
Allelopathy and Allelochemicals of Leucaenaleucocephala as an Invasive Plant Species


Abstract
:1. Introduction
2. Field Observation
3. Allelopathy
3.1. Plant Extract
3.2. Leachate
3.3. Plant Litter and Residue
3.4. Rhizosphere Soil and Root Exudate
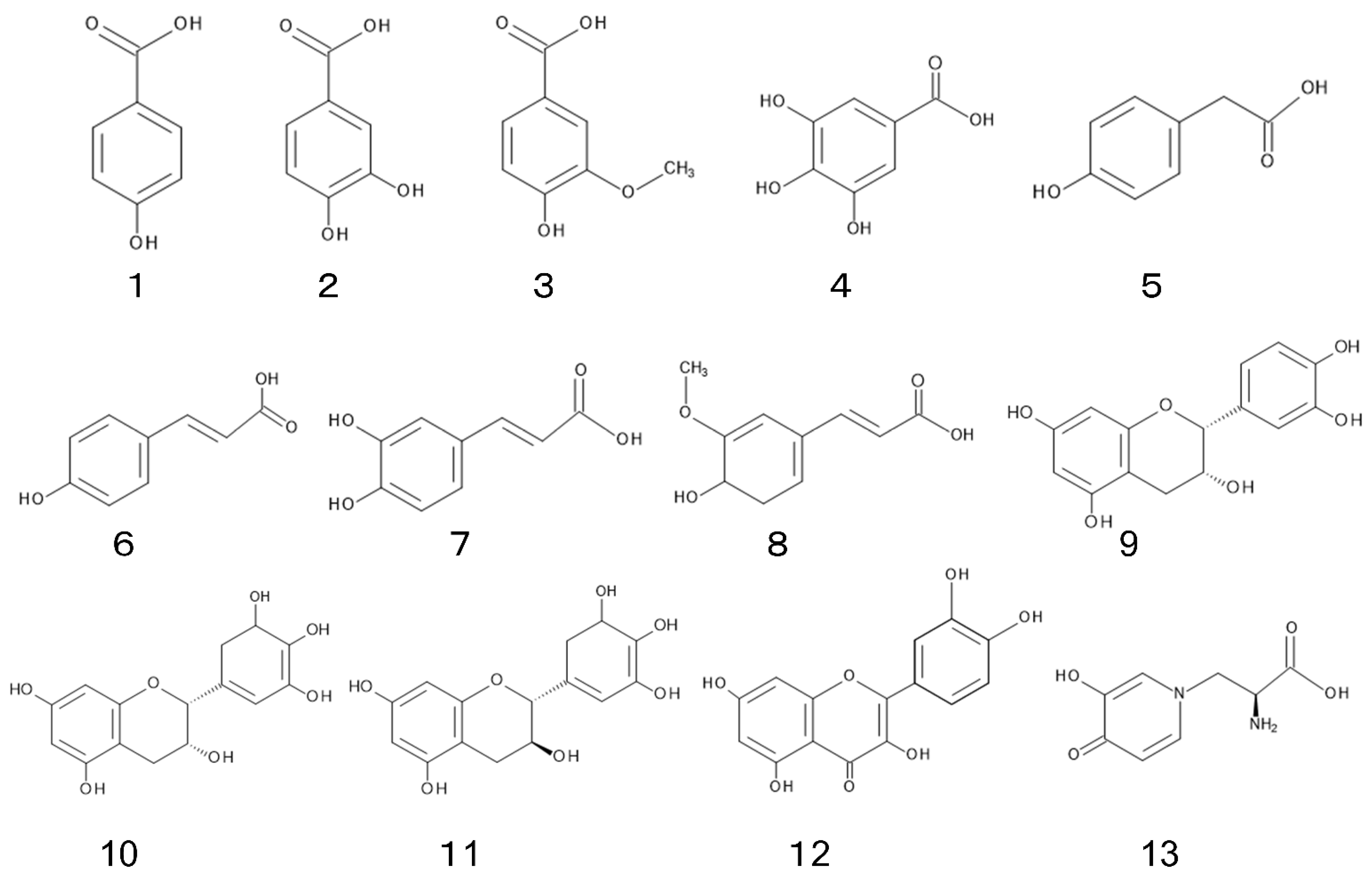
4. Allelochemical
4.1. Phenolic Acid
4.2. Flavonoid
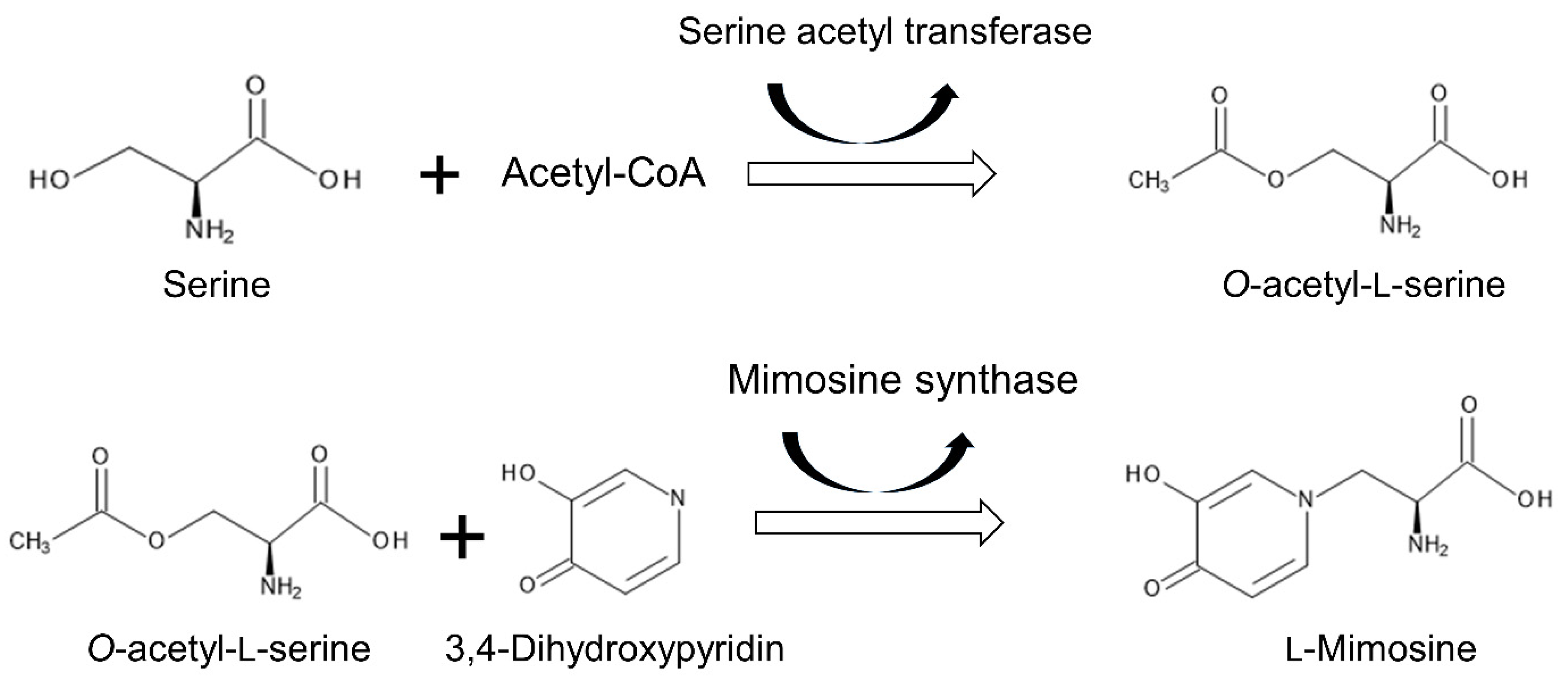
4.3. Mimosine
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Invasive Species Compendium. Leucaena leucocephala (Leucaena). Available online: https://www.cabi.org/ISC/abstract/20127801953 (accessed on 8 April 2022).
- Global Invasive Species Database. Species Profile: Leucaena leucocephala. Available online: http://www.iucngisd.org/gisd/speciesname/Leucaena+leucocephala (accessed on 8 April 2022).
- National Research Council. Leucaena: Promising Forage and Tree Crop for the Tropics; The National Academies Press: Washington, DC, USA, 1984; pp. 1–129. [Google Scholar] [CrossRef]
- Van den Beldt, R.J.; Brewbaker, J.L. Leucaena Wood Production and Use; Nitrogen Fixing Tree Association: Waimanalo, HI, USA, 1985; pp. 1–50. [Google Scholar]
- Verma, S. A review study on Leucaena leucocephala: A multipurpose tree. Int. J. Sci. Res. Sci. Eng. Technol. 2016, 2, 103–105. [Google Scholar]
- Abair, A.; Hughes, C.E.; Bailey, C.D. The evolutionary history of Leucaena: Recent research, new genomic resources and future directions. Trop. Grassl. Forrajes Trop. 2019, 7, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Sohtome, Y.; Tokunaga, T.; Ueda, K.; Yamamura, S.; Ueda, M. Leaf-closing substance in Leucaena leucocephala. Biosci. Biotechnol. Biochem. 2002, 66, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Walton, C. Leucaena (Leucaena leucocephala) in Queensland; Department of Natural Resources and Mines: Brisbane, QLD, Australia, 2003; pp. 1–51. [Google Scholar]
- Olckers, T. Biological control of Leucaena leucocephala (Lam.) de Wit (Fabaceae) in South Africa: A tale of opportunism, seed feeders and unanswered questions. Afr. Entomol. 2011, 19, 356–365. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Hegde, N.; Hutton, E.M.; Jones, R.J.; Lowry, J.B.; Moog, F.; van den Beldt, R. Leucaena—Forage Production and Use; Nitrogen Fixing Tree Association: Waimanalo, HI, USA, 1985; pp. 1–39. [Google Scholar]
- Campbell, S.; Vogler, W.; Brazier, D.; Vitelli, J.; Brooks, S. Weed Leucaena and its significance, implications and control. Trop. Grassl. Forrajes Trop. 2019, 7, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.W.; Ferguson, T.U.; Neckels, F.A.; Archibald, K.A.E. The nutritive value and forage productivity of Leucaena leucocephala. Anim. Feed Sci. Technol. 1996, 60, 29–41. [Google Scholar] [CrossRef]
- Wanapat, M.; Kang, S.; Polyorach, S. Development of feeding systems and strategies of supplementation to enhance rumen fermentation and ruminant production in the tropics. J. Anim. Sci. Biotechnol. 2013, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, A.; Gasco, L.; Parisi, G.; Danieli, P.P. A multipurpose Leguminous plant for the mediterranean countries: Leucaena leucocephala as an alternative protein source: A review. Animals 2021, 11, 2230. [Google Scholar] [CrossRef]
