
Photosynthetic Variability of Oblačinska Sour Cherry Ecotypes under Drought

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Visual Symptoms of Drought Stress on Plants
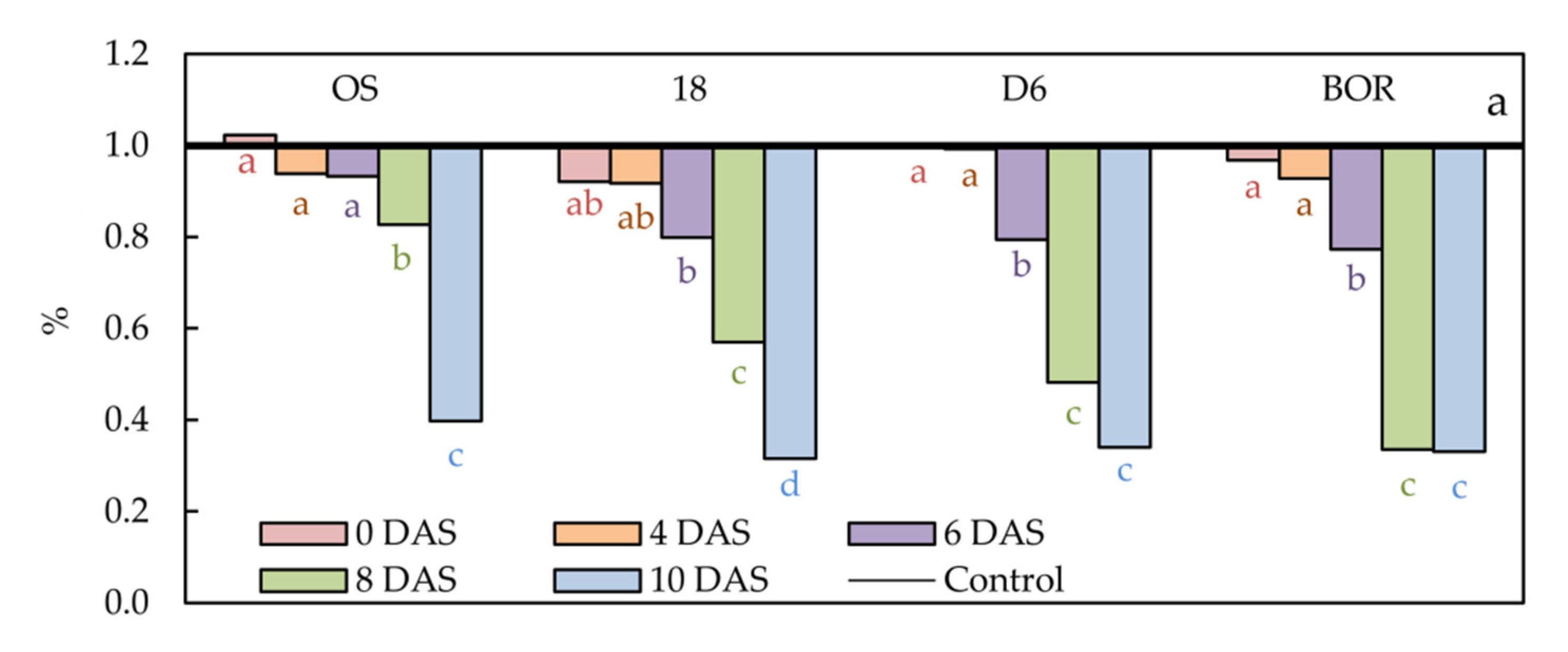
2.2. Relative Water Content
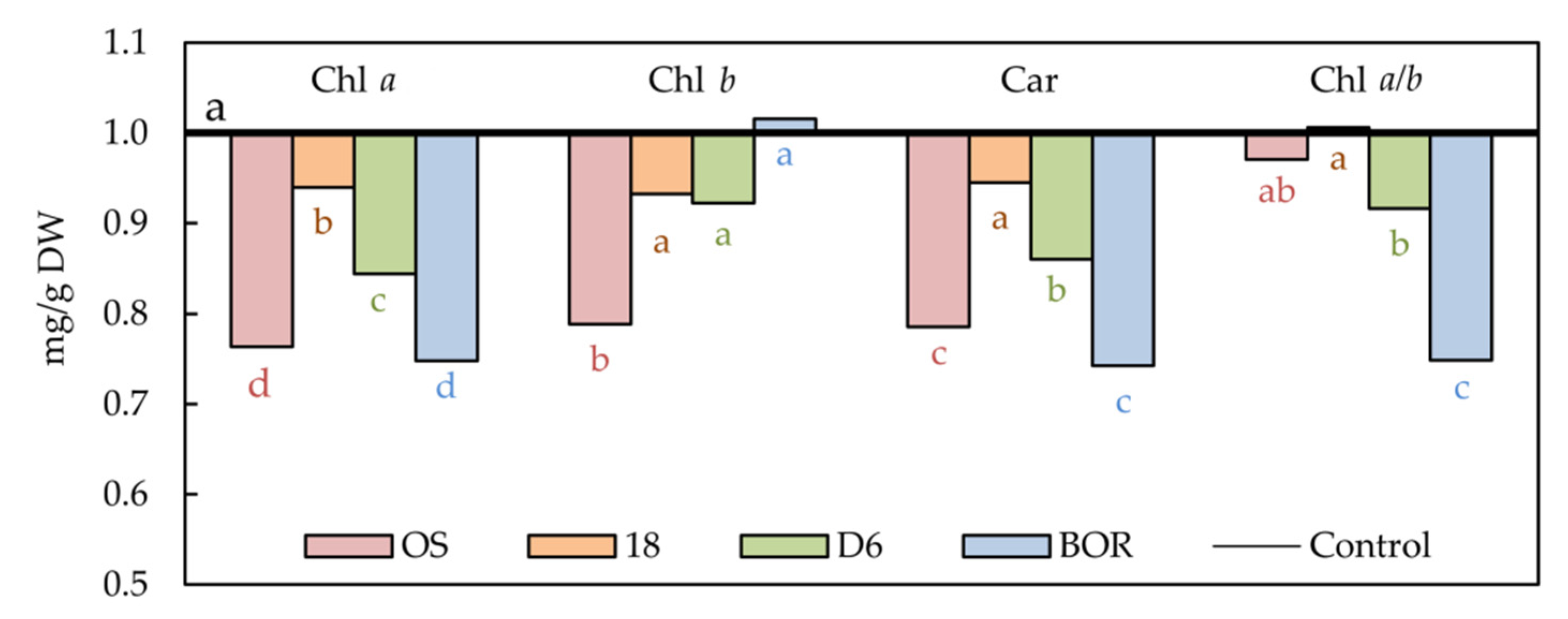
2.3. Photosynthetic Pigments
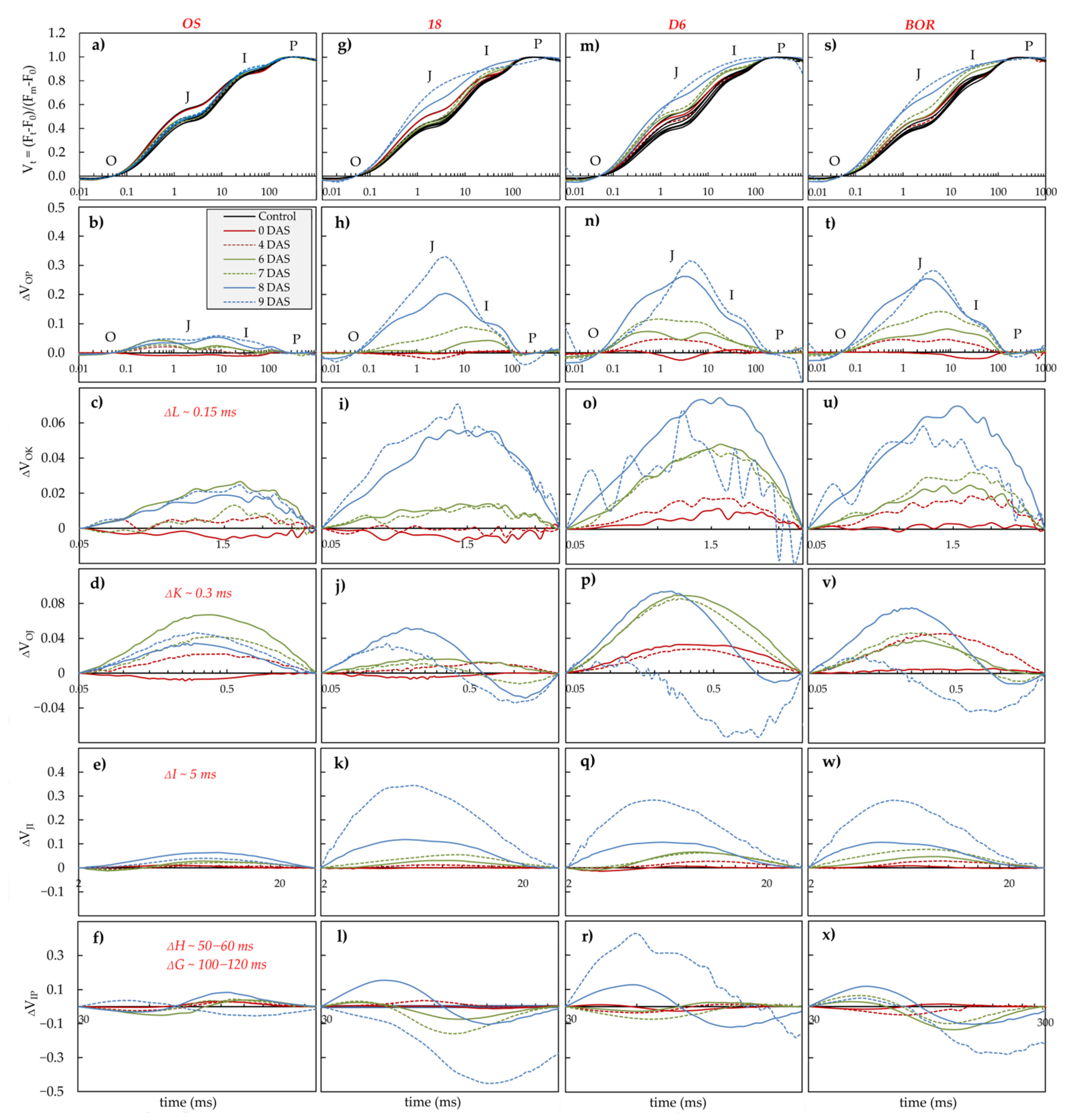
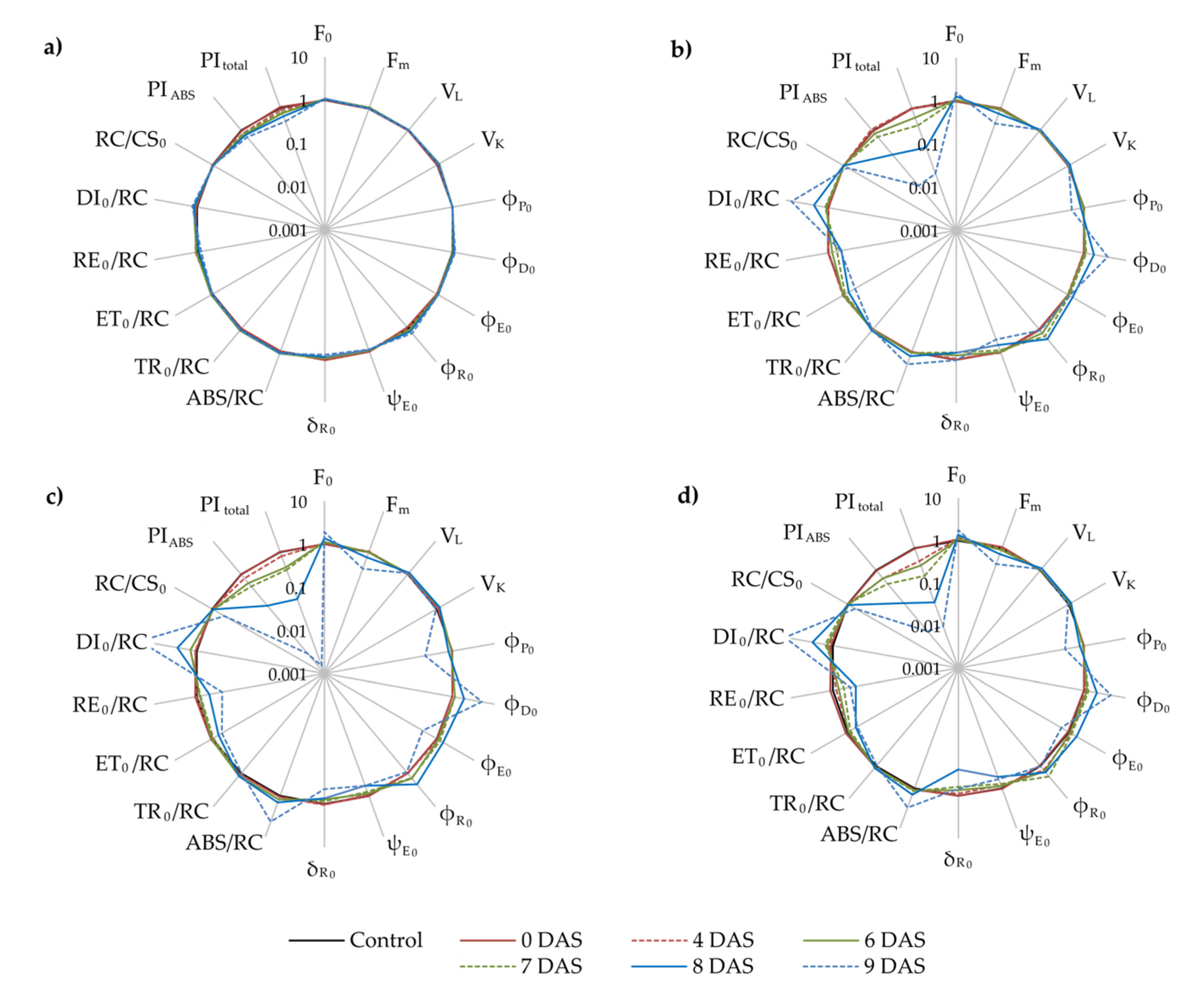
2.4. Photosynthetic Efficiency
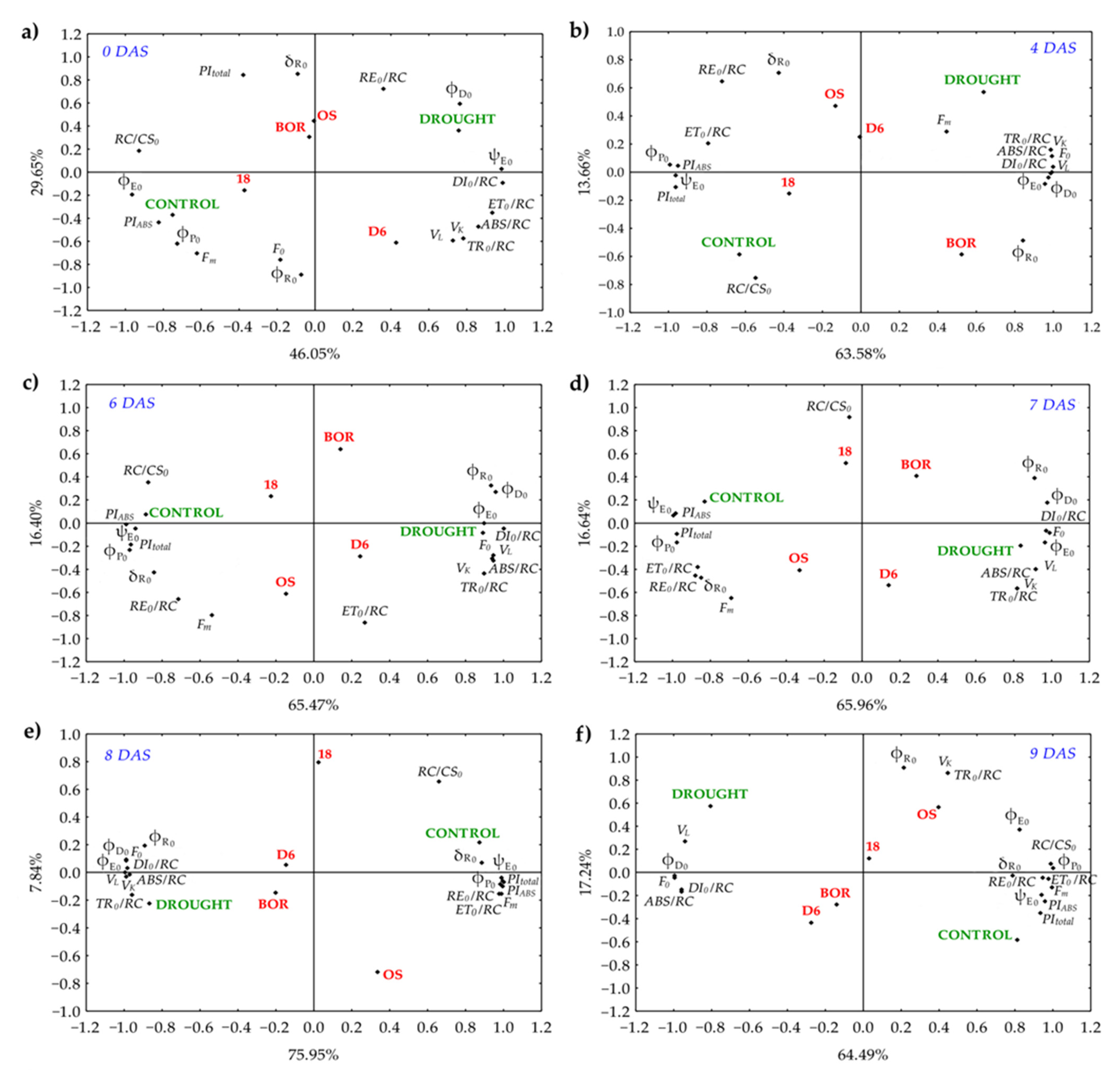
2.5. Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Greenhouse Experiment
4.3. Chlorophyll a Fluorescence Transient
4.4. Relative Water Content
4.5. Determination of Photosynthetic Pigments Concentration
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jaleel, C.A.; Manivannan, P.; Wahidi, A.; Ferooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Pannerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Martínez-García, P.J.; Hartung, J.; Pérez de los Cobos, F.; Martínez-García, P.; Jalili, S.; Sánchez-Roldán, J.M.; Rubio, M.; Dicenta, F.; Martínez-Gómez, P. Temporal Response to Drought Stress in Several Prunus Rootstocks and Wild Species. Agronomy 2020, 10, 1383. [Google Scholar] [CrossRef]
- Mihaljević, I.; Viljevac Vuletić, M.; Šimić, D.; Tomaš, V.; Horvat, D.; Josipović, M.; Zdunić, Z.; Dugalić, K.; Vuković, D. Comparative study of drought stress effects on traditional and modern apple cultivars. Plants 2021, 10, 561. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Tanveer, M.; Naeem, M.; Ali, I.; Tabassum, T.; Nazir, U. Growth and developmental responses of crop plants under drought stress: A review. Zemdirbyste 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Bushal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hort. