
Advances in the Micropropagation and Genetic Transformation of Agave Species

Abstract
:1. Introduction
1.1. Agave genus
1.2. Economic Importance of Agave Species

{kind=link}
{kind=link}
| Use | Part of the Plant | Species |
|---|---|---|
| Distilled beverages (mezcal) | Stems and leaves | Agave americana var. americana, A. americana var. oaxacensis, A. angustifolia, A. asperrima, A. convallis, A. duranguensis, A. esperrimia, A. karwinskii, A. marmorata, A. palmeri, A. potatorum, A. rodacantha, A. salmiana, A. seemaniana, A.shrevei, A. tequilana var. azul, A. weberi, A. womomahi, A. zebra |
| Fermented beverages (Aguamiel and pulque) | Stems and leaves | A. americana var. americana, A. angustifolia, A. atrovirens, A. gracillispina, A. hookeri, A. macroculmis, A. malliflua, A. mapisaga, A. salmiana var. salmiana, A. tecta |
| Fibers | Leaves | A. americana var. americana, A. americana var. oaxacensis, A. angustifolia var. angustifolia, A. convallis, A. fourcroydes, A. ghiesbreghtii, A. horrida, A. lechuguilla, A. sisalana |
| Food | Stems, leaves, flower stalk, flowers | A. americana, A. angustiarum, A. angustifolia, A. applanata, A. chiapensis, A. karwinskii, A. marmorata, A. potatorum |
| Medicinal | Leaves, cuticle, juice | A. americana, A. angustiarum, A. marmorata, A. potatorum, A. seemanniana |
| Ornamental | Whole plant | A. americana “Marginata”, A. applanata, A. dasylirioides, A. desmettiana, A. guiengola, A. isthmensis, A. macrocacantha, A. salmiana, A. stricta |
1.3. Genetic Improvement of Agave Species
2. Micropropagation in Agave Species