- Durango, S.G.; Barahona, R.; Bolívar, D.; Chirinda, N.; Arango, J. Feeding strategies to increase nitrogen retention and improve rumen fermentation and rumen imcrobial population in beef steers fed with tropical forages. Sustainability 2021, 13, 10312. [Google Scholar] [CrossRef]
- Anupam, K.; Swaroop, V.; Deepika, L.P.S.; Bist, V. Turning Leucaena leucocephala bark to biochar for soil application via statistical modelling and optimization technique. Ecol. Eng. 2015, 82, 26–39. [Google Scholar] [CrossRef]
- Jha, P.; Neenu, S.; Rashmi, I.; Meena, B.P.; Jatav, R.C.; Lakaria, B.L.; Biswas, A.L.; Singh, M.; Patra, A.K. Ameliorating effects of Leucaena biochar on soil acidity and exchangeable Ions. Commun. Soil Sci. Plant Anal. 2016, 47, 1252–1262. [Google Scholar] [CrossRef]
- Dalzell, S.A. Leucaena cultivars—Current releases and future opportunities. Trop. Grass. 2019, 7, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Khanna, N.K.; Shukla, O.P.; Gogate, M.G.; Narkhede, S.L. Leucaena for paper industry in Gujarat, India: Case study. Trop. Grassl. Forrajes Trop. 2019, 7, 200–209. [Google Scholar] [CrossRef]
- Mac Dicken, K.G. Nitrogen Fixing Trees for Wastelands; FAO Regional Office for Asia and the Pacific—Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 1988; pp. 1–104. [Google Scholar]
- Hansted, A.L.S.; Nakashima, G.T.; Martins, M.P.; Yamamoto, H.; Yamaji, F.M. Comparative analyses of fast growing species in different moisture content for high quality solid fuel production. Fuel 2016, 184, 180–184. [Google Scholar] [CrossRef]
- Nguyen, M.P.; Vaast, P.; Pagella, T.; Sinclair, F. Local knowledge about ecosystem services provided by trees in coffee agroforestry practices in northwest Vietnam. Land 2020, 9, 486. [Google Scholar] [CrossRef]
- Yu, G.; Huang, H.Q.; Wang, Z.; Brierley, G.; Zhang, K. Rehabilitation of a debris-flow prone mountain stream in southwestern China—Strategies, effects and implications. J. Hydrol. 2012, 414–415, 231–243. [Google Scholar] [CrossRef]
- Adhikary, P.P.; Hombegowda, H.C.; Barman, D.; Jakhar, P.; Madhu, M. Soil erosion control and carbon sequestration in shifting cultivated degraded highlands of eastern India: Performance of two contour hedgerow systems. Agrof. Syst. 2017, 91, 757–771. [Google Scholar] [CrossRef]
- Ishihara, K.L.; Honda, M.D.H.; Bageel, A.; Borthakur, D. Leucaena leucocephala: A leguminous tree suitable for eroded habitats of Hawaiian Islands. In Ravine Lands: Greening for Livelihood and Environmental Security; Dagar, J., Singh, A., Eds.; Springer: Singapore, 2018; pp. 413–431. [Google Scholar]
- Standley, P.C. Trees and Shrubs of Mexico, Contributions from the United State National Herbarium; Government Printing Office: Washington, DC, USA, 1922; pp. 1–1721. [Google Scholar]
- Badalamenti, E.; Pasta, S.; Sala, G.; Catania, V.; Quatrini, P.; La Mantia, T. The paradox of the alien plant Leucaena leucocephala subsp. glabrata (Rose) S. Zárate in Sicily: Another Threat for the Native Flora or a Valuable Resource? Int. J. Plant Biol. 2020, 11, 8637. [Google Scholar] [CrossRef]
- Chou, C.H.; Kuo, Y.L. Allelopathic research of subtropical vegetation in Taiwan. J. Chem. Ecol. 1986, 12, 1431–1448. [Google Scholar] [CrossRef]