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Omae, H.; Kumar, A.; Kashiwaba, K.; Shono, M. Assessing drought tolerance of snap bean (Phaseolus vulgaris) from genotypic differences in leaf water relations, shoot growth and photosynthetic parameters. Plant Prod. Sci. 2007, 10, 28–35. [Google Scholar] [CrossRef]
- Faraloni, C.; Cutino, I.; Petruccelli, R.; Leva, A.R.; Lazzeri, S.; Torzillo, G. Chlorophyll fluorescence as a rapid tool for in vitro screening of olive cultivars (Olea europaea L.) tolerant to drought stress. Environ. Exp. Bot. 2011, 73, 49–56. [Google Scholar] [CrossRef]
- Tatari, M.; Jafari, A.; Shirmardi, M.; Mohamadi, M. Using morphological and physiological traits to evaluate drought tolerance of pear populations (Pyrus spp.). Int. J. Fruit Sci. 2020, 20, 837–854. [Google Scholar] [CrossRef]
- Ping, M.A.; Tuan-Hui, B.A.I.; Feng-Wang, M.A. Effects of progressive drought on photosynthesis and portioning of absorbed light in apple trees. J. Integr. Agric. 2015, 14, 681–690. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: London, UK, 2000; pp. 445–483. [Google Scholar]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Christen, D.; Schönmann, S.; Jermini, M.; Strasser, R.J.; Dfago, G. Characterisation and early detection of grapevine (Vitis vinifera) stress responses to esca disease by in situ chlorophyll fluorescence and comparison with drought stress. Environ. Exp. Bot. 2007, 60, 504–514. [Google Scholar] [CrossRef]
