18 h | 14.02 ± 3.11 d | 16.23 ± 1.13 b | 5.11 ± 2.13 f | |
| C12 | 4.61 ± 1.6 a | 7.27 ± 3.1 a | 3.36 ± 1.5 ab | 3.18 ± 1.5 b | 32.35 ± 1.11 b | 5.88 ± 3.19 h | 5.32 ± 2.49 e | 14.45 ± 6.95 a | |
| C13 | 17.87 ± 9.1 a | 7.60 ± 3.6 b | 4.18 ± 3.4 a | 22.78 ± 13.8 a | 5.84 ± 3.19 h | 10.60 ± 4.98 ef | 4.95 ± 2.49 ef | 4.28 ± 1.96 f | |
| Parameters | Genotype | GA3 Concentrations Treatment under AT | GA3 Concentrations Treatment under HS | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 mg/L GA3 | 30 mg/L GA3 | 70 mg/L GA3 | 90 mg/L GA3 | 0 mg/L GA3 | 30 mg/L GA3 | 70 mg/L GA3 | 90 mg/L GA3 | ||
| Heat-resistant genotypes | |||||||||
| Plant height (cm) | C3 | 7.39 ± 0.2 b | 8.10 ± 0.5 ab | 10.74 ± 1.1 a | 10.17 ± 0.5 a | 2.31 ± 0.0 ef | 6.46 ± 1.1 cd | 8.78 ± 1.2 d | 5.41 ± 0.2 f |
| C7 | 7.04 ± 0.7 c | 6.83 ± 0.9 c | 12.11 ± 0.9 a | 11.44 + 0.7 b | 1.64 ± 0.1 g | 2.85 ± 0.5 h | 4.41 ± 0.1 g | 7.01 + 0.3 de | |
| C8 | 7.39 ± 0.4 b | 11.15 ± 1.0 a | 12.12 ± 0.7 a | 12.99 + 1.1 a | 1.83 ± 0.0 f | 6.17 ± 0.9 de | 6.89 ± 0.2 e | 10.22 + 1.2 c | |
| C9 | 5.24 ± 0.1 c | 7.75 ± 0.6 b | 12.15 ± 0.8 a | 11.82 + 0.1 a | 3.94 ± 0.3 c | 7.04 ± 0.4 c | 13.54 ± 1.7 ab | 14.89 + 0.4 b | |
| C10 | 7.93 ± 0.2 c | 13.25 ± 0.5 b | 14.06 ± 0.2 b | 16.07 ± 0.6 a | 5.52 ± 0.0 b | 10.77 ± 0.7 a | 15.43 ± 1.1 a | 17.98 ± 0.7 a | |
| C14 | 6.08 ± 0.8 c | 12.63 ± 0.1 b | 11.02 ± 0.4 d | 12.76 ± 0.3 a | 6.27 ± 0.4 a | 10.59 ± 0.4 a | 12.79 ± 0.4 b | 14.29 ± 0.8 b | |
| C15 | 5.74 ± 0.3 d | 8.88 ± 0.5 c | 9.94 ± 0.7 b | 12.07 ± 1.2 a | 2.94 ± 0.2 d | 7.14 ± 0.6 c | 10.55 ± 1.5 c | 16.17 ± 1.8 a | |
| C16 | 5.72 ± 0.5 c | 7.82 ± 0.3 b | 9.94 ± 0.2 a | 10.06 ± 1.2 a | 4.12 ± 0.1 c | 6.64 ± 0.3 cd | 8.96 ± 0.1 d | 12.35 ± 1.1 bc | |
| Heat-sensitive genotypes | |||||||||
| C1 | 3.24 ± 0.4 d | 3.98 ± 0.8 c | 9.76 ± 1.5 a | 6.73 ± 0.8 b | 1.56 ± 0.2 g | 3.29 ± 0.3 fg | 7.19 ± 1.1 e | 5.90 ± 0.7 f | |
| C2 | 2.51 ± 0.1 c | 7.92 ± 0.4 b | 8.47 ± 0.8 a | 9.43 ± 1.0 a | 1.31 ± 0.3 h | 3.45 ± 0.4 fg | 5.43 ± 0.8 f | 6.60 ± 3.5 e | |