- Hata, K.; Suzuki, J.I.; Kachi, N. Fine-scale spatial distribution of seedling establishment of the invasive plant, Leucaena leucocephala, on an oceanic island after feral goat extermination. Weed Res. 2010, 50, 472–480. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Hata, K.; Suzuki, J.I.; Kachi, N. Effects of an alien shrub species, Leucaena leucocephala, on establishment of native mid-successional tree species after disturbance in the national park in the Chichijima island, a subtropical oceanic island. Tropics 2007, 16, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Hwang, C.Y.; Hsu, L.M.; Liou, Y.J.; Wang, C.Y. Distribution, growth, and seed germination ability of lead tree (Leucaena leucocephala) plants in Penghu Islands, Taiwan. Weed Technol. 2010, 24, 574–582. [Google Scholar] [CrossRef]
- Marler, T.N.; Nirmala Dongol, N.; Cruz, G.M. Leucaena leucocephala and adjacent native limestone forest habitats contrast in soil properties on Tinian Island. Commu. Integr. Biol. 2016, 9, e1212792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mello, T.J.; Oliveira, A.A. Making a bad situation worse: An invasive species altering the balance of interactions between local species. PLoS ONE 2016, 11, e0152070. [Google Scholar] [CrossRef] [Green Version]
- Njunge, J.T.; Kaholongo, I.K.; Amutenya, M.; Hove, K. Invasiveness and biomass production of Leucaena leucocephala under harsh ecological conditions of north-central Namibia. J. Trop. For. Sci. 2017, 29, 297–304. [Google Scholar]
- Idol, T. A short review of Leucaena as an invasive species in Hawaii. Trop. Grassl. Forrajes Trop. 2019, 7, 290–294. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.C.; Chen, C.T.; Jump, A.S. Forest disturbance leads to the rapid spread of the invasive Leucaena leucocephala in Taiwan. J. Rakuno Gakuen Univ. 2014, 38, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, B.T.; Van Bloem, S.J. Subtropical dry forest regeneration in grass-invaded areas of Puerto Rico: Understanding why Leucaena leucocephala dominates and native species fail. For. Ecol. Manag. 2012, 267, 253–261. [Google Scholar] [CrossRef]
- Marques, A.R.; Costa, C.F.; Atman, A.P.F.; Garcia, Q.S. Germination characteristics and seedbank of the alien species Leucaena leucocephala (Fabaceae) in Brazilian forest: Ecological implications. Weed Res. 2014, 54, 576–583. [Google Scholar] [CrossRef]
- Egler, F.E. Indigene vs. alien in the development of arid vegetation in Hawaiian vegetation. Ecology 1942, 23, 14–23. [Google Scholar] [CrossRef]
- Hatheway, W.H. Composition of certain native dry forests: Mokuleia, Oahu, T.H. Ecol. Monogr. 1952, 22, 153–168. [Google Scholar] [CrossRef]
- Colón, S.M. Recovery of a subtropical dry forest after abandonment of different land. Biotropica 2006, 38, 354–364. [Google Scholar] [CrossRef]
- Francis, J.K.; Parrotta, J.A. Vegetation response to grazing and planting of Leucaena leucocephala in a Urochloa maximum-dominated grassland in Puerto Rico. Caribb. J. Sci. 2006, 42, 67–74. [Google Scholar]
- Yoshida, K.; Oka, S. Impact of biological invasion of Leucaena leucocephala on successional pathway and species diversity of secondary forest on Hahajima Island, Ogasawara (Bonin) Islands, northwestern Pacific. Jpn. J. Ecol. 2000, 50, 111–119. [Google Scholar]