- Gomes, M.T.G.; da Luz, A.C.; Rossi dos Santos, M.; Do Carmo Pimentel Batitucci, M.; Moura Silva, D.; Falqueto, A.R. Drought tolerance of passion fruit plants assessed by the OJIP chlorophyll a fluorescence transient. Sci. Hortic. 2012, 142, 49–56. [Google Scholar] [CrossRef]
- Wang, Z.X.; Chen, L.; Ai, J.; Qin, H.Y.; Liu, Y.X.; Xu, P.L.; Jiao, Z.Q.; Zhao, Y.; Zhang, Q.T. Photosynthesis and activity of photosystem II in response to drought stress in Amur Grape (Vitis amurensis Rupr.). Photosynthetica 2012, 50, 189–196. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [Green Version]
- Mihaljević, I.; Viljevac Vuletić, M.; Tomaš, V.; Horvat, D.; Zdunić, Z.; Vuković, D. PSII photochemistry responses to drought stress in autochthonous and modern sweet cherry cultivars. Photosynthetica 2021, 59, 517–528. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Yu, H.Y.; Kong, D.S.; Yan, F.; Zhang, Y.J. Effects of drought stress on growth and chlorophyll fluorescence of Lycium ruthenicum Murr. seedlings. Photosynthetica 2016, 54, 524–531. [Google Scholar] [CrossRef]
- Dias, M.C.; Correia, S.; Serodio, J.; Silva, A.M.S.; Freitas, H.; Santos, C. Chlorophyll fluorescence and oxidative stress endpoints to discriminate olive cultivars tolerance to drought and heat episodes. Sci. Hortic. 2018, 231, 31–35. [Google Scholar] [CrossRef]
- Haider, M.S.; Kurjogi, M.M.; Khalil-ur-Rehman, M.; Pervez, T.; Songtao, J.; Fiaz, M.; Jogaiah, S.; Wang, C.; Fang, J. Drought stress revealed physiological, biochemical and gene-expressional variations in ‘Yoshihime’ peach (Prunus persica L) cultivar. J. Plant Interact. 