| C4 | 4.10 ± 0.3 c | 6.70 ± 2.1 b | 10.08 ± 0.9 a | 10.50 + 1.1 a | 2.27 ± 0.2 f | 3.81 ± 0.8 f | 8.85 ± 0.6 d | 7.10 + 0.7 de | |
| C5 | 6.96 ± 0.2 c | 7.18 ± 0.3 a | 6.72 ± 0.4 d | 7.09 + 0.5 b | 6.24 ± 0.4 a | 7.72 ± 0.3 b | 6.68 ± 0.5 e | 7.19 + 0.5 d | |
| C6 | 5.72 ± 0.4 d | 6.55 ± 0.3 c | 8.47 ± 0.9 a | 7.58 + 0.6 b | 1.23 ± 0.2 h | 3.62 ± 0.3 f | 8.22 ± 0.3 d | 6.24 + 2.4 e | |
| C11 | 5.60 ± 0.2 a | 6.31 ± 0.7 a | 6.54 ± 0.4 a | 7.02 ± 0.8 a | 5.74 ± 0.2 b | 5.76 ± 0.2 e | 9.44 ± 0.9 c | 10.80 ± 0.3 c | |
| C12 | 5.73 ± 0.4 c | 6.60 ± 0.5 b | 8.89 ± 1.0 a | 7.50 ± 0.7 a | 2.56 ± 0.1 d | 3.19 ± 0.3 gh | 6.37 ± 1.1 ef | 4.44 ± 0.2 g | |
| C13 | 5.59 ± 0.6 d | 6.75 ± 0.5 c | 8.24 ± 0.4 b | 9.51 ± 0.8 a | 2.22 ± 0.3 f | 6.19 ± 0.5 d | 7.59 ± 0.8 de | 8.19 ± 0.9 d | |
| Seedling Vigor Index (SVI) | Heat-resistant genotypes | ||||||||
| C3 | 1095.9 ± 1.4 d | 1476.2 ± 1.3 c | 1936.5 ± 1.2 a | 1844.4 ± 0.9 b | 167.17 ± 9.8 e | 489.88 ± 10.1 c | 647.3 ± 15.7 cd | 359.02 ± 8.4 h | |
| C7 | 965.3 ± 2.3 d | 1228.2 ± 0.9 c | 1795.5 ± 2.1 a | 1683.7 ± 3.1 b | 65.25 ± 9.8 hi | 78.66 ± 5.5 i | 249.26 ± 8.7 j | 422.64 ± 10.3 g | |
| C8 | 797.7 ± 1.2 d | 1102.3 ± 3.1 c | 1499.8 ± 0.9 b | 2229.2 ± 1.6 a | 60.26 ± 9.9 i | 342.16 ± 15.6 e | 432.91 ± 6.5 f | 673.11 ± 9.8 d | |
| C9 | 849.6 ± 1.3 d | 1133.1 ± 1.8 c | 1331.4 ± 0.9 b | 1562.1 ± 1.2 a | 215.45 ± 11.9 d | 375.10 ± 11.0 e | 1136.54 ± 17.8 a | 1190.01 ± 13.8 b | |
| C10 | 803.4 ± 1.3 d | 1916.0 ± 0.9 c | 1997.4 ± 1.5 b | 2168.3 ± 2.6 a | 263.29 ± 12.1 c | 856.54 ± 16.7 a | 858.50 ± 9.12 b | 1295.57 ± 7.8 a | |
| C14 | 504.7 ± 1.2 d | 1044.8 ± 1.0 c | 1326.2 ± 0.5 b | 1635.0 ± 0.3 a | 335.21 ± 12.4 a | 671.95 ± 12.1 b | 867.20 ± 10.9 b | 1216.39 ± 15.9 a | |
| C15 | 317.3 ± 0.3 d | 1097.1 ± 0.5 c | 1505.7 ± 0.7 b | 1727.8 ± 1.2 a | 116.93 ± 3.6 f | 409.38 ± 13.0 d | 844.30 ± 9.7 b | 1184.10 ± 20.7 b | |
| C16 | 601.0 ± 0.5 d | 883.7 ± 0.7 ac | 1976.1 ± 0.0 a | 1763.0 ± 1.2 b | 303.88 ± 7.9 b | 361.43 ± 10.0 e | 676.57 ± 6.3 c | 1068.72 ± 23.4 c | |
| Heat-sensitive genotypes | |||||||||