2.1. Medium, Growth Regulators and Response Comparison of Agave Species Micropropagation
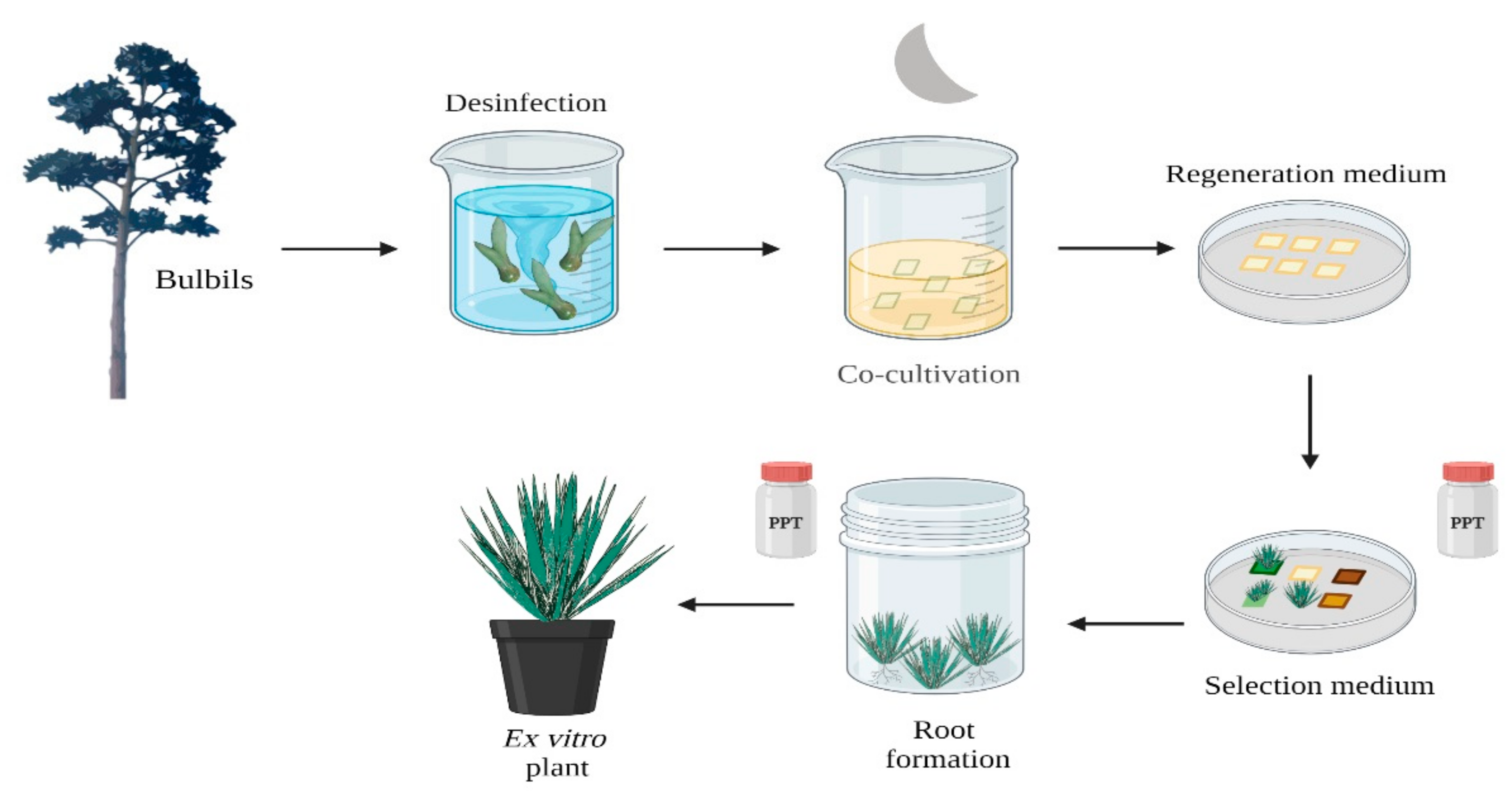
2.2. Genetic Transformation in Agave Species
3. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. 2009, 161, 105–121. [Google Scholar] [CrossRef] [Green Version]
- García-Mendoza, A.J.; Franco Martínez, I.S.; Sandoval Gutiérrez, D. Cuatro especies nuevas de Agave (Asparagaceae, Agavoideae) del sur de Mexico. Acta Bot. Mex. 2019, 126, 1–18. [Google Scholar] [CrossRef]
- García-Mendoza, A.J. Flora del Valle de Tehuacán-Cuicatlán, 1st ed.; Universidad Autónoma de México: Mexico City, Mexico, 2011; pp. 1–95. [Google Scholar]
- Gentry, H.S. Agaves of Continental North America, 1st ed.; University of Arizona Press: Tucson, AZ, USA, 1982; p. 662. [Google Scholar]
- Escobar-Guzman, R.E.; Hernandez, F.Z.; Vega, K.G.; Simpson, J. Seed production and gametophyte formation in Agave tequilana and Agave americana. Botany 2008, 86, 1343–1353. [Google Scholar] [CrossRef]
- Valenzuela-Zapata, A.G. El agave Tequilero, 1st ed.; Monsanto: Guadalajara, Mexico, 1997; p. 204. [Google Scholar]
- Zizumbo-Villarreal, D.; Vargas-Ponce, O.; Rosales-Adame, J.J.; Colunga-GarcíaMarín, P. Sustainability of the traditional management of Agave genetic resources in the elaboration of mezcal and tequila spirits in western Mexico. Genet. Resour. Crop Evol. 2013, 60, 33–47. [Google Scholar] [CrossRef]
- Consejo Regulador del Tequila. Available online: https://www.crt.org.mx/EstadisticasCRTweb/ (accessed on 14 February 2022).
