
Evaluation of Ethanolic Powdered Extract of Magnolia tamaulipana Vazquez against Oligonychus punicae Hirst (Trombidiformes: Tetranychidae)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Phytochemical Composition
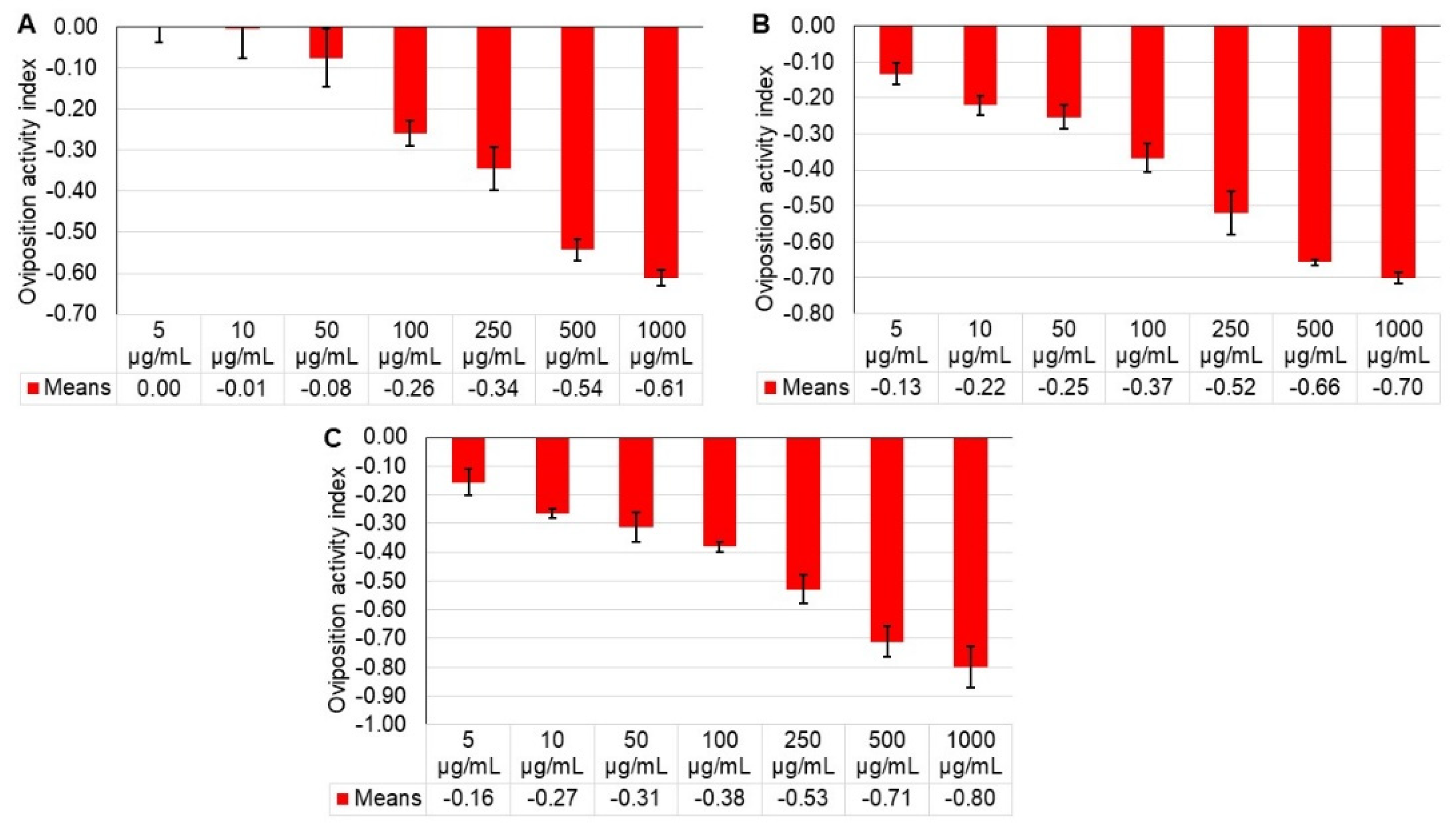
2.2. Oviposition
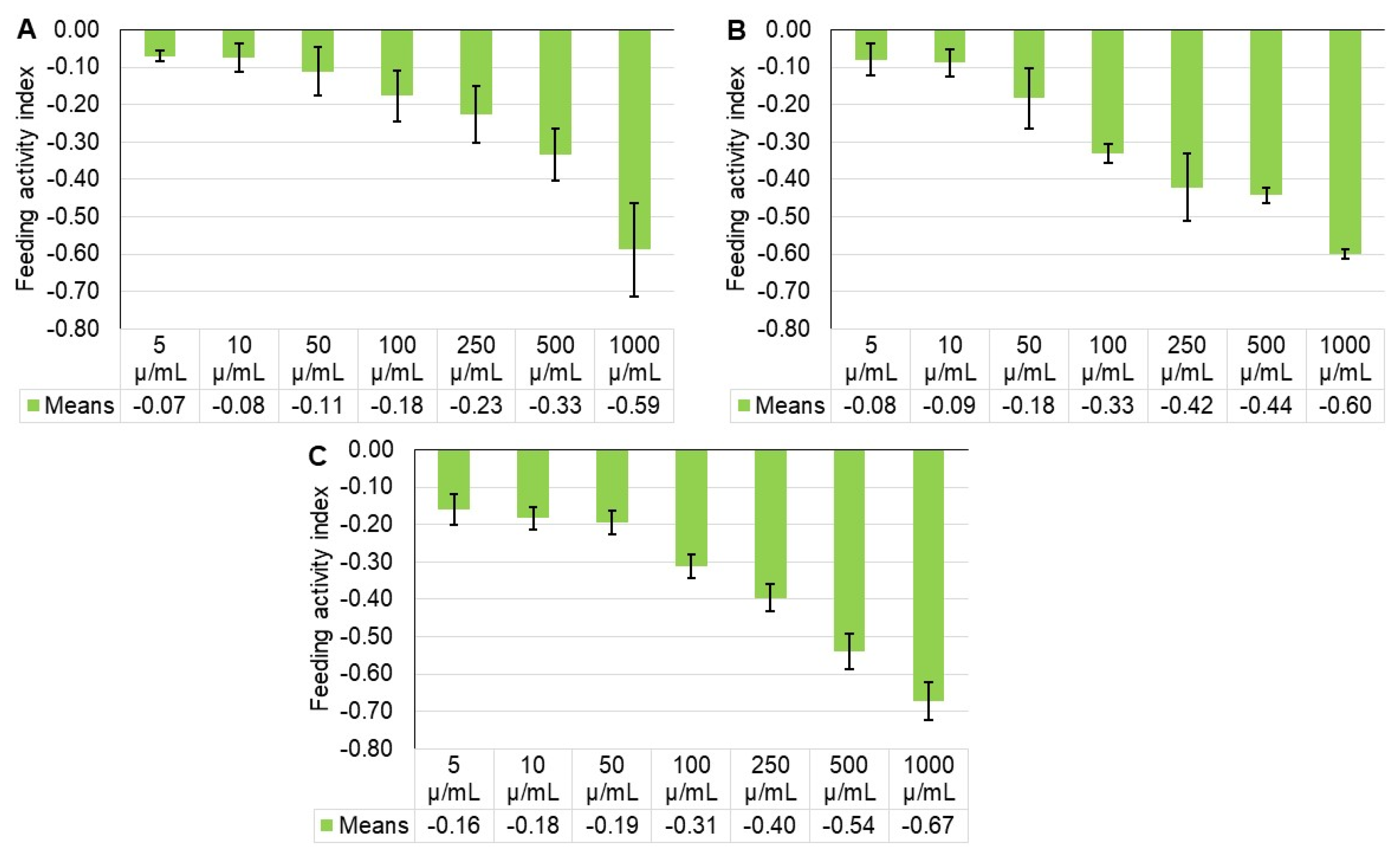
2.3. Feeding Damage
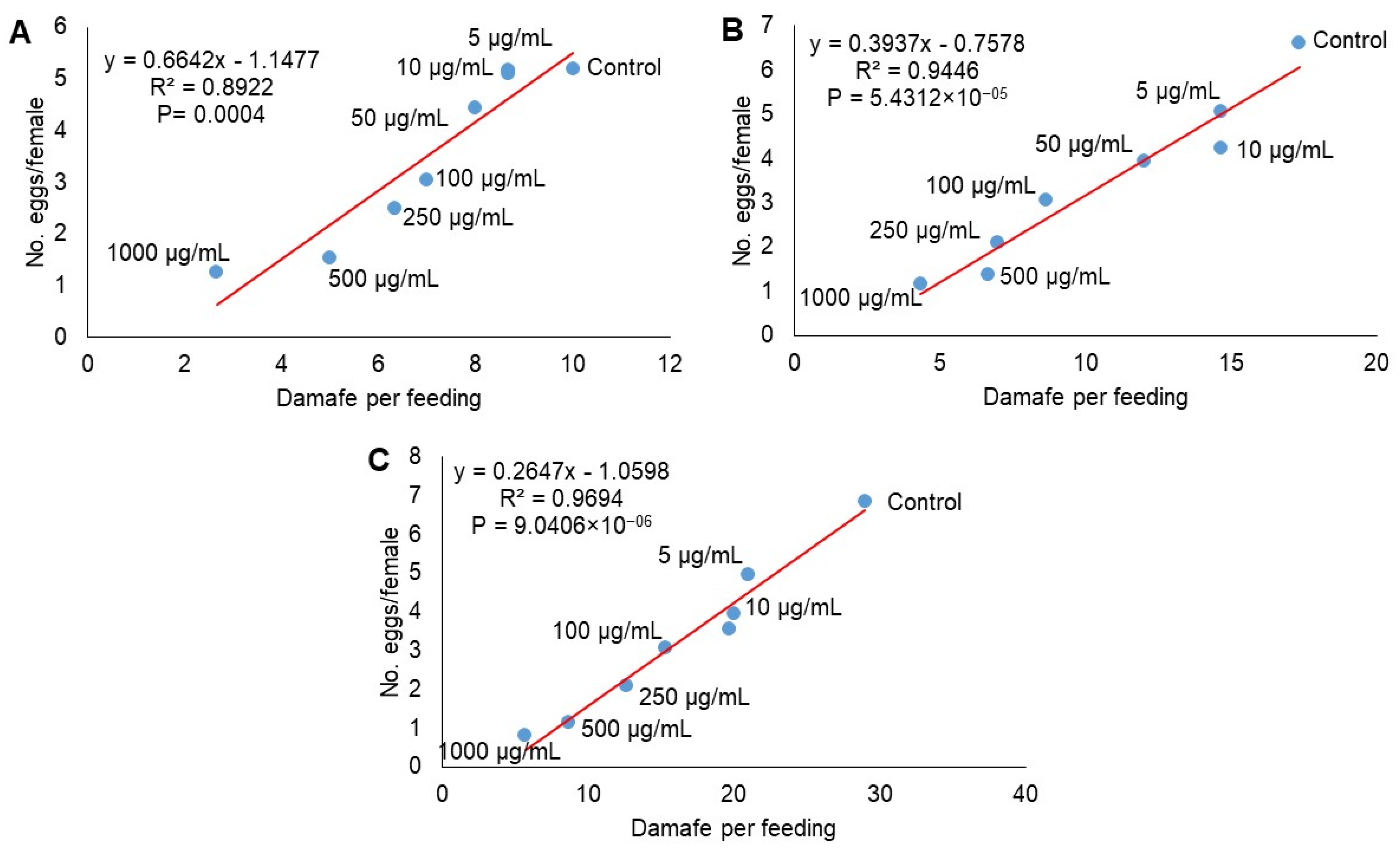
2.4. Demographic Parameters
3. Discussion
4. Materials and Methods
4.1. Colonia de Red Spider Mite Colony
4.2. Preparation of the Plant Material and the Extract
4.3. Phytochemical Extract Analysis
4.4. Experimental Design
4.5. Oligonychus punicae Oviposition
4.6. Oligonychus punicae Feeding
4.7. Oligonychus punicae Growth Population
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peña, J.; Wysoki, M. Plagas en Israel, México, Estados Unidos, Centroamérica y Perú: Plagas del palto en México. In Manejo Integrado de Plagas en Paltos y Cítricos; Ripa, R., Larral, P., Eds.; Colección Libros INIA. No. 23; Instituto de Investigaciones Agropecuarias INIA: Valparaíso, Chile, 2008; pp. 303–309. [Google Scholar]
- Cerna, E.; Badii, M.H.; Ochoa, Y.; Aguirre, L.A.; Landeros, J. Life table of Oligonychus punicae Hirst (Acari: Tetranychidae) inavocado leaves (Persea americana Mill) in the hass, fuerte and criollo cultivars. Univ. Cienc. 2009, 25, 133–140. [Google Scholar]
- Aponte, O.; McMurtry, J.A. Damage on Hass avocado leaves, webbing and nesting behavior of Oligonychus perseae (Acari: Tetranychidae). Exp. Appl. Acarol. 1997, 21, 265–272. [Google Scholar] [CrossRef]
- Humeres, E.C.; Morse, J.G. Baseline susceptibility of persea mite (Acari: Tetranychidae) to abamectin and milbemectin in avocado groves in Southern California. Exp. Appl. Acarol. 2005, 36, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, C.; Aponte, O.; Morales, J.; Sanabria, M.E.; García, G. Biological studies of Oligonychus punicae (Acari: Tetranychidae) on grapevine cultivars. Exp. Appl. Acarol. 