
Effect of 17β-Estradiol on Growth and Biosynthesis of Microalgae Scenedesmus quadricauda (CPCC-158) and Duckweed Lemna minor (CPCC-490) Grown in Three Different Media


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culturing Conditions
2.2. Water Chemistry
2.3. The Range of E2 Concentrations
2.4. Sampling and Analytical Methods
2.4.1. Algal Cell Counting
2.4.2. Assessment of Biomass Production
2.4.3. Assessment of Chlorophyll-a and Total Carotenoids Concentrations
2.4.4. Duckweed Assessment
2.5. Statistical Analyses
3. Results
3.1. Water Chemistry
3.2. Effect of E2 on S. quadricauda and Duckweed (Lemna minor) Growth and Biomass Production
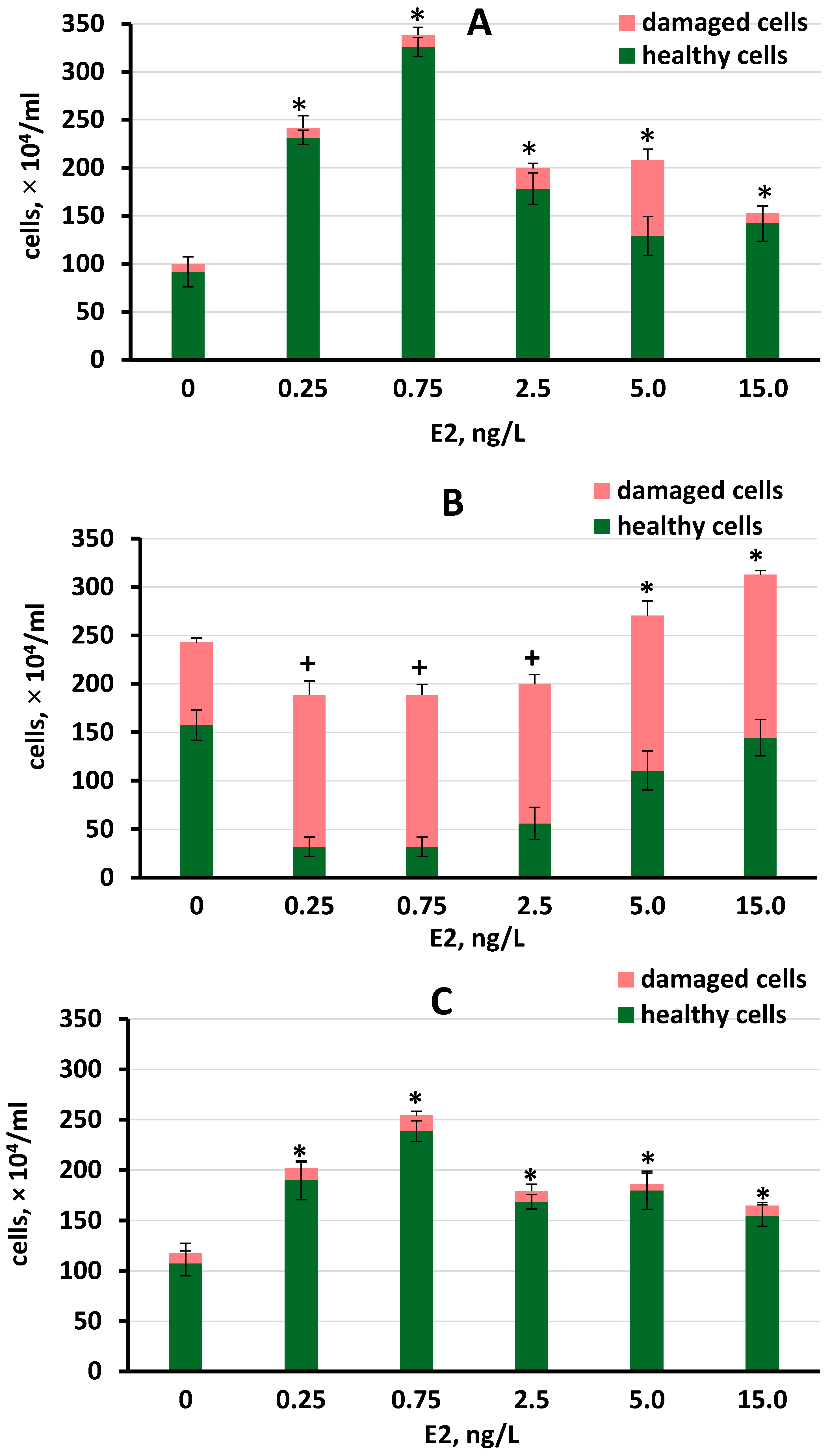
3.2.1. Growth and Biomass Production in S. quadricauda under Different E2 Concentrations
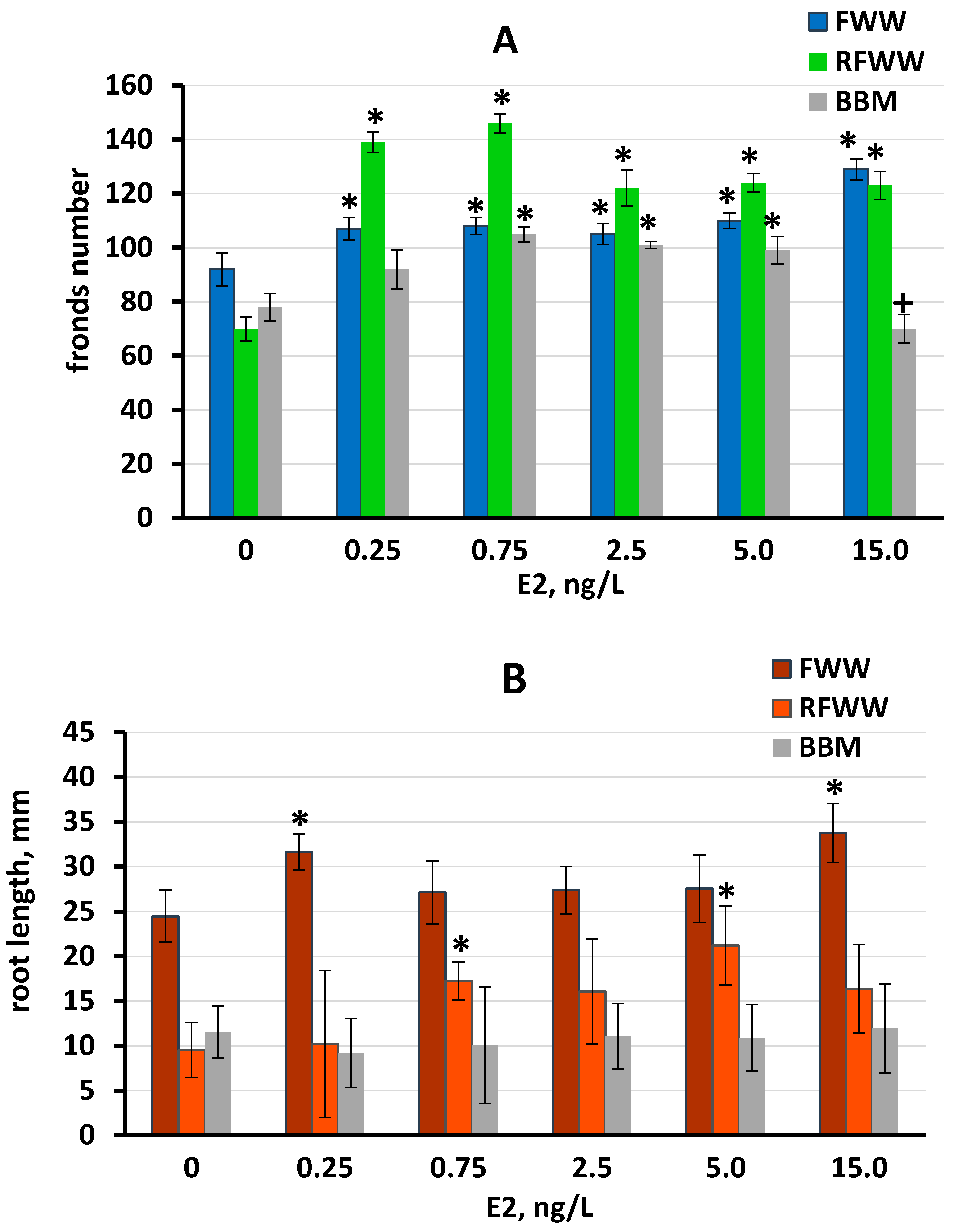
3.2.2. L. minor Growth under Different E2 Concentrations in Three Media
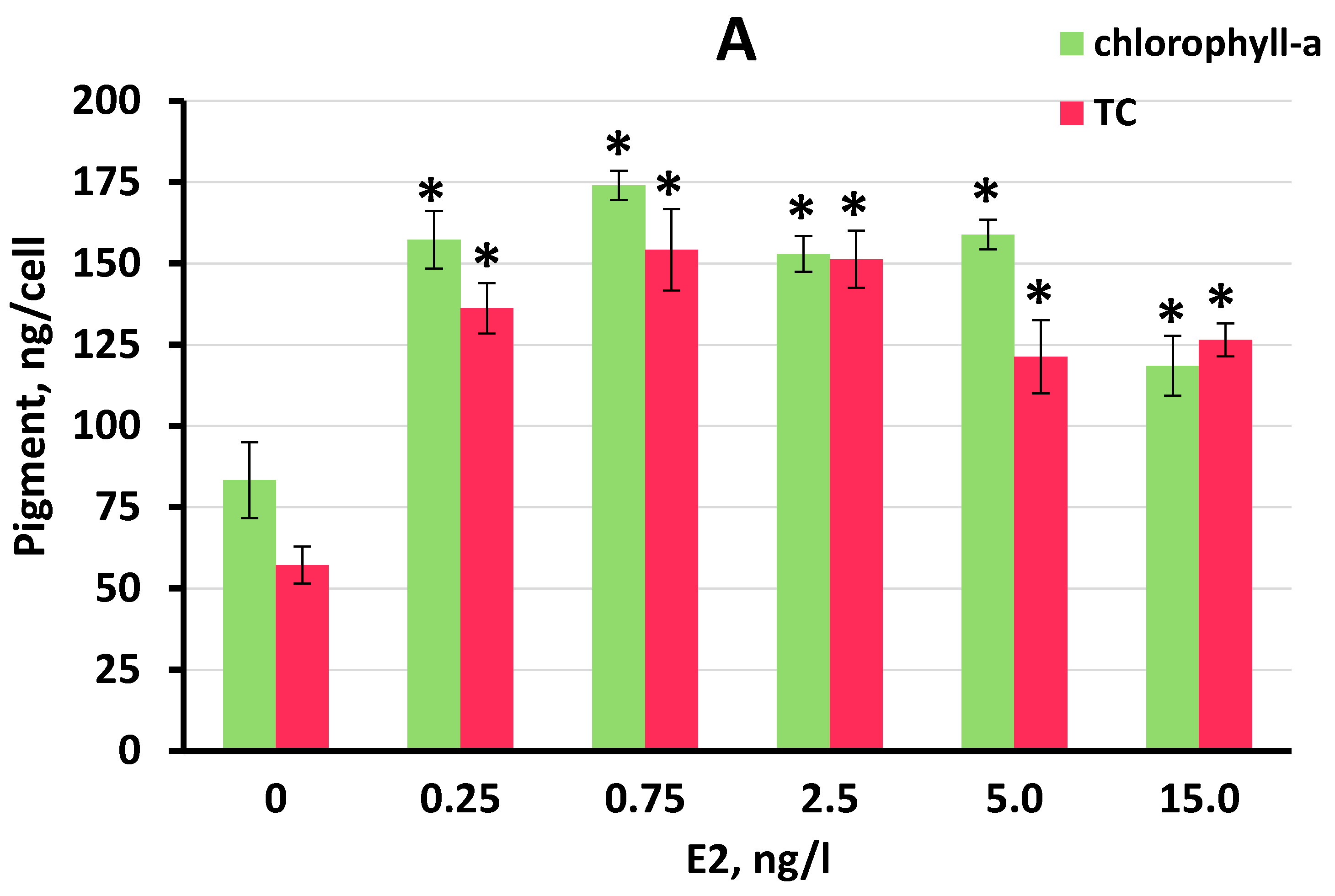
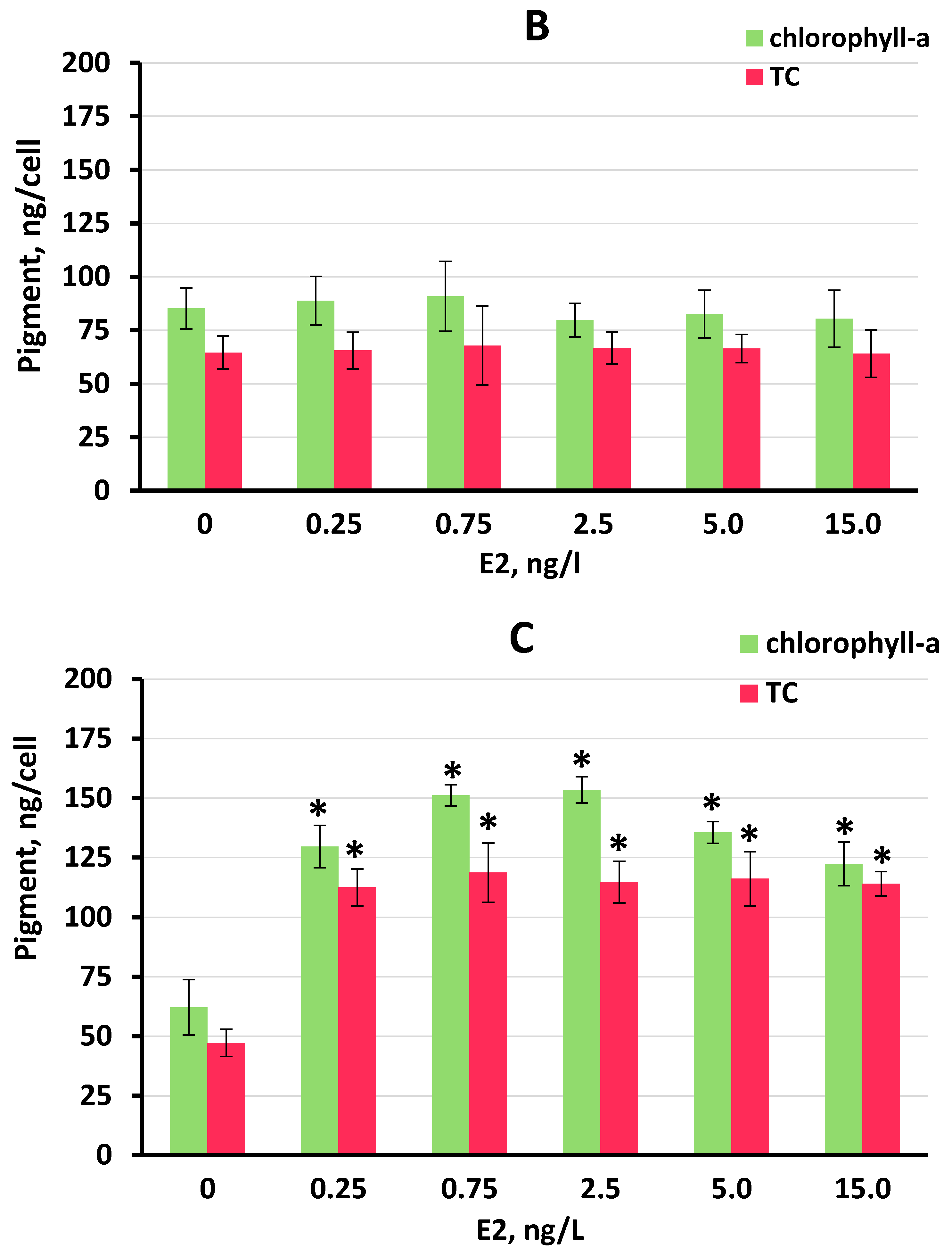
3.3. Effect of E2 on Chlorophyll-a and Total Carotenoid Production by S. quadricauda Cells
3.4. Soluble Protein Production by L. minor and S. quadricauda
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, D.S.; Kramer, J.R.; Bell, R.A. Metal speciation with organic matter. Comp. Biochem. Physiol. 2002, 133, 65–74. [Google Scholar]
- Richards, J.C.; Curtis, P.J.; Bumison, B.K.; Playle, R. Effect of natural organic matter sources on reducing metal toxicity to rainbow trout (Oncorhynchus mykiss) and on metal binding to their gills. Environ. Toxicol. Chem. 2001, 20, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wang, L.; Rousseau, D.P.L.; Lens, P.N.L. Removal of estrone, 17α-ethinylestradiol, and 17ß-estradiol in algae and Duckweed-based wastewater treatment systems. Environ. Sci. Pollut. Res. 2010, 17, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Hu, H.Y.; Gan, K.; Sun, Y.X. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, Q.; He, N.; Sun, K.; Sun, D.; Wu, X.; Duan, S. Removal and biodegradation of 17β-Estradiol and diethylstilbestrol by the freshwater microalgae Raphidocelis subcapitata. Int. J. Environ. Res. Public Health 2018, 15, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pojana, G.; Gomiero, A.; Jonkers, N.; Marcomini, A. Natural and synthetic endocrine disrupting compounds (EDCs) in water, sediment, and biota of a coastal lagoon. Environ. Int. 2007, 33, 929–936. [Google Scholar] [CrossRef]
- Zhou, W.; Li, Y.; Min, M.; Hu, B.; Chen, P.; Ruan, R. Local bioprospecting for high-lipid producing microalgal strains to be grown on concentrated municipal wastewater for biofuel production. Bioresour. Technol. 2011, 102, 6909–6919. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, Y.; Li, Q.; Li, G.; Guo, Q.; Yan, C. Estrogenic compounds and estrogenicity in surface water, sediments, and organisms from Yundang lagoon in Xiamen, China. Arch. Environ. Contam. Toxicol. 2011, 61, 93–100. [Google Scholar] [CrossRef]
- Esteban, S.; Gorga, M.; Petrovic, M.; Gonzalez-Alonso, S.; Barcelo, D.; Valcarcel, Y. Analysis and occurrence of endocrine-disrupting compounds and estrogenic activity in the surface waters of central Spain. Sci. Total Environ. 2014, 466–467, 939–951. [Google Scholar] [CrossRef]
- Barry, T.P.; Riebe, J.D.; Parrish, J.J.; Malison, J.A. Effects of 17α, 20β-Dihydroxy-4-pregnen-3-one on cortisol production by Rainbow Trout interrenal tissue in vitro. Gen. Comp. Endocrinol. 1997, 107, 172–181. [Google Scholar] [CrossRef]
- Kidd, C.E.; Kidd, M.R.; Hofmann, H.A. Measuring multiple hormones from a single water sample using enzyme immunoassays. Gen. Comp. Endocrinol. 2010, 165, 277–285. [Google Scholar] [CrossRef]