2018, 13, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Baccari, S.; Elloumi, O.; Chaari-Rkhis, A.; Fenollosa, E.; Morales, M.; Drira, N.; Ben Abdallah, F.; Fki, L.; Munné-Bosch, S. Linking leaf water potential, photosynthesis and chlorophyll loss with mechanisms of photo- and antioxidant protection in juvenile olive trees subjected to severe drought. Front. Plant Sci. 2020, 11, 614144. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Puškar, B. Listing and evaluation of Oblačinska cherry types with the aim of further selection. Pomol. Croat. 2002, 4, 81–93. [Google Scholar]
- Nikolić, D.; Rakonjac, V.; Milutinović, M.; Fotirić, M. Genetic divergence of Oblačinska sour cherry (Prunus cerasus L.) clones. Genetika 2005, 37, 191–198. [Google Scholar] [CrossRef]
- Rakonjac, V.; Fotirić Akšić, M.; Nikolić, D.; Milatović, D.; Čolić, S. Morphological characterization of Oblačinska sour cherry by multivariate analysis. Sci. Hortic. 2010, 125, 679–684. [Google Scholar] [CrossRef]
- Viljevac, M.; Dugalić, K.; Jurković, V.; Mihaljević, I.; Tomaš, V.; Puškar, B.; Lepeduš, H.; Sudar, R.; Jurković, Z. Relation between polyphenols content and skin colour in sour cherry fruits. J. Agric. Sci. 2012, 57, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Vuković, D. Pomological Characteristics of Sour Cherry (Prunus cerasus L. var. Oblačinska) Genotypes in Eastern Slavonia. Master’s Thesis, Josip Juraj Strossmayer University of Osijek, Osijek, Croatia, 2014. [Google Scholar]
- Viljevac, M.; Dugalić, K.; Mihaljević, I.; Šimić, D.; Sudar, R.; Jurković, Z.; Lepeduš, H. Chlorophyll content, photosynthetic efficiency and genetic markers in two sour cherry (Prunus cerasus L.) genotypes under drought stress. Acta Bot. Croat. 