2008, 45, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Saren, J.; Mukhopadhyay, A. Acaricide susceptibility of Oligonychus coffeae Nietner (Acari: Tetranychidae) with corresponding changes in detoxifying enzyme levels from tea plantations of sub-Himalayan Terai, India. Acarologia 2017, 57, 581–590. [Google Scholar] [CrossRef]
- Ferraz, J.C.B.; Silva, P.R.R.; Amaranes, M.P.; Silva-Melo, J.W.; Lima, D.B.; França, S.M. Biology and fertility life table of Oligonychus punicae Hirst (Acari: Tetranychidae) associated with eucaliptus in a clonal minigarden. Syst. Appl. Acarol. 2020, 25, 103–112. [Google Scholar] [CrossRef]
- Attia, S.; Grissa, K.L.; Lognay, G.; Bitume, E.; Hance, T.; Mailleux, A.C. A review of the major biological approaches to control the worldwide pest Tetranychus urticae (Acari: Tetranychidae) with special reference to natural pesticides. J. Pest Sci. 2013, 86, 361–386. [Google Scholar] [CrossRef]
- Giraldo-Rivera, A.I.; Guerrero-Álvarez, G.D. Botanical biopesticides: Research and development trends, a focus on the Annonaceae family. Rev. Colomb. Cienc. Hort. 2019, 13, 371–383. [Google Scholar] [CrossRef]
- Lengai, G.M.W.; Muthomi, J.W.; Mbga, E.R. Phytochemical activity and role of botanical pesticides in pest management for sustainable agricultural crop production. Sci. Afr. 2020, 7, e00239. [Google Scholar] [CrossRef]
- Miyazawa, M.; Ishikawa, Y.; Kasahara, H.; Yamanaka, J.I.; Kameoka, H. An insect growth inhibitory ljgnan from flower buds of Magnolia fargesii. Phytochemistry 1994, 35, 611–613. [Google Scholar] [CrossRef]
- Park, I.K.; Park, J.D.; Kim, C.S.; Shin, S.C.; Ahn, Y.J.; Park, S.C.; Lee, S.G. Insecticidal and acaricidal activities of domestic plant extracts against five major arthropod pests. Korean J. Pestic. Sci. 2002, 6, 271–278. [Google Scholar]