- Hala, D.; Petersen, L.H.; Martinovic, D.; Huggett, D.B. Constraints-based stoichiometric analysis of hypoxic stress on steroidogenesis in fathead minnows, Pimephales promelas. J. Exp. Biol. 2012, 215, 1753–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, A.P.; Ellis, T. Measurement of fish steroids in water—A review. Minireview. Gen. Comp. Endocrinol. 2007, 153, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Norvill, Z.N.; Toledocervantes, A.; Blanco, S.; Shilton, A.; Guieysse, B.; Muñoz, R. Photodegradation and sorption govern tetracycline removal during wastewater treatment in algal ponds. Bioresour. Technol. 2017, 232, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, K.M.; Scrimshaw, M.D.; Lester, J.N. Prediction of the bioaccumulation factors and body burden of natural and synthetic estrogens in aquatic organisms in the river systems. Sci. Total Environ. 2002, 289, 159–168. [Google Scholar] [CrossRef]
- Wang, P.; Wong, Y.S.; Tam, F.Y. Green microalgae in removal and biotransformation of estradiol and ethinylestradiol. J. Appl. Phycol. 2017, 29, 263–273. [Google Scholar] [CrossRef]
- Hom-Diaz, A.; Llorca, M.; Rodríguez-Mozaz, S.; Vicent, T.; Barceló, D.; Blánquez, P. Microalgae cultivation on wastewater digestate: β-estradiol and 17α-ethynylestradiol degradation and transformation products identification. J. Environ. Manag. 2015, 155, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Beecher, L. Assessment of 17β-Estradiol Removal from Wastewater via Abiotic and Biotic Routes and Potential Effects on Food Chain Pathways. Ph.D. Thesis, Clemson University, Clemson, SC, USA, 2013; 152p. [Google Scholar]
- Zhang, J.S.; Yang, Z.H.; Tsao, T.H. The occurrence of estrogens in relation to reproductive processes in flowering plants. Sex. Plant Reprod. 1991, 4, 193–196. [Google Scholar] [CrossRef]
- Janeczko, A.; Skoczowski, A. Mammalian sex hormones in plants. Folia Histochemica et cytobiologica 2005, 43, 71–79. [Google Scholar]
- Itagaki, E.; Iwaya, T. Purification and characterization of 17β-hydroxysteroid dehydrogenase from Cylindrocarpon Radicicola. J. Biochem. 1988, 103, 1039–1044. [Google Scholar] [CrossRef]