- Jurado, E.; Flores, J.; Navar, J.; Jiménez, J. Seedling establishment under native tamaulipan thornscrub and Leucaena leucocephala plantation. For. Ecol. Manag. 1998, 105, 151–157. [Google Scholar] [CrossRef]
- Yoshida, K.; Oka, S. Invasion of Leucaena leucocephala and its effects on the native plant community in the Ogasawara (Bonin) Islands. Weed Technol. 2004, 18, 1371–1375. [Google Scholar] [CrossRef]
- Sandhu, J.; Sinha, M.; Ambasht, R.S. Nitrogen release from decomposing litter of Leucaena leucocephala in the dry tropics. Soil Biol. Biochem. 1990, 22, 859–863. [Google Scholar] [CrossRef]
- Valiente, C.A. The invasion ecology of Leucaena leucocephala on Moorea, French Polynesia. Biol. Geomorphol. Trop. Islands 2010, 19, 65–72. [Google Scholar]
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen losses from the soil/plant system: A review. Ann. Appl. Biol. 2013, 162, 145–173. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef] [PubMed]
- Boaprem, P. Allelopathic effects of leucaena leaves extract (Leucaena leucocephala (Lam.) de Wit) on the growth of rice (Oryza sativa L.), wrinkle duck-beak (Ischaemum rugosum Salisb), and mung bean (Vigna radiata (L.) R. Wilczek). Songklanakarin. J. Sci. Technol. 2019, 41, 619–623. [Google Scholar]
- Ishak, M.S.; Sahid, I. Allelopathic effects of the aqueous extract of the leaf and seed of Leucaena leucocephala on three selected weed species. AIP Conf. Proc. 2016, 1744, 020029. [Google Scholar]
- Sahoo, U.K.; Upadhyaya, K.; Meitei, C.B. Allelopathic effects of Leucaena leucocephala and Tectona grandis on germination and growth of maize. Allelopath. J. 2007, 20, 135–144. [Google Scholar]
- Pires, N.M.; Prates, H.T.; Filho, I.A.P.; de Oliveira, R.S., Jr.; de Faria, T.C.L. Allelopathic activity of Leucaena on weed species. Sci. Agric. 2001, 58, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Mauli, M.M.; Fortes, A.M.T.; Rosa, D.M.; Piccolo, G.; Marques, D.S.; Corsato, J.M.J.; Leszczynski, R. Leucaena allelopathy on weeds and soybean seed germination. Semin. Ciênc. Agrár. 2009, 30, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Pires, N.M.; de Souza, I.R.P.; Prates, H.T.; Faria, T.C.L.; Filho, I.A.P. Effect of Leucaena aqueous extract on the development, mitotic index, and peroxidase activity in maize seedlings. R. Bras. Fisiol. Veg. 2001, 13, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.; Alamri, S.; Al-Rumman, S.; Moustafa, M. Allelopathic and cytotoxic effects of medicinal plants on vegetable crop pea (Pisum sativum). Cytologia 2018, 83, 277–282. [Google Scholar] [CrossRef]
- John, J.; Sreekumar, K.M.; Rekha, P. Allelopathic effects of leaf leachates of multipurpose trees on vegetables. Allelopath. J. 2007, 19, 507–516. [Google Scholar]
- Maiti, P.P.; Bhakat, R.K.; Bhattacharjee, A. Allelopathic effects of Lantana camara on physio-biochemical parameters of Mimosa pudica seeds. Allelopath. J. 2008, 22, 59–67. [Google Scholar]
- Kalpana, P.; Navin, M.K. Assessment of allelopathic potential of Leucaena leucocephala (Lam) De Vit on Raphanus sativus L. Int. J. Sci. Res. Publ. 2015, 5, 1–3. [Google Scholar]