- Flores-Estévez, N.; Vasquez-Morales, S.G.; Cano-Medina, T.; SánchezVelásquez, L.R.; Noa-Carrazana, J.C.; Diáz-Fleischer, F. Insecticidal activity of raw ethanolic extracts from Magnolia dealbata Zucc on a tephritid pest. J. Environ. Sci. Health Part B 2013, 48, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Vásquez-Morales, S.G.; Flores-Estévez, N.; Sánchez-Velásquez, L.F.; Pineda-López, M.R.; Viveros-Viveros, H.; Díaz-Fleischer, F. Bioprospecting of botanical insecticides: The case of ethanol extracts of Magnolia schiedeana Schltl. applied to a tephritid fruit fly Anastrepha ludens Loew. J. Entomol. Zool. Stud. 2015, 3, 1–5. [Google Scholar]
- Chacón-Hernández, J.C.; Arredondo-Valdés, R.; Anguiano-Cabello, J.C.; Ordaz-Silva, S.; Hernández-Juárez, A.; Reyes-Zepeda, F. Effect of Magnolia tamaulipana extract on egg laying and food intake of Tetranychus urticae (Acari: Tetranychidae). Int. J. Acarol. 2019, 46, 108–110. [Google Scholar] [CrossRef]
- Luu-Dam, N.A.; Tabanca, N.; Estep, A.S.; Nguyen, D.H.; Kendra, P.E. Insecticidal and attractant activities of Magnolia citrata leaf essential oil against two major pests from Diptera: Aedes aegypti (Culicidae) and Ceratitis capitata (Tephritidae). Molecules 2021, 26, 2311. [Google Scholar] [CrossRef]
- Dieringer, G.; Cabrera, L.; Lara, M.; Loya, L.; Reyes-Castillo, P. Beetle pollination and floral thermogenicity in Magnolia tamaulipana (Magnoliaceae). Int. J. Plant Sci. 1999, 160, 64–71. [Google Scholar] [CrossRef]
- Vázquez-G., J.A. Magnolia (Magnoliaceae) in Mexico and Central America: A synopsis. Britonia 1994, 46, 1–23. [Google Scholar] [CrossRef]
- Arredondo-Valdés, R.; Chacón-Hernández, J.C.; Reyes-Zepeda, F.; Hernández-Castillo, F.D.; Anguiano-Cabello, J.C.; Heinz-Castro, R.T.Q.; Mora-Ravelo, S.G. In vitro antibacterial activity of Magnolia tamaulipana against tomato phytopathogenic bacteria. Plant Prot. Sci. 2020, 56, 268–274. [Google Scholar] [CrossRef]