- Rosati, F.; Danza, G.; Guarna, A.; Cini, N.; Racchi, M.L.; Serio, M. New evidence of similarity between human and plant steroid metabolism: 5α-reductase activity in Solanum malacoxylon. Endocrinology 2003, 144, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Milanesi, L.; Monje, P.; Boland, R. Presence of estrogens and estrogen receptor-like proteins in Solanum glaucophyllum. Biochem. Biophys. Res. Commun. 2001, 289, 1175–1179. [Google Scholar] [CrossRef] [PubMed]
- Shore, L.S.; Kapulnik, Y.; Ben-Dor, B.; Fridman, Y.; Wininger, S.; Shemesh, M. Effects of estrone and 17β-estradiol on vegetative growth of Medicago sativa. Physiol. Plant. 1992, 84, 217–222. [Google Scholar] [CrossRef]
- Janeczko, A.; Filek, W. Stimulation of generative development in partly vernalized winter wheat by animal sex hormones. Acta Physiol. Plant. 2002, 24, 291–295. [Google Scholar] [CrossRef]
- Kopcewicz, J. Influence of estrogens on the auxins content in plants. Naturwissenschaffen 1970, 57, 48. [Google Scholar] [CrossRef] [PubMed]
- Kopcewicz, J.; Porazinski, Z. Effect of growth regulators, steroids and estrogen fraction from sage plants on flowering of a long-day plant, Salvia splendens, grown under non-inductive light conditions. Biol. Plant. 1974, 16, 132–135. [Google Scholar] [CrossRef]
- Geuns, J.M.C. Steroid hormones and plant growth and development. Phytochemistry 1978, 17, 1–14. [Google Scholar] [CrossRef]
- Zhong-Han, Y.; Yin, T.; Zong-Xun, C.; Tsao, T.H. The changes of steroidal sex hormone–testosterone contents in reproductive organs of Lilium davidii Duch. Acta Bot. Sin. 1994, 36, 215–220. [Google Scholar]
- Czerpak, R.; Szamrej, I.K. The Effect of β-estradiol and corticosteroids on chlorophylls and carotenoids content in Wolffia arrhiza (L.) Wimm. (Lemnaceae) Growing in Municipal Bialystok Tap Water. Pol. J. Environ. Stud. 2003, 12, 677–684. [Google Scholar]
- Szamrej, I.K.; Czerpak, R. The effect of sex steroids and corticosteroids on the content of soluble proteins, nucleic acids and reducing sugars in Wolffia arrhiza (L.) Wimm. (Lemnaceae). Pol. J. Environ. Stud. 2004, 13, 565–571. [Google Scholar]