- Chai, T.T.; Ooh, K.F.; Ooi, P.W.; Chue, P.S.; Wong, F.C. Leucaena leucocephala leachate compromised membrane integrity, respiration and antioxidative defense of water hyacinth leaf tissues. Bot. Stud. 2013, 54, 8. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, T.; Hoque, A.T.M.R.; Hossain, M.K. Allelopathic effects of Leucaena leucocephala leaf litter on some forest and agricultural crops grown in nursery. J. For. Res. 2008, 19, 298–302. [Google Scholar] [CrossRef]
- Ishak, M.S.; Ismail, B.S.; Yusoff, N. Allelopathic potential of Leucaena leucocephala (Lam.) de Wit on the germination and seedling growth of Ageratum conyzoides L., Tridax procumbens L. and Emilia sonchifolia (L.) DC. Allelopath. J. 2016, 37, 109–122. [Google Scholar]
- Kamara, A.Y.; Akobundu, I.O.; Sanginga, N.; Jutzi, S.C. Effects of mulch from 14 multipurpose tree species (MPTs) on early growth and nodulation of cowpea (Vigna unguiculata L.). J. Agron. Crop Sci. 1999, 182, 127–133. [Google Scholar] [CrossRef]
- Parvin, R.; Shapla, T.L.; Amin, M.H.A. Effect of leafs and Leucaena leucocephala different three depth soil on the allelopathy of agricultural crops. J. Innov. Dev. Strategy 2011, 5, 61–69. [Google Scholar]
- Sharma, P.; Chaurasia, S. Evaluation of total phenolic, flavonoid contents and antioxidant activity of Acokanthera oppositifolia and Leucaena leucocephala. Int. J. Pharmacogn. Phytochem. Res. 2014, 15, 175–180. [Google Scholar]
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202. [Google Scholar] [CrossRef]
- Dalton, B.R. The occurrence and behavior of plant phenolic acids in soil environments and their potential involvement in allelochemical interference interactions: Methodological limitations in establishing conclusive proof of allelopathy. In Principals and Practices in Plant Ecology: Allelochemical Interactions; Inderjit, Dakshini, K.M.M., Foy, C.L., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 57–74. [Google Scholar]
- Batish, D.R.; Kaur, S.; Singh, H.P.; Kohli, R.K. Role of root-mediated interactions in phytotoxic interference of Ageratum conyzoides with rice (Oryza sativa). Flora 2009, 204, 388–395. [Google Scholar] [CrossRef]
- Inderjit; Seastedt, T.R.; Callaway, R.M.; Pollock, J.L.; Kaur, J. Allelopathy and plant invasions: Traditional, congeneric, and bio-geographical approaches. Biol. Inva. 2008, 10, 875–890. [Google Scholar] [CrossRef]
- Einhellig, F.A. Mode of action of allelochemical action of phenolic compounds. In Chemistry and Mode of Action of Allelochemicals; Macías, F.A., Galindo, J.C.G., Molino, J.M.G., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 2004; pp. 217–238. [Google Scholar]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, A.J.; Ramsewak, R.S.; Smucker, A.J.; Nair, M.G. Nitrification Inhibitors from the Roots of Leucaena leucocephala. J. Agric. Food Chem. 2000, 48, 6174–6177. [Google Scholar] [CrossRef]
- Xu, Y.; Tao, Z.; Jin, Y.; Yuan, Y.; Dong, T.T.X.; Tsim, K.W.K.; Zhou, Z. Flavonoids, a potential new insight of Leucaena leucocephala foliage in ruminant health. J. Agric. Food Chem. 2018, 66, 7616–7626. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Constabel, C.P. Tannins in plant–herbivore interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef]