- Hikal, W.M.; Baeshen, R.S.; Said-Al Ahl, H.A.H. Botanical insecticide as simple extractives for pest control. Cogent Biol. 2017, 3, 1404274. [Google Scholar] [CrossRef]
- Tlak Gajger, I.; Dar, S.A. Plant allelochemicals as sources of insecticides. Insects 2021, 12, 189. [Google Scholar] [CrossRef]
- Kim, J.; Tooker, J.F.; Luthe, D.S.; De Moraes, C.M.; Felton, G.W. Insect eggs can enhance wound response in plants: A study system of tomato Solanum lycopersicum L. and Helicoverpa zea Boddie. PLoS ONE 2012, 7, e37420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivira, A.; Sanabria, M.E.; Valera, N.; Vásquez, C. Toxicity of ethanolic extracts from Lippia origanoides and Gliricidia sepium to Tetranychus cinnabarinus (Boisduval) (Acari: Tetranychidae). Neotrop. Entomol. 2010, 40, 375–379. [Google Scholar]
- Pavela, R.; Murugan, K.; Canale, A.; Benelli, G. Saponaria officinalis-synthesized silver nanocrystals as effective biopesticides and oviposition inhibitors against Tetranychus urticae Koch. Ind. Crops Prod. 2017, 97, 338–344. [Google Scholar] [CrossRef]
- Pavela, R.; Dall’Acqua, S.; Sut, S.; Baldan, V.; Ngahang, S.L.; Biapa, P.C.; Cappellacci, L.; Petrelli, R.; Nicoletti, M.; Canale, A.; et al. Oviposition inhibitory activity of the Mexican sunflower Tithonia diversifolia (Asteraceae) polar extracts against the two-spotted spider mite Tetranychus urticae (Tetranychidae). Physiol. Mol. Plant Pathol. 2018, 101, 85–92. [Google Scholar] [CrossRef]
- Ribeiro, L.P.; Zanardi, O.Z.; Vendramim, J.D.; Yammoto, J.D. Comparative toxicity of an acetogenin-based extract and commercial pesticides against citrus red mite. Exp. Appl. Acarol. 2014, 64, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Dimetry, N.Z.; El-Gengaihi, S.; Reda, A.S.; Amer, S.A.A. Toxicity of some compounds isolated from Abrus precatorius L. seeds towards the two-spotted spider mite Tetranychus urticae Koch. Acarologia 1990, 31, 361–366. [Google Scholar]