- Czygan, J.C. Blütenbildung bei Lemna minor nach Zusatz von Oestrogen. Naturwissenschaften 1962, 49, 285–286. [Google Scholar] [CrossRef]
- Sayegh, A.; Greppin, H. Chlorella rubescens essai de synchronisation et mise en evidence de rythmes endogens. Arch. Sci. Geneve 1973, 8, 6–18. [Google Scholar]
- Bajguz, A.; Czerpak, R. Metabolic activity of estradiol in Chlorella vulgaris Beijerinck (Chlorophytaceae) Part 1. Content of photosynthetic pigments. Pol. Arch. Hydrobiol. 1996, 43, 421–426. [Google Scholar]
- Bajguz, A.; Czerpak, R. Metabolic activity of estradiol in Chlorella vulgaris Beijerinck (Chlorophyceae) Part II. Content of the cellular sugar and protein accumulation. Pol. Arch. Hydrobiol. 1996, 43, 427–430. [Google Scholar]
- Czerpak, R.; Szamrej, I.K. Metabolitic activity of 11-deoxycorticosterone and prednisolone in the alga Chlorella vulgaris Beijerinck. Acta Soc. Bot. Pol. 2000, 69, 25. [Google Scholar] [CrossRef]
- Czerpak, R.; Bajguz, A.; Iwaniuk, D. Comparison of the influence of hydrocortisone and progesterone on the content of protein and sugar in the green alga Chlorella vulgaris Beijerinck. Ecohydrol. Hydrobiol. 2001, 1, 473. [Google Scholar]
- Kozlova, T.A.; Hardy, B.P.; Levin, D.B. Effect of Fish Steroids 17β-estradiol and 17,20β-dihydroxy-4-pregnen-3-one on Growth, Accumulation of Pigments, and Fatty Acid Profiles in the Microalgae Scenedesmus quadricauda (CPCC-158). Renew. Energy 2019, 148, 798–806. [Google Scholar] [CrossRef]
- Kim, M.K.; Park, J.W.; Park, C.S.; Kim, S.J.; Jeune, K.H.; Chang, M.U.; Acreman, J. Enhanced production of Scenedesmus spp. (green microalgae) using a new medium containing fermented swine wastewater. Bioresour. Technol. 2007, 98, 2220–2228. [Google Scholar] [CrossRef]