- Servicio de Información Agroalimentaria y Pesquera. Panorama Agroalimentario 2021; SIAP: Mexico City, Mexico, 2021; p. 30. [Google Scholar]
- Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Planeación Agrícola Nacional 2017–2030, 1st ed.; Agave Tequilero y Mezcalero; SAGARPA: Mexico City, Mexico, 2017; p. 2. [Google Scholar]
- García-Mendoza, A.J. Agaváceas. In Biodiversidad de Oaxaca; García-Mendoza, A.J., Ordoñez, M.J., Briones-Salas, M., Eds.; Instituto de Biología, UNAM–Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Fund: Mexico City, Mexico, 2004; pp. 159–169. [Google Scholar]
- López-Romero, J.C.; Ayala-Zavala, J.F.; González-Aguilar, G.A.; Peña-Ramos, E.A.; González-Ríos, H. Biological activities of Agave by products and their possible applications in food and pharmaceuticals. J. Sci. Food Agric. 2018, 98, 2461–2474. [Google Scholar] [CrossRef]
- Ortíz-Basurto, R.I.; Pourcelly, G.; Doco, T.; Williams, P.; Dornier, M.; Belleville, M.P. Analysis of the main components of the aguamiel produced by the maguey-pulquero (Agave mapisaga) throughout the harvest period. J. Agric. Food Chem. 2008, 56, 3682–3687. [Google Scholar] [CrossRef]
- Borland, A.M.; Griffiths, H.; Hartwell, J.; Smith, J.A.C. Exploiting the potential of plants with crassulacean acid metabolism for bioenergy production on marginal lands. J. Exp. Bot. 2009, 60, 2879–2896. [Google Scholar] [CrossRef]
- Davis, S.C.; Dohleman, F.G.; Long, S.P. The global potential for Agave as a biofuel feedstock. GCB Bioenergy 2011, 3, 68–78. [Google Scholar] [CrossRef]
- Parascanu, M.M.; Sanchez, N.; Sandoval-Salas, F.; Carreto, C.M.; Soreanu, G.; Sanchez-Silva, L. Environmental and economic analysis of bioethanol production from sugarcane molasses and agave juice. Environ. Sci. Pollut. Res. 2021, 28, 64374–64393. [Google Scholar] [CrossRef]
- Monja-Mio, K.M.; Herrera-Alamillo, M.A.; Sánchez-Teyer, L.F. Robert, M.L. Breeding Strategies to Improve Production of Agave (Agave spp.). In Advances in Plant Breeding Strategies: Industrial and Food Crops, 1st ed.; Al-Khayri, J.M., Mohan Jain, S., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2019; Volume 6, pp. 319–362. [Google Scholar]
- Lock, G.W. Sisal: Twenty-Five Years’ Sisal Research, 2nd ed.; Longmans, Green & Co., Ltd.: Harlow, UK, 1962; p. 660. [Google Scholar]
- Huang, X.; Xie, L.; Gbokie, T.; Xi, J.; Yi, K. Transcriptome dataset of leaf tissue in Agave H11648. Data 2019, 4, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Vinent Serrano, E. Fajardo Gutiérrez, O. Mejoramiento genético de Agaves en Cuba. Temas de Ciencia y Tecnología. Temas Sci. Tecnol. 2009, 37, 37–43. [Google Scholar]
- Secretaría de Comercio y Fomento Industrial. Norma Oficial Mexicana NOM-006-SCFI-2012. Bebidas alcohólicas- Tequila-Especificaciones. Diario Oficial de la Federación. (12 December 2012). Mexico, D.F. Available online: https://www.dof.gob.mx/nota_detalle.php?codigo=5282165&fecha=13/12/2012#gsc.tab=0 (accessed on 15 May 2022).
- Arroyo, J.; Real de Pénjamo, Penjamo, Guanajuato, Mexico. Personal communication, 2015.