- Dimetry, N.Z.; Amer, S.A.A.; El-Gengaihi, S. Toxicological evaluation and biological potency of petroleum ether extract of two plants and their isolates towards the two spotted spider mite “Tetranychus urticae” Koch. Acarologia 2003, 43, 67–73. [Google Scholar]
- Hosny, A.H.; Keratum, A.Y.; Hasan, N.E. Comparative efficiency of pesticides and some predators to control spider mites: II Biological and behavioral characteristics of predators Stethorus gilvifrons, Amblyseius gossipi and Phytoseiulus macropili and their host two spotted spider mite, Tetranychus urticae under some chemicals treatments. J. Plant Prot. Path. Mansoura Univ. 2010, 1, 1065–1085. [Google Scholar] [CrossRef]
- Fetoh, B.E.A.; Al-Shammery, K.A. Acaricidal ovicial and repellent activities of some plant extracts on the date palm dust mite, Oligonychus afrasiaticus Meg. (Acari: Tetranychidae). Int. J. Environ. Sci. Eng. 2011, 2, 45–52. [Google Scholar]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Koul, O. Antifeedant phytochemicals in insect management. In Ecofriendly Pest Management for Food Security; Omkar, Ed.; Elsevier Inc.: London, UK, 2016; pp. 525–544. [Google Scholar]
- Abiodun, J.; Efe-Imafidon, A.; Benson-Oluwafemi, A.; Ajibola-Aluko, P. Efficacy of selected plant extracts in the management of tomato early blight disease caused by Alternaria solani. Asian J. Plant Pathol. 2017, 11, 48–52. [Google Scholar] [CrossRef]
- Moreno-Limón, S.; González-Solís, L.N.; Salcedo-Martínez, S.M.; Cárdenas-Avila, M.L.; Perales-Ramírez, A. Efecto antifúngico de extractos de gobernadora (Larrea tridentata L.) sobre la inhibición in vitro de Aspergillus flavus y Penicillium sp. Polibotánica 2011, 32, 193–205. [Google Scholar]