2013, 72, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Viljevac Vuletić, M.; Dugalić, K.; Mihaljević, I.; Tomaš, V.; Vuković, D.; Zdunić, Z.; Puškar, B.; Jurković, Z. Season, location and cultivar influence on bioactive compounds of sour cherry fruits. Plant Soil Environ. 2017, 63, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Repajić, M.; Puškar, B.; Dugalić, K.; Vahčić, N.; Srečec, S.; Dragović-Uzelac, D.; Jurković, Z.; Levaj, B. Quality and sensory study of fresh sour cherry juices upon cultivar, growing area and weather conditions. J. Food Sci. 2019, 84, 3264–3274. [Google Scholar] [CrossRef]
- Baraket, G.; Abdallah, D.; Boukhalfa, Y.; Mustapha, S.B. Analysis of genetic diversity and water stress tolerance in Tunisian plums (Prunus spp.; Rosaceae). Sci. Hortic. 2021, 285, 110141. [Google Scholar] [CrossRef]
- Shafqat, W.; Mazrou, Y.S.A.; Sami-ur-Rehman; Nehela, Y.; Ikram, S.; Bibi, S.; Naqvi, S.A.; Hameed, M.; Jaskani, M.J. Effect of three water regimes on the physiological and anatomical structure of stem and leaves of different citrus rootstocks with distinct degrees of tolerance to drought stress. Horticulturae 2021, 7, 554. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Kaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar] [CrossRef] [PubMed]
- Babaei, L.; Sharifani, M.M.; Darvishzadeh, R.; Abbaspour, N.; Henareh, M. Impact of drought stress on photosynthetic response of some pear species. Int. J. Hort. Sci. Technol. 2021, 8, 353–369. [Google Scholar] [CrossRef]
- Gulen, H.; Kesici, M.; Cetinkaya, C.; Ergin, S. Proline and antioxidant enzyme activities in some strawberry cultivars under drought and recovery. Not. Bot. Horti. Agrobot. 2018, 46, 570–578. [Google Scholar] [CrossRef] [Green Version]