| C1 | 333.8 ± 1.7 d | 563.3 ± 3.4 c | 1261.5 ± 5.3 b | 1354.7 ± 0.3 a | 79.4 ± 8.3 h | 113.9 ± 3.3 h | 439.5 ± 14.5 f | 449.1 ± 7.8 fg | |
| C2 | 501.2 ± 0.4 d | 1146.0 ± 0.8 c | 1306.4 ± 1.2 b | 1829.0 ± 2.5 a | 132.42 ± 11.1 f | 258.1 ± 14.5 f | 408.24 ± 14.2 g | 509.28 ± 8.5 f | |
| C4 | 492.8 ± 1.5 d | 687.5 ± 0.9 c | 1245.6 ± 0.9 b | 1319.5 ± 2.5 a | 67.496 ± 13.4 hi | 198.46 ± 5.6 g | 591.72 ± 20.9 d | 371.26 ± 9.9 h | |
| C5 | 976.1 ± 1.3 d | 1041.7 ± 0.9 c | 1524.1 ± 0.8 a | 1358.4 ± 1.3 b | 146.22 ± 10.9 ef | 274.20 ± 7.6 f | 327.88 ± 12.4 h | 533.99 ± 5.6 e | |
| C6 | 1108.6 ± 2.4 d | 1240.3 ± 3.1 c | 1657.5 ± 0.8 a | 1401.9 ± 1.2 b | 16.71 ± 4.6 j | 84.61 ± 6.9 i | 480.35 ± 10.9 e | 249.41 ± 9.7 i | |
| C11 | 385.9 ± 2.4 d | 466.5 ± 2.1 c | 830.3 ± 3.1 b | 1276.2 ± 0.7 a | 107.30 ± 9.9 g | 173.79 ± 14.5 gh | 279.35 ± 11.3 i | 565.46 ± 5.6 e | |
| C12 | 746.0 ± 0.9 d | 830.2 ± 2.1 c | 1614.7 ± 3.1 a | 1284.2 ± 2.6 b | 94.12 ± 12.7 g | 160.85 ± 18.8 gh | 304.41 ± 12.8 h | 71.94 ± 6.2 j | |
| C13 | 543.5 ± 0.6 d | 855.2 ± 2.3 c | 1379.1 ± 0.4 a | 1358.0 ± 0.8 b | 74.90 ± 13.5 h | 200.03 ± 11.8 g | 440.02 ± 13.7 f | 534.29 ± 8.8 e | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cornea-Cipcigan, M.; Cordea, M.I.; Mărgăoan, R.; Pamfil, D. Exogenously Applied GA3 Enhances Morphological Parameters of Tolerant and Sensitive Cyclamen persicum Genotypes under Ambient Temperature and Heat Stress Conditions. Plants 2022, 11, 1868. https://doi.org/10.3390/plants11141868
Cornea-Cipcigan M, Cordea MI, Mărgăoan R, Pamfil D. Exogenously Applied GA3 Enhances Morphological Parameters of Tolerant and Sensitive Cyclamen persicum Genotypes under Ambient Temperature and Heat Stress Conditions. Plants. 2022; 11(14):1868. https://doi.org/10.3390/plants11141868
Chicago/Turabian StyleCornea-Cipcigan, Mihaiela, Mirela Irina Cordea, Rodica Mărgăoan, and Doru Pamfil. 2022. "Exogenously Applied GA3 Enhances Morphological Parameters of Tolerant and Sensitive Cyclamen persicum Genotypes under Ambient Temperature and Heat Stress Conditions" Plants 11, no. 14: 1868. https://doi.org/10.3390/plants11141868