- Monja-Mio, K.M.; Olvera-Casanova, D.; Herrera-Herrera, G.; Herrera-Alamillo, M.Á.; Sánchez-Teyer, F.L.; Robert, M.L. Improving of rooting and ex vitro acclimatization phase of Agave tequilana by temporary immersion system (BioMINTTM). Vitr. Cell. Dev. Biol.-Plant 2020, 56, 662–669. [Google Scholar] [CrossRef]
- Powers, D.E.; Backhaus, R.A. In vitro propagation of Agave arizonica Gentry and Weber. Plant Cell Tissue Organ Cult. 1989, 16, 57–60. [Google Scholar] [CrossRef]
- Rodríguez-Garay, B.; Gutiérrez-Mora, A.; Acosta-Duefias, B. Somatic embryogenesis of Agave victoria-reginae Moore. Plant Cell Tissue Organ Cult. 1996, 46, 85–87. [Google Scholar] [CrossRef]
- Robert, M.; Herrera, J.L.; Contreras, F.; Scorer, K.N. In vitro propagation of Agave fourcroydes Lem. (Henequen). Plant Cell Tissue Organ Cult. 1987, 8, 37–48. [Google Scholar] [CrossRef]
- Área Agrícola de Casa Sauza. Available online: https://www.casasauza.com/procesos-tequila-sauza/micropropagacion-agave-azul (accessed on 31 October 2017).
- Flores-Benítez, S.; Jiménez-Bremont, J.F.; Rosales-Mendoza, S.; Argüello-Astorga, G.R.; Castillo-Collazo, R.; Alpuche-Solís, A.G. Genetic transformation of Agave salmiana by Agrobacterium tumefaciens and particle bombardment. Plant Cell Tissue Organ Cult. 2007, 91, 215–224. [Google Scholar] [CrossRef]
- Ramírez-Malagón, R.; Borodanenko, A.; Pérez-Moreno, L.; Salas-Araiza, M.D.; Nuñez-Palenius, H.G.; Ochoa-Alejo, N. In vitro propagation of three Agave species used for liquor distillation and three for landscape. Plant Cell Tissue Organ Cult. 2008, 94, 201–207. [Google Scholar] [CrossRef]
- Binh, L.T.; Muoi, L.T.; Oanh, H.T.K.; Thang, T.D.; Phong, D.T. Rapid propagation of agave by in vitro tissue culture. Plant Cell Tissue Organ Cult. 1990, 23, 67–70. [Google Scholar] [CrossRef]
- Monja-Mio, K.M.; Robert, M.L. Direct somatic embryogenesis of Agave fourcroydes Lem. through thin cell layer culture. Vitr. Cell. Dev. Biol.-Plant 2013, 49, 541–549. [Google Scholar] [CrossRef]
- Valenzuela-Sánchez, K.K.; Juárez-Hernández, R.E.; Cruz-Hernández, A.; Olalde-Portugal, V.; Valverde, M.E.; Paredes-Lopez, O. Plant regeneration of Agave tequilana by indirect organogenesis. Vitr. Cell. Dev. Biol.-Plant 2008, 42, 336–340. [Google Scholar] [CrossRef]
- Tejavathi, D.H.; Rajanna, M.D.; Sowmya, R.; Gayathramma, K. Induction of somatic embryos from cultures of Agave vera-cruz Mill. Vitr. Cell. Dev. Biol.-Plant 2007, 43, 423–428. [Google Scholar] [CrossRef]
- Monja-Mio, K.M.; Olvera-Casanova, D.; Herrera-Alamillo, M.Á.; Sánchez-Teyer, F.L.; Robert, M.L. Comparison of conventional and temporary immersion systems on micropropagation (multiplication phase) of Agave angustifolia Haw.’Bacanora’. 3 Biotech 2021, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Santacruz-Ruvalcaba, F.; Portillo, L. Thin cell suspension layer as a new methodology for somatic embryogenesis in Agave tequilana Weber cultivar azul. Ind. Crops Prod. 2009, 29, 609–614. [Google Scholar] [CrossRef]
- Santacruz-Ruvalcaba, F.; Gutiérrez-Pulido, H.; Rodríguez-Garay, B. Efficient in vitro propagation of Agave parrasana Berger. Plant Cell Tissue Organ Cult. 1999, 56, 163–167. [Google Scholar] [CrossRef]