- Javadi, T.; Rohollahi, D.; Ghaderi, N.; Nazari, F. Mitigating the adverse effects of drought stress on the morpho-physiological traits and anti-oxidative enzyme activities of Prunus avium through β-amino butyric acid drenching. Sci. Hortic. 2017, 218, 156–163. [Google Scholar] [CrossRef]
- Dbara, S.; Boussetta, W.; Hafi, M.; Mars, M. Performance assessment of three old pear cultivars (Pyrus communis L.) to cope drought caused by climate change. J. Hortic. Postharvest Res. 2021, 4, 351–366. [Google Scholar] [CrossRef]
- Zohouri, M.; Abdollahi, H.; Arji, I.; Abdossi, V. Variations in growth and photosynthetic parameters of some clonal semi-dwarfing and vigorous seedling pear (Pyrus spp.) rootstocks in response to deficit irrigation. Acta Sci. Pol. Hortorum Cultus 2020, 19, 105–121. [Google Scholar] [CrossRef]
- Tanaka, R.; Tanaka, A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes. Biochim. Biophys. Acta 2011, 1807, 968–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahns, P.; Holzwarth, A.R. The role of xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Tomášková, I.; Pastierovič, F.; Krejzková, A.; Čepl, J.; Hradecký, J. Norway spruce ecotypes distinguished by chlorophyll a fluorescence kinetics. Acta Physiol. Plant. 2021, 43, 24. [Google Scholar] [CrossRef]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Govindjee; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta BBA-Bioenerg. 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- van Heerden, P.D.R.; Swanepoel, J.W.; Kruger, G.H.J. Modulation of photosynthesis by drought in two desert scrub species exhibiting C3-mode CO2 assimilation. Environ. Exp. Bot. 2007, 61, 124–136. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.J.; Zhang, S.B.; Liu, T. Cyclic electron flow around photosystem I promotes ATP synthesis possibly helping the rapid repair of photodamaged photosystem II at low light. Front. Plant Sci. 2018, 9, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaji, H.M.; Bąba, W.; Gediga, K.; Goltsev, V.; Samborska, I.A.; Cetner, M.D.; Dimitrova, S.; Piszcz, U.; Bielecki, K.; Karmowska, K.; et al. Chlorophyll fluorescence as a tool for nutrient status identification in rapeseed plants. Photosynth. Res. 2018, 136, 329–343. [Google Scholar] [CrossRef] [Green Version]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindjee. Chlorophyll a fluorescence: A Bit of Basics and History. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 1–42. [Google Scholar]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I.; et al. Drought-induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochim. Biophys. Acta 2012, 1817, 1490–1498. [Google Scholar] [CrossRef] [Green Version]
- Kovačević, J.; Mazur, M.; Drezner, G.; Lalić, A.; Sudarić, A.; Dvojsković, K.; Viljevac Vuletić, M.; Josipović, M.; Josipović, A.; Markulj Kulundžić, A.; et al. Photosynthetic efficiency parameters as indicators of agronomic traits of winter wheat cultivars in different soil water conditions. Genetika 2017, 49, 891–910. [Google Scholar] [CrossRef] [Green Version]
- Mihaljević, I.; Dugalić, K.; Tomaš, V.; Viljevac, M.; Pranjić, A.; Čmelik, Z.; Puškar, B.; Jurković, Z. In vitro sterilization procedures for micropropagation of Oblačinska sour cherry. J. Agric. Sci. 2013, 58, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Živčak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Physiol. Bioch. 2014, 81, 16–25. [Google Scholar] [CrossRef]