- Jasso de Rodríguez, D.; Trejo-González, F.A.; Rodríguez-García, R.; Díaz-Jiménez, M.L.V.; Sáenz-Galindo, A.; Hernández-Castillo, F.D.; Villarreal-Quintanilla, J.A.; Peña-Ramos, F.M. Antifungal activity in vitro of Rhus muelleri against Fusarium oxysporum f. sp. lycopersici. Ind. Crops Prod. 2015, 75, 150–158. [Google Scholar] [CrossRef]
- Sahgal, G.; Ramanathan, S.; Sasidharan, S.; Mordi, M.N.; Ismail, S.; Mansor, S.M. Phytochemical and antimicrobial activity of Swietenia mahagoni crude methanolic seed extract. Trop. Biomed. 2009, 26, 274–279. [Google Scholar]
- Usman, H.; Abdulrahman, F.I.; Usman, A. Qualitative phytochemical screening and in vitro antimicrobial effects of methanol stem bark extract of Ficus thonningii (Moraceae). Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, A. Demographic toxicology as a method for studying the dicofol-twospotted spider mite (Acari: Tetranychidae) system. J. Econ. Entomol. 1983, 76, 239–242. [Google Scholar] [CrossRef]
- Kramer, W.L.; Mulla, S. Oviposition attractants and repellents of mosquitoes: Oviposition responses of Culex1 mosquitoes to organic infusions. Environ. Entomol. 1979, 8, 1111–1117. [Google Scholar] [CrossRef]
- Hussey, N.W.; Parr, W.J. The effect of glasshouse red spider mite (Tetranychus urticae Koch) on the yield of cucumbers. J. Hortic. Sci. 1963, 38, 255–263. [Google Scholar] [CrossRef]
- Nachman, G.; Zemek, R. Interactions in a tritrophic acarine predator-prey metapopulation system III: Effects of Tetranychus urticae (Acari: Tetranychidae) on host plant condition. Exp. Appl. Acarol. 2002, 26, 27–42. [Google Scholar] [CrossRef]
- Birch, L.C. The intrinsic rate of increase of an insect population. J. Anim. Ecol. 1948, 17, 15–26. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971; p. 331. [Google Scholar]
- SAS Institute. SAS/STAT 9.1: User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2002; pp. 3703–3796. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Bioactive Compound | Test | Bioactive Compound | Test | ||