- Zhou, M.; Wei, L.; Sun, Z.; Gao, L.; Meng, Y.; Tang, Y.; Wu, Y. Production and transcriptional regulation of proanthocyanidin biosynthesis in forage legumes. Appl. Microbiol Biotechnol. 2015, 99, 3797–3806. [Google Scholar] [CrossRef]
- Paengkoum, S.; Petlum, A.; Purba, R.A.P.; Paengkoum, P. Protein binding affinity of various condensed tannin molecular weights from tropical leaf peel. J. Appl. Pharm. Sci. 2021, 11, 114–120. [Google Scholar]
- Serniak, L.T. Comparison of the allelopathic effects and uptake of Fallopia japonica phytochemicals by Raphanus sativus. Weed Res. 2016, 56, 97–101. [Google Scholar] [CrossRef]
- Okada, S.; Iwasaki, A.; Kataoka, I.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic activity of kiwifruit leaves and isolation of a phytotoxic substance. Sci. Hortic. 2019, 250, 243–248. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interaction. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Duke, S.O.; Blair, A.C.; Dayan, F.E.; Johnson, R.D.; Meepagala, K.M.; Cook, D.; Bajsa, J. Is (−)-catechin a novel weapon of spotted knapweed (Centaurea stoebe)? J. Chem. Ecol. 2009, 35, 141–153. [Google Scholar] [CrossRef]
- Renz, J. Mimosine. Physiol. Chem. 1936, 244, 153–158. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Hylin, J.W. Variations in mimosine content among Leucaena species and related mimosaceae. Crop Sci. 1965, 5, 348–349. [Google Scholar] [CrossRef] [Green Version]
- Smith, I.K.; Fowden, L.A. Study of mimosine toxicity in plants. J. Exp. Bot. 1996, 17, 750–761. [Google Scholar] [CrossRef]
- Soedarjo, M.; Borthakur, D. Mimosine produced by the tree-legume Leucaena provides growth advantages to some Rhizobium strains that utilize it as a source of carbon and nitrogen. Plant Soil 1996, 186, 87–92. [Google Scholar] [CrossRef]
- Nguyen, B.C.; Tawata, S. The chemistry and biological activities of mimosine: A review. Phytother. Res. 2016, 30, 1230–1242. [Google Scholar] [CrossRef]
- John, J.; Narwal, S.S. Allelopathic plants. 9. Leucaena Leucocephala (Lam.) De Wit. Allelopath. J. 2003, 12, 13–36. [Google Scholar]
- Hussain, M.I.; Gonzalez-Rodriguez, L.; Roger, M.R. Germination and growth response of four plant species to different allelochemicals and herbicides. Allelopath. J. 2008, 22, 101–110. [Google Scholar]
- Murakoshi, I.; Ikegami, F.; Hinuma, Y.; Hanma, Y. Purification and characterization of L-mimosine synthase from Leucaena leucocephala. Phytochemistry 1984, 23, 1905–1908. [Google Scholar] [CrossRef]
- Ikegami, F.; Mizuno, M.; Kihara, M.; Murakoshi, I. Enzymatic synthesis of the thyrotoxic amino acid mimosine by cysteine synthase. Phytochemistry 1990, 29, 3461–3465. [Google Scholar] [CrossRef]
- Harun-Ur-Rashid, M.; Iwasaki, H.; Parveen, S.; Oogai, S.; Fukuta, M.; Amzad Hossain, M.A.; Anai, T.; Oku, H. Cytosolic cysteine synthase switch cysteine and mimosine production in Leucaena leucocephala. Appl. Biochem. Biotechnol. 2018, 186, 613–632. [Google Scholar] [CrossRef] [PubMed]
- MetaCyc, Pathway: Mimosine Biosynthesis. Available online: https://metacyc.org/META/new-image?type=PATHWAY&object=PWY-4985 (accessed on 1 May 2022).