- Hazra, S.K.; Das, S.; Das, A.K. Sisal plant regeneration via organogenesis. Plant Cell Tissue Organ Cult. 2002, 70, 235–240. [Google Scholar] [CrossRef]
- Sánchez-Urbina, A.; Ventura-Canseco, L.M.C.; Ayora-Talavera, T.; Abud-Archila, M.; Peez-Farrera, M.A.; Dendooven, L.; Miceli, F.G. Seed germination and in vitro propagation of Agave grijalvensis an endemic endangered Mexican species. Asian J. Plant Sci. 2008, 7, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Li, X.; Chen, Z.; Li, J.F.; Lu, J.Y.; Zhou, W.Z. Shoot organogenesis and plant regeneration in Agave hybrid, No. 11648. Sci. Hortic. 2013, 161, 30–34. [Google Scholar] [CrossRef]
- Puente-Garza, C.A.; Gutiérrez-Mora, A.; García-Lara, S. Micropropagation of Agave salmiana: Means to production of antioxidant and bioactive principles. Front. Plant Sci. 2015, 6, 1026. [Google Scholar] [CrossRef] [Green Version]
- Lecona-Guzmán, C.A.; Reyes-Zambrano, S.; Barredo-Pool, F.A.; Abud-Archila, M.; Montes-Molina, J.A.; Rincón-Rosales, R.; Gutierrez-Miceli, F.A. In vitro propagation of Agave americana by indirect organogenesis. HortScience 2017, 52, 996–999. [Google Scholar] [CrossRef]
- Naziri, M.; Sadat, S.; Soltani Howyzeh, M. The effect of different hormone combinations on direct and indirect somatic embryogenesis in Agave americana. Iran. J. Plant Physiol. 2019, 9, 2739–2747. [Google Scholar]
- Delgado-Aceves, L.; Portillo, L.; Folgado, R.; Romo-Paz, F.J.; Gonzalez Arnao, M.T. New Approaches for Micropropagation and Cryopreservation of Agave peacockii, An Endangered Species. Plant Cell Tissue Organ Cult. 2022, 150, 85–89. [Google Scholar] [CrossRef]
- Chávez-Ortiz, L.I.; Morales-Domínguez, J.F.; Rodríguez-Sahagún, A.; Pérez-Molphe-balch, E. In vitro propagation of Agave guiengola Gentry using semisolid medium and temporary immersion bioreactors. Phyton 2021, 90, 1003–1013. [Google Scholar] [CrossRef]
- Cabrera-Ponce, J.L. Transformación Genética en El Género Agave Y Producción de Plantas Transgénicas Resistentes a Herbicidas. JL02000044. 2002. Available online: https://svt.cinvestav.mx/Portals/svt/descarga/Fichas/Agave%20tequilero-MXJL02000044A.pdf (accessed on 16 May 2022).
- Rodríguez, G.; Morales, F.; Gutiérrez, R.; Aguilar, S.; Pérez, E. Generación de raíces transformadas de Agave salmiana Otto y su colonización por Glomus intraradices. Rev. Fitotec. 2007, 30, 215–222. [Google Scholar] [CrossRef]
- Gao, J.; Yang, F.; Zhang, S.; Li, J.; Chen, H.; Liu, Q.; Zheng, J.; Xi, J.; Yi, K. Expression of a hevein-like gene in transgenic Agave hybrid No. 11648 enhances tolerance against zebra stripe disease. Plant Cell Tissue Organ Cult. 2014, 119, 579–585. [Google Scholar] [CrossRef]
- Gutiérrez-Aguilar, P.R.; Gil-Vega, K.C.; Simpson, J. Development of an Agrobacterium tumefasciens mediated transformation protocol for two Agave species by organogenesis. Sustainable and Integral Exploitation of Agave. Gutiérrez-Mora, A., Rodríguez-Garay, B., Contreras-Ramos, S.M., Kirchmayr, M.R., González-Ávila, M., Eds.; 2014. Available online: http://www.ciatej.net.mx/agave/1.7agave.pdf (accessed on 15 May 2022).