- Goswami, R.D.; Kalita, M.C. Scenedesmus dimorphus and Scenedesmus quadricauda: Two potent indigenous microalgae strains for biomass production and CO2 mitigation—A study on their growth behavior and lipid productivity under different concentration of urea as nitrogen source. J. Algal Biomass Utln. 2011, 2, 42–49. [Google Scholar]
- Mata, T.; Meloa, A.C.; Meireles, S.; Mendes, A.M.; Martins, A.A.; Caetano, N.C. Potential of microalgae Scenedesmus obliquus grown in brewery wastewater for biodiesel production. Chem. Eng. Trans. 2013, 32, 901–907. [Google Scholar]
- Leng, R.A.; Stambolie, J.H.; Bell, R. Duckweed—A potential high-protein feed resource for domestic animals and fish. Livest. Res. Rural Dev. 1995, 7, 36. [Google Scholar]
- Landolt, E.; Kandeler, R. The family of Lemnaceae—A monographic study, Phytochemistry, physiology, application and bibliography. In Biosystematic Investigations in the Family of Duckweeds (Lemnaceae); Geobotanischen Instutites der ETH, Stiftung Rubel: Zurich, Switzerland, 1987; Volume 4, p. 638. [Google Scholar] [CrossRef]
- Guimaraes, F.P.; Aguiar, R.; Oliveira, J.A.; Silva, J.A.A.; Karam, D. Potential of macrophyte for removing arsenic from aqueous solution. Planta Daninha 2012, 30, 112–118. [Google Scholar] [CrossRef]
- Stein, J. (Ed.) Handbook of Phycological Methods. Culture Methods and Growth Measurements; Cambridge University Press: Cambridge, UK, 1973; 448p. [Google Scholar]
- EC. Biological Test Method: Growth Inhibition Test Using a Freshwater Alga, EPS1/RM/25. 2007. Available online: https://www.canada.ca/en/environment-climate-change/services/wildlife-research-landscape-science/biological-test-method-publications/growth-inhibition-test-freshwater-alga.html (accessed on 10 April 2018).
- EC. Biological Test Method: Test for Measuring the Inhibition of Growth Using the Freshwater Macrophyte, Lemna minor, EPS1/RM/37. 2014. Available online: https://www.canada.ca/en/environment-climate-change/services/wildlife-research-landscape-science/biological-test-method-publications/inhibition-growth-freshwater-macrophyte/chapter-2.html (accessed on 24 April 2018).