- Soedarjo, M.; Borthakur, D. Mimosine, a toxin produced by the tree-legume Leucaena provides a nodulation competition advantage to mimosine-degrading Rhizobium strains. Soil Biol. Biochem. 1998, 30, 1605–1613. [Google Scholar] [CrossRef]
- Xuan, T.D.; Elzaawely, A.A.; Deba, F.; Tawata, S. Mimosine in Leucaena as a potent bio-herbicide. Agron. Sustain. Dev. 2006, 26, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.D.H.; Borthakur, D. Mimosine concentration in Leucaena leucocephala under various environmental conditions. Trop. Grassl. Forrajes Trop. 2019, 7, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Corrêa, K.C.D.S.; Honda, M.D.H.; Borthakur, D.; Fett-Neto, A.G. Mimosine accumulation in Leucaena leucocephala in response to stress signaling molecules and acute UV exposure. Plant Physiol. Biochem. 2019, 135, 432–440. [Google Scholar] [CrossRef]
- Vestena, S.; Fett-Neto, A.G.; Duarte, R.C.; Ferreira, A. Regulation of mimosine accumulation in Leucaena leucocephala seedlings. Plant Sci. 2001, 161, 597–604. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Pan, Y.-J.; Chang, B.-W.; Hu, Y.-B.; Guo, X.-R.; Tang, Z.H. Ethylene induced vinblastine accumulation is related to activated expression of downstream TIA pathway genes in Catharanthus roseus. BioMed. Res. Int. 2016, 2016, 3708187. [Google Scholar]
- Wasternack, C.; Strnad, M. Jasmonate signaling in plant stress responses and development—Active and inactive compounds. New Biotech. 2016, 33, 604–613. [Google Scholar] [CrossRef]
- Williams, R.D.; Hoagland, R.E. Phytotoxicity of mimosine and albizziine on seed germination and seedling growth of crops and weeds. Allelopath. J. 2007, 19, 423–430. [Google Scholar]
- Sahid, I.; Ishak, S.M.; Bajrai, F.S.; Jansar, K.M.; Yusoff, N. Quantification and herbicidal activity of mimosine from Leucaena leucocephala (Lam.) de Wit. Trans. Sci. Technol. 2017, 4, 62–67. [Google Scholar]
- Negi, V.S.; Borthakur, D. Heterologous expression and characterization of mimosinase from Leucaena leucocephala. In Biotechnology of Plant Secondary Metabolism: Methods and Protocols, Methods in Molecular Biology; Fett-Neto, A., Ed.; Humana Press: New York, NY, USA, 2016; p. 59. [Google Scholar]
- Honda, M.D.H.; Ishihara, L.L.; Pham, D.T.; Borthakur, D. Highly expressed genes in the foliage of giant Leucaena (Leucaena leucocephala subsp. glabrata), a nutritious fodder legume in the tropics. Plant Biosyst. 2020, 154, 107–116. [Google Scholar] [CrossRef]
- Perennes, C.; Qin, L.X.; Glab, N.; Bergounioux, C. Petunia p34cdc2 protein kinase activity in G2/M cells obtained with a reversible cell cycle inhibitor, mimosine. FEBS Lett. 1993, 333, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.A.; Cook, P.R. Mimosine arrests the cell cycle after cells enter S-phase. Exp. Cell. Res. 1996, 222, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, S.; Fukumoto, Y.; Ishibashi, K.; Soeda, S.; Kubota, S.; Yuki, R.; Nakayama, Y.; Aoyama, K.; Yamaguchi, N.; Yamaguchi, N. Activation of the prereplication complex is blocked by mimosine through reactive oxygen species-activated ataxia telangiectasia mutated (ATM) protein without DNA damage. J. Biol. Chem. 2014, 289, 5730–5746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallon, A.M. Effects of mimosine on Wolbachia in mosquito cells: Cell cycle suppression reduces bacterial abundance. In Vitro Cell. Dev. Biol. Animal. 2015, 51, 958–963. [Google Scholar] [CrossRef] [Green Version]
- Prasad, M.N.; Subhashini, P. Mimosine-inhibited seed germination, seedling growth, and enzymes of Oryza sativa L. J. Chem. Ecol. 1994, 20, 1689–1696. [Google Scholar] [CrossRef]
- Andrade, A.B.; Ferrarese, M.L.L.; Teixeira, A.F.; Ferrarese-Filho, O. Mimosine-inhibited soybean (Glycine max) root growth, lignification and related enzymes. Allelopath. J. 2008, 21, 145–154. [Google Scholar]
- Ishaaya, I.; Hirashima, A.; Yablonski, S.; Tawata, S.; Eto, M. Mimosine, a nonprotein amino acid, inhibits growth and enzyme systems in Tribolium castaneum. Pestic. Biochem. Physiol. 1991, 39, 35–42. [Google Scholar] [CrossRef]
- Cavalcante, G.M.; Moreira, A.F.C.; Vasconcelos, S.D. Insecticidal potential of aqueous extracts from arboreous species against whitefly. Pesq. Agropec. Bras. 2006, 41, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, B.C.; Chompoo, J.; Tawata, S. Insecticidal and nematicidal activities of novel mimosine derivatives. Molecules 2015, 20, 16741–16756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Nuby, A.S.M.; Eman, A. Phytochemical and nematicidal activity studies of some extracts of different plant parts of Leucaena leucocephala against Meloidogyne incognita. J. Chem. Pharm. Sci. 2020, 11, 1–17. [Google Scholar]