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]


| Species | Source of Explant | Culture Media and Growth Regulators | Response | Rate | References |
|---|---|---|---|---|---|
| A. fourcroydes | Rhizome and stem tissue | Callus induction: Gamborg 1.12 µM 2,4-D + 4.44 µM BA Shoot regeneration: MMS 1.12 µM 2,4-D + 4.44 µM BA | IO | Abundant rootlike structures. 4 shoots per callus after 12 to 16 weeks of subculture | [26] |
| A. arizonica | Bulbils | Callus induction: MMS 1.4µM 2,4-D Shoot regeneration: MMS 44.4µM BA + 0.5 or 5.4 µM NAA | IO | 5–10 shoots per callus after 12 weeks | [24] |
| A. sisalana, A. fourcroydes, A. cantala | Stolons | MS 0.40 µM NAA + 0.49 µM IBA + 2.32 µM KIN | DO | Multiplication index ranged between 3 and 4 for 4–5 weeks of subculture | [30] |
| A. victoria-reginae Moore | Young leaf blades from in vitro plantlets | Induction of somatic embryogenesis: MS medium + L2 vitamins + 1.4 µM 2,4-D Germination: half-strength MS without growth regulators | DSE | Not specified, but there were somatic embryos at the globular stage on 100% of the explants after 2 weeks | [25] |
| A. parrasana | Offshoots multiplied in vitro | MS + L2 vitamins + 13.3 µM BA | DO | 19.9 shoots per explant | [36] |
| A. sisalana | In vitro grown immature leaves and rhizome | Callus initiation: MS + 9.05 µM 2,4-D + 4.6 µM KIN (In light conditions) Shoot regeneration: MS + 26.6 µM BA | IO | 25.3 shoots per callus after 21 days of co-culture. | [37] |
| A. vera-cruz Mill | Shoot apices; cotyledons and leaf segments; and seeds | Induction of somatic embryogenesis: MS + L2 vitamins + 4.52 μM 2,4-D or 5.37 μM NAA, 4.8 μM IBA and 5.70 μM IAA Maturation: 5.37 μM NAA + 0.91 μM zeatin | ISE | Shoot tip: 16 embryos per explant Cotyledon: 17 embryos per explant In vitro leaf: 17 embryos per explant | [33] |
| A. grijalvensis | Germinated seeds in vitro | Shoot induction: MS + 38.2 µM BA | DO | Not specified | [38] |
| A. tequilana | Offsets | MMS + 6.8 mM 2,4-D for 3 days, | DO | 12 axillary shoots per explant after 60 days | [29] |
| A. salmiana ssp. crassispina, | Offsets | 0.49 µM IBA + 4.44 µM BA | DO | 3 axillary shoots per explant after 60 days | [29] |
| A. duranguensis | Offsets | 0.049 µM IBA + 4.44 µM BA | DO | 6 axillary shoots per explant after 60 days | [29] |
| A. oscura | Offsets | 2.46 µM IBA + 4.44 µM BA | DO | 13 axillary shoots per explant after 60 days | [29] |
| A. pigmaea | Offsets | 0.49 µM IBA + 13.31 µM BA | DO | 6 axillary shoots per explant after 60 days | [29] |
| A. victoria-reginae | Offsets | 2.46 µM IBA + 2.22 µM BA | DO | 6 axillary shoots per explant after 60 days | [29] |
| A. tequilana var. azul | Callus from in vitro plants | 9 µM 2,4-D + 1.3 µM BA | ISE | 556.8 embryos using a celular suspension 161 × 103 cell mL−1 | [35] |
| Hybrid N11648 | Shoot tip and immature leaf tissues | Callus induction: MS + 8.88 µM BA + 1.07 µM NAA Shoot regeneration: SH medium + 22.2 µM BA + 0.1 mg/l NAA + 0.492 µM IBA | IO | Shoot tip: 13.9 shoots | [39,40] |
| A. fourcroydes | In vitro plantlets | Induction of somatic embryogenesis: MS + L2 vitamins + 2.26 µM dicamba or 2.07 µM picloram Germination: half-strength MS without growth regulators | DSE | 92.22 embryos/explant, and 81.72 embryos/explant respectively | [31] |