- Sitko, K.; Rusinowski, S.; Pogrzeba, M.; Daszkowska-Golec, A.; Kalaji, H.M.; Małkowski, E. Development and aging of photosynthetic apparatus of Vitis vinifera L. during growing season. Photosynthetica 2020, 58, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Driver, J.A.; Kuniyuki, A.H. In vitro propagation of Paradox walnut Juglans hindsii × Juglans regia rootstock. HortSci. 1984, 19, 507–509. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecotype | 6 DAS | 7 DAS | 8 DAS | 9 DAS | 10 DAS |
|---|---|---|---|---|---|
| OS | 0 | 0 | 20 | 25 | 45 |
| 18 | 95 | 100 (10) | 100 (30) | 100 (60) | 100 (75) |
| D6 | 60 | 80 (15) | 90 (55) | 100 (85) | 100 (85) |
| BOR | 70 | 85 (55) | 95 (70) | 95 (75) | 100 (85) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viljevac Vuletić, M.; Horvat, D.; Mihaljević, I.; Dugalić, K.; Šimić, D.; Čupić, T.; Jurković, V.; Lepeduš, H. Photosynthetic Variability of Oblačinska Sour Cherry Ecotypes under Drought. Plants 2022, 11, 1764. https://doi.org/10.3390/plants11131764
Viljevac Vuletić M, Horvat D, Mihaljević I, Dugalić K, Šimić D, Čupić T, Jurković V, Lepeduš H. Photosynthetic Variability of Oblačinska Sour Cherry Ecotypes under Drought. Plants. 2022; 11(13):1764. https://doi.org/10.3390/plants11131764
Chicago/Turabian StyleViljevac Vuletić, Marija, Daniela Horvat, Ines Mihaljević, Krunoslav Dugalić, Domagoj Šimić, Tihomir Čupić, Vlatka Jurković, and Hrvoje Lepeduš. 2022. "Photosynthetic Variability of Oblačinska Sour Cherry Ecotypes under Drought" Plants 11, no. 13: 1764. https://doi.org/10.3390/plants11131764