|---|---|---|---|---|---|
| Tannins | + | FeCl3 (catechol) | Flavonoids | + | Shinoda’s |
| + | K3[Fe(CN)6] | + | NaOH | ||
| + | Gelatine | Saponins | + | Foam | |
| Carbohydrates | + | Molisch’s | Sterols and terpenes | + | Burchard’s |
| Carotenoids | + | H2SO4 | Glycosides | − | Gringnard’s |
| + | FeCl3 | Quinones | + | Börntraguer’s | |
| Coumarins | − | NH4OH | + | H2SO4 | |
| Free reducing sugars | + | Fehling | Soluble starch | + | KOH and H2SO4 |
| Alkaloids | + | Benedict’s | + | Gelatine | |
| − | Dragendorff | Purins | + | HCI | |
| − | Sonneschain’s | Polysaccharides | − | Lugol’ |
| Treatment | Eggs/Female ± SE * | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| Control | 5.20 ± 0.51 a ** | 6.60 ± 0.12 a | 6.87 ± 0.62 a |
| 5 µg/mL | 5.17 ± 0.18 a | 5.07 ± 0.26 b | 4.97 ± 0.12 ab |
| 10 µg/mL | 5.10 ± 0.23 a | 4.23 ± 0.23 bc | 3.97 ± 0.22 bc |
| 50 µg/mL | 4.43 ± 0.23 a | 3.93 ± 0.22 cd | 3.57 ± 0.88 bc |
| 100 µg/mL | 3.03 ± 0.15 b | 3.07 ± 0.23 de | 3.07 ± 0.18 cd |
| 250 µg/mL | 2.50 ± 0.12 bc | 2.1 ± 0.31 ef | 2.10 ± 0.21 de |
| 500 µg/mL | 1.53 ± 0.15 cd | 1.37 ± 0.03 f | 1.13 ± 0.17 e |
| 1000 µg/mL | 1.27 ± 0.18 d | 1.17 ± 0.07f | 0.80 ± 0.30 e |
| EC50 (IC95) | EC90 (IC95) | b ± EE | χ2 |
| 308.16 | 2982 | 2.23 ± 0.43 | 26.74 *** |
| (184.06–430.17) | (1718–9240) | ||
| Treatment | Percentage of Feeding Damage ± SE * | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| Control | 10.00 ± 0.58 a | 17.33 ± 1.45 a | 29.00 ± 1.00 a |
| 5 µg/mL | 8.67 ± 0.33 ab | 14.67 ± 0.33 ab | 21.00 ± 1.00 b |
| 10 µg/mL | 8.67 ± 0.88 ab | 14.67 ± 1.86 ab | 20.00 ± 0.58 b |
| 50 µg/mL | 8.00 ± 0.58 abc | 12.00 ± 1.53 bc | 19.67 ± 1.76 b |
| 100 µg/mL | 7.00 ± 0.58 abc | 8.67 ± 0.33 cd | 15.33 ± 1.45 bc |
| 250 µg/mL | 6.33 ± 0.67 bc | 7.00 ± 1.00 cd | 12.67 ± 1.15 cd |