| A. salmiana | Plantlets from in vitro germinated seeds | MS + L2 vitamins + 0.18 µM 2,4-D + 44.4 µM BA | DO | 14 axillary shoots | [40] |
| A. americana | Meristematic tissue | Callus induction: MS + 0.11 µM 2,4-D + 58.7 or 73.3 µM BA Shoot regeneration: MS medium without growth regulators | IO | 71 shoots per callus after 36 weeks | [41] |
| A. americana | Shoots extracted from rhizomes | Embryo induction: MS + L2 vitamins + 10.3 µM picloram Callus induction 9.04 µM 2,4-D Germination: MSB medium without growth regulators | ISE | Not specified | [42] |
| A. peacockii | Rhizomatous shoots | MS + 26.6µM BA + 27.84 µM KIN | DO | 87 shoots after 60 days of co-culture | [43] |
| A. angustifolia | In vitro plants | MMS 0.1 µM 2,4-D + 44.4 µM BA SS: MS + 0.1 µM 2,4-D + 44.4 µM BA TIS: RITA bioreactor | DO | 6.23 shoots per plant | [34] |
| A. guiengola | Axillary sprouting from stem segments | MS + 8.88 µM BA | DO | 3.7 shoots per explant | [44] |
| Species | Method | Selectable Marker and Reporter Gene | Culture Conditions | Rate | Reference |
|---|---|---|---|---|---|
| Agave genus | Co-cultivation with A. tumefaciens | bar; uidA | Information not specified | Resistant plants to herbicide or antibiotic | [45] |
| Particle borbadment | hpt; uidA | Resistant plants to antibiotic | |||
| A. salmiana | Co-cultivation with A. tumefaciens | nptII; uidA | Bacteria exposure: 30 min Co-cultivation: MS50 + 5 µM BA + 2.7 µM NAA + 100 µM acetosyringone Callus induction: MS50 + 5 µM BA + 2.7 µM NAA + 10 mL L−1 cocktail 20 + 50 mg L−1 Kn + 250 mg L−1 Cf Rooted media: MS50 + 1.14 µM IAA + 12.5 µM BA + 50 mg L−1 Kn | 32 rooted transgenic plants; transgenic calli | [28] |
| Particle bombardment | bar, uidA | Callus induction: MS + 5 µM BA + 2.7 µM NAA Somatic embryos induction: MS + 5 µM BA + 2.7 µM NAA + 10 mL L−1 cocktail 20 Bombard pressure 1100 psi Selection media: MS + 5 µM BA + 2.7 µM NAA + 10 mL L−1 cocktail 20 + 0.5 mg L−1 BASTA | Green calli | ||
| A. salmiana | Co-cultivation with A. rhizogenes | nptII, uidA | Mechanical injury with a needle + 1 × 109 bacteria mL−1 + 200 µM acetosyringone Co-cultivation 6 days with no light Response induction: MS + 500 mg L−1 Cf | Transformed roots | [46] |
| Hybrid 11648 | Co-cultivation with A. tumefaciens | bar, uidA | Callus induction: SH + 13.2 µM BA + 2.68 µM NAA + 0.45 µM 2,4-D + 6.5 g L−1 carrageenan Induction medium: SH + 6.66 µM BA + 2.68 µM NAA + 6.5 g L−1 carrageenan + 2 mg L−1 PPT Rooting induction: SH IAA | 37 Transformed lines | [47] |
| A. tequilana and A. desmettiana | Co-cultivation with A. tumefaciens | bar, uidA | Strain: GV2260 and LBA4404 Shoot induction: BA + IBA, concentrations not especified | Shoots, transformation not specified | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bautista-Montes, E.; Hernández-Soriano, L.; Simpson, J. Advances in the Micropropagation and Genetic Transformation of Agave Species. Plants 2022, 11, 1757. https://doi.org/10.3390/plants11131757
Bautista-Montes E, Hernández-Soriano L, Simpson J. Advances in the Micropropagation and Genetic Transformation of Agave Species. Plants. 2022; 11(13):1757. https://doi.org/10.3390/plants11131757
Chicago/Turabian StyleBautista-Montes, Erika, Laura Hernández-Soriano, and June Simpson. 2022. "Advances in the Micropropagation and Genetic Transformation of Agave Species" Plants 11, no. 13: 1757. https://doi.org/10.3390/plants11131757