- Silva, C.P.; Otero, M.; Esteves, V. Processes for the elimination of estrogenic steroid hormones from water: A review. Environ. Pollut. 2012, 165, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Mota, V.C.; Martins, C.I.M.; Eding, E.H.; Canário, A.V.M.; Verreth, J.A.J. Steroids accumulate in the rearing water of commercial recirculating aquaculture systems. Aquac. Eng. 2014, 62, 9–16. [Google Scholar] [CrossRef]
- Markovic, M.; Neale, P.A.; Nidumolu, B.; Kumar, R. Combined toxicity of therapeutic pharmaceuticals to duckweed, Lemna minor. Ecotoxicol. Environ.Saf. 2021, 208, 111428. [Google Scholar] [CrossRef]
- Santos, A.N.; Fachini, A.A.; Pena, A.; Delerue-Matos, C.; Montenegro, M.C.B.S.M. Ecotoxicological aspects related to the presence of pharmaceuticals in the aquatic Environment. J. Hazard. Mater. 2010, 175, 45–95. [Google Scholar] [CrossRef] [Green Version]
- Eaton, A.D.; Franson, M.A.H. American Public Health, American Water Works, and Water Environment: Standard Methods for the Examination of Water & Wastewater; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- International Standard ISO/FDIS/20079; Water Quality—Determination of Toxic Effect of Water Constituents and Wastewater to Duckweed (Lemna minor)-Duckweed Growth Inhibition Test. International Organization for Standardization: Geneva, Switzerland, 2005.
- Greenberg, B.M.; Huang, X.D.; Dixon, D.G. Applications of the aquatic higher plant Lemna gibba for Ecotoxicological Assessment. J. Aquat. Ecosyst. Health 1992, 1, 147–155. [Google Scholar] [CrossRef]
- Franklin, N.M.; Stauber, J.L.; Markich, S.J.; Lim, R.P. pH-dependent toxicity of copper and uranium to a tropical freshwater alga (Chlorella sp.). Aquat. Toxicol. 2000, 48, 275–289. [Google Scholar] [CrossRef]
- Wilde, K.L.; Stauber, J.L.; Markich, S.J.; Franklin, N.M.; Brown, P.L. The effect of pH on the uptake and toxicity of copper and zinc in a tropical freshwater alga (Chlorella sp.). Arch. Environ. Contam. Toxicol. 2006, 51, 174–185. [Google Scholar] [CrossRef]
- Nichols, D.J.; Daniel, T.C.; Edwards, D.R.; Moore, P.A., Jr.; Pote, D.H. Use of grass filter strips to reduce 17 beta-estradiol in runoff from fescue-applied poultry litter. J. Soil Water Conserv. 1998, 53, 74–77. [Google Scholar]
- Peterson, E.W.; Davis, R.K.; Orndorff, H.A. 17h-Estradiol as an indicator of animal waste contamination in mantled karst aquifers. J. Environ. Qual. 2001, 29, 826–834. [Google Scholar] [CrossRef]
- Ying, G.G.; Kookanaa, R.S.; Ru, Y.J. Occurrence and fate of hormone steroids in the environment. Environ. Int. 2002, 28, 545–551. [Google Scholar] [CrossRef]
- Patel, M.; Kumar, R.; Kishor, K.; Mlsna, T.; Pittman, C.U., Jr.; Mohan, D. Pharmaceuticals of Emerging Concern in Aquatic Systems: Chemistry, Occurrence, Effects, and Removal Methods. Chem. Rev. 2019, 119, 3510–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ECETOC (European Centre for Ecotoxicology and Toxicology of Chemicals). Environmental Risk Assessment of Difficult Substances; Technical Report No. 88; ECETOC: Brussels, Belgium, 2003; pp. 17–33. [Google Scholar]
- De Bree, L.C.J.; Janssen, R.; Aaby, P.; van Crevel, R.; Joosten, L.A.B.; Stabell Benn, C.; Netea, M.G. The impact of sex hormones on BCG-induced trained immunity. J. Leukoc. Biol. 2018, 104, 573–578. [Google Scholar] [CrossRef]
- Escribese, M.M.; Kraus, T.; Rhee, E.; Fernandez-Sesma, A.; López, C.B.; Moran, T.M. Estrogen inhibits dendritic cell maturation to RNA viruses. Blood 2008, 112, 4574–4584. [Google Scholar] [CrossRef] [Green Version]
- Saraceno, G.E.; Bellini, M.J.; Garcia-Segura, L.M.; Capani, F. Estradiol activates PI3K/Akt/GSK3 pathway under chronic neurodegenerative conditions triggered by perinatal asphyxia. Front. Pharmacol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Verma, M.; Pandey, S.; Bhat, I.A.; Mukesh, B.; Anand, J.; Chandra, V.; Sharma, G.T. Impact of l-carnitine on lipid content and post thaw survivability of buffalo embryos produced in vitro. Cryobiology 2018, 82, 99–105. [Google Scholar] [CrossRef]
- Fujiwara, K.; Matsumoto, Y.; Kawakami, H.; Aoki, M.; Tuzuki, M. Evaluation of metal toxicity in Chlorella kessleri from the perspective of the Periodic Table. Bull. Chem. Soc. Jpn. 2008, 81, 478–488. [Google Scholar] [CrossRef]
- Morris, J.M.; Lipton, J.; Brinkman, S. Copper toxicity to Rainbow Trout and Fathead Minnows in Low-Hardness Waters: Comparisons of BLM Predictions of Toxicity to Bioassay Results Using Laboratory Water and Site-Collected Water from Upper Talarik Creek in Bristol Bay, Southwest Alaska Salmon Science Workshop, Anchorage, AK, 2013, December 4–5. Available online: http://www.southwestsalmon.org/wpcontent/uploads/2014/01/Morris_Stratus_CuTox_RBT_FHM_LowHardnessH2O.pdf/ (accessed on 12 January 2021).