- Mitchell, C.E.; Power, A.G. Release of invasive plants from fungal and viral pathogens. Nature 2003, 421, 625–627. [Google Scholar] [CrossRef]
- Cappuccino, N.; Carpenter, D. Invasive exotic plants suffer less herbivory than non-invasive plants. Biol. Lett. 2005, 1, 435–438. [Google Scholar] [CrossRef] [Green Version]
- Chengxu, W.; Mingxing, Z.; Xuhui, C.; Bo, Q. Review on allelopathy of exotic invasive plants. Procedia. Eng. 2011, 18, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Wang, Y.D. Secondary metabolites from Leucaena leucocephala. Chem. Nat. Compd. 2011, 47, 145–146. [Google Scholar] [CrossRef]
- Salem, A.Z.M.; Salem, M.Z.; Gonzalez-Ronquillo, M.; Camacho, L.M.; Cipriano, M. Major chemical constituents of Leucaena leucocephala and Salix babylonica leaf extracts. J. Trop. Agric. 2011, 49, 95–98. [Google Scholar]
- Herrera, R.S.; Verdecia, D.M.; Ramírez, J.L.; García, M.; Cruz, A.M. Secondary metabolites of Leucaena leucochephala. Their relationship with some climate elements, different expressions of digestibility and primary metabolites. Cub. J. Agric. Sci. 2017, 51, 107–116. [Google Scholar]
- She, L.-C.; Liu, C.-M.; Chen, C.-T.; Li, H.-T.; Li, W.J.; Chen, C.-Y. The anti-cancer and anti-metastasis effects of phytochemical constituents from Leucaena leucocephala. Biomed. Res. 2017, 28, 2893–2897. [Google Scholar]
- Deivasigamani, R. Phytochemical analysis of Leucaena leucocephala on various extracts. J. Phytopharm. 2018, 7, 480–482. [Google Scholar] [CrossRef]
- Zayed, M.Z.; Wu, A.; Sallam, S. Comparative phytochemical constituents of Leucaena leucocephala (Lam.) leaves, fruits, stem barks, and wood branches grown in Egypt using GC-MS method coupled with multivariate statistical approaches. BioResources 2019, 14, 996–1013. [Google Scholar]
- Lockwood, J.L.; Simberloff, D.; McKinney, M.L.; Von Holle, B. How many, and which, plants will invade natural areas. Biol. Inva. 2001, 3, 1–8. [Google Scholar] [CrossRef]
- Cappuccino, N.; Arnason, J.T. Novel chemistry of invasive exotic plants. Biol. Lett. 2006, 2, 189–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Target Plant Species | Inhibition | Stimulation | Condition | Reference |
|---|---|---|---|---|---|
| Plant extract | |||||
| Leaf | Lactuca sativa, Oryza sativa | Growth | Laboratory | [28] | |
| Ischaemum rugosum, Vigna radiata | Growth | Laboratory | [53] | ||
| Leaf, seed | procumbens, Emilia sonchifolia | Germination, growth | Laboratory | [54] | |
| Leaf, bark, seed | Zea mays | Germination, growth, crop yield | Laboratory, greenhouse | [55] | |
| Aerial part | Bidens pilosa, Amaranthus hybridus | Growth | Laboratory, greenhouse | [56] | |
| Ipomoea grandifolia, Arrowleaf sida, Bidens pilosa | Growth | Laboratory | [57] | ||
| Aerial part | Zea mays | Cell division | Peroxidase activity | Laboratory | [58] |
| Leaf | Pisum sativum | Cell division | Laboratory | [59] | |
| Ageratum conyzoides, Tridax | |||||
| Leachate | |||||
| Senescent leaf | Raphanus sativus | Germination, growth | Laboratory | [60,61,62] | |
| Leaf | Eichhornia crassipes | Electrolyte leakage, catalase, and ascorbate peroxidase activities | Laboratory | [63] | |
| Litter, residue | |||||
| Leaf | Albiza procera, Vigna unguiculata, Cicer arietinum, Cajanus cajan | Germination, growth | Greenhouse | [64] | |
| Decomposing leaf | Alnus formosana, Acacia confusa, Liquidambar formosana, Casuarina glauca, Mimosa pudica | Mortality | Greenhouse | [28] | |
| Liter extract | Lolium multiflorum | Germination, growth | Laboratory | [28] | |
| Liter extract | Ageratum conyzoides, Tridax procumbens, Emilia sonchifolia | Germination, growth | Laboratory | [65] | |
| Leaf mulch | Vigna unguiculata | Germination, growth, nodulation | Greenhouse | [66] | |
| Soil | |||||
| Rhizosphere soil | Ageratum conyzoides, Tridax procumbens, Emilia sonchifolia | Germination, growth | Greenhouse | [65] | |
| Vigna radiata, Glycine max | Germination, growth | Greenhouse | [67] | ||
| Soil extract | Lactuca sativa | Growth | Laboratory | [28] | |
| Root exudate | Ageratum conyzoides, Tridax procumbens, Emilia sonchifolia | Germination, growth | Laboratory | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato-Noguchi, H.; Kurniadie, D. Allelopathy and Allelochemicals of Leucaenaleucocephala as an Invasive Plant Species. Plants 2022, 11, 1672. https://doi.org/10.3390/plants11131672
Kato-Noguchi H, Kurniadie D. Allelopathy and Allelochemicals of Leucaenaleucocephala as an Invasive Plant Species. Plants. 2022; 11(13):1672. https://doi.org/10.3390/plants11131672
Chicago/Turabian StyleKato-Noguchi, Hisashi, and Denny Kurniadie. 2022. "Allelopathy and Allelochemicals of Leucaenaleucocephala as an Invasive Plant Species" Plants 11, no. 13: 1672. https://doi.org/10.3390/plants11131672