| 500 µg/mL | 5.00 ± 0.58 cd | 6.67 ± 0.33 d | 8.67 ± 0.88 de |
| 1000 µg/mL | 2.67 ± 0.88 d | 4.33 ± 0.33 d | 5.67 ± 0.88 e |
| Treatment | Demographic Parameters | ||
|---|---|---|---|
| Growth Rate * | Finite Rate of Growth | Doubling Time | |
| Control | 0.9929 ± 0.01 a | 2.6991 ± 0.01 a | 0.6982 ± 0.00 f |
| 5 µg/mL | 0.9283 ± 0.00 b | 2.5303 ± 0.01 b | 0.7467 ± 0.00 ef |
| 10 µg/mL | 0.8865 ± 0.01 bc | 2.4268 ± 0.02 c | 0.7821 ± 0.01 ef |
| 50 µg/mL | 0.8532 ± 0.00 c | 2.3473 ± 0.01 c | 0.8124 ± 0.00 de |
| 100 µg/mL | 0.7729 ± 0.01 d | 2.1662 ± 0.01 d | 0.8969 ± 0.01 d |
| 250 µg/mL | 0.6796 ± 0.02 e | 1.9736 ± 0.03 e | 1.0212 ± 0.03 c |
| 500 µg/mL | 0.5376 ± 0.02 f | 1.7125 ± 0.03 f | 1.2929 ± 0.05 b |
| 1000 µg/mL | 0.4809 ± 0.01 g | 1.6175 ± 0.01 f | 1.4421 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Zepeda, F.; Heinz-Castro, R.T.Q.; Olazaran-Santibañez, F.E.; Ordaz-Silva, S.; Pedro-Méndez, J.G.; Chacón-Hernández, J.C. Evaluation of Ethanolic Powdered Extract of Magnolia tamaulipana Vazquez against Oligonychus punicae Hirst (Trombidiformes: Tetranychidae). Plants 2022, 11, 1711. https://doi.org/10.3390/plants11131711
Reyes-Zepeda F, Heinz-Castro RTQ, Olazaran-Santibañez FE, Ordaz-Silva S, Pedro-Méndez JG, Chacón-Hernández JC. Evaluation of Ethanolic Powdered Extract of Magnolia tamaulipana Vazquez against Oligonychus punicae Hirst (Trombidiformes: Tetranychidae). Plants. 2022; 11(13):1711. https://doi.org/10.3390/plants11131711
Chicago/Turabian StyleReyes-Zepeda, Francisco, Rapucel Tonantzin Quetzalli Heinz-Castro, Fabian Eliseo Olazaran-Santibañez, Salvador Ordaz-Silva, José Guadalupe Pedro-Méndez, and Julio César Chacón-Hernández. 2022. "Evaluation of Ethanolic Powdered Extract of Magnolia tamaulipana Vazquez against Oligonychus punicae Hirst (Trombidiformes: Tetranychidae)" Plants 11, no. 13: 1711. https://doi.org/10.3390/plants11131711