- Agami, M.; Reddy, R. Inter-relationships between Salvinia rotundifolia and Spirodela polyrhiza at various interaction stages. J. Aquat. Plant Manag. 1989, 27, 96–102. [Google Scholar]
- Davies, P.J. Plant Hormones; Kluwer Academic Publishers: Alphen aan den Rijn, The Netherlands, 2004; 716p. [Google Scholar] [CrossRef] [Green Version]
- Borker, A.R.; Mane, A.V.; Saratale, G.D.; Pathade, G.R. Phytoremediation potential of Eichhornia crassipes for the treatment of cadmium in relation with biochemical and water parameters. Emir. Emir. J. Food Agric. 2013, 25, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.B.; Hu, Q.; Sommerfeld, M.; Chen, F. Cell wall proteomics of the green alga Haematococcus pluvialis (Chlorophyceae). Proteomics 2004, 4, 692–708. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Du, W.; Liu, D. Perspectives of microbial oils for biodiesel production. Appl. Microbiol. Biotechnol. 2008, 80, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Damiani, M.C.; Popovich, C.A.; Constenla, D.; Leonardi, P.I. Lipid analysis in Haematococcus pluvialis to assess its potential use as a biodiesel feedstock. Bioresour. Technol. 2010, 101, 3801–3807. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, Y.; Schoefs, B. Secondary ketocarotenoid astaxanthin biosynthesis in algae: A multifunctional response to stress. Photosynth. Res. 2010, 106, 155–177. [Google Scholar] [CrossRef] [PubMed]
- Joss, A.; Andersen, H.; Ternes, T.; Richle, P.R.; Siegrist, H. Removal of estrogens in municipal wastewater treatment under aerobic and anaerobic conditions: Consequences for Plant Optimization. Environ. Sci. Technol. 2004, 38, 3047–3055. [Google Scholar] [CrossRef]
- Muradov, N.; Fidalgo, B.; Gujara, A.C.; T-Raissi, A. Pyrolysis of fast-growing aquatic biomass—Lemna minor (Duckweed): Characterization of pyrolysis products. Bioresour. Technol. 2014, 101, 8424–8428. [Google Scholar] [CrossRef]
- Landesman, L.; Parker, N.C.; Fedler, C.B.; Konikoff, M. Modeling duckweed growth in wastewater treatment systems. Livest. Res. Rural Dev. 2005, 17, 2005. [Google Scholar]
- Frédéric, M.; Samir, L.; Louise, M.; Abdelkrim, A. Comprehensive modeling of mat density effect on duckweed (Lemna minor) growth under controlled eutrophication. Water Res. 2006, 40, 2901–2910. [Google Scholar] [CrossRef]
- Khvatkov, P.; Chernobrovkina, M.; Okuneva, A.; Dolgov, S. Creation of culture media for efficient Duckweeds micropropagation (Wolffia arrhiza and Lemna minor) using artificial mathematical optimization models. Plant Cell Tissue Organ Cult. 2018, 139, 85–100. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. quadricauda | L. minor | |||
|---|---|---|---|---|
| E2 Concentration, (ng/L) | Cell Size (µm) | Biomass (g drw/L) | Cell Density (×104/mL) | Biomass (g drw/L) |
| FWW trial | ||||
| Control | 44.77 ± 3.26 | 0.40 ± 0.07 | 100.07 ± 5.23 | 3.40 ± 0.05 |
| 0.25 | 46.65 ± 4.73 a | 1.15 ± 0.11 *a | 241.43 ± 3.87 *a | 5.05 ± 0.09 *a |
| 0.75 | 46.87 ± 2.31 a | 1.79 ± 0.05 *b | 338.34 ± 3.18 *b | 4.94 ± 0.06 *a |
| 2.5 | 42.05 ± 1.45 a | 0.88 ± 0.10 *c | 199.61 ± 4.93 *c | 4.86 ± 0.08 *a |
| 5 | 37.23 ± 1.85 b | 0.86 ± 0.14 *c | 208.12 ± 8.89 *d | 4.89 ± 0.09 *a |
| 15 | 31.65 ± 2.22 +c | 0.78 ± 0.08 *c | 152.63 ± 5.47 *e | 5.59 ± 0.08 *b |
| RFWW trial | ||||
| Control | 43.77 ± 4.25 | 0.69 ± 0.09 | 242.67 ± 4.45 | 2.49 ± 0.07 |
| 0.25 | 47.14 ± 3.43 a | 0.44 ± 0.05 +a | 188.81 ± 3.22 +a | 5.45 ± 0.07 *a |
| 0.75 | 48.25 ± 2.62 a | 0.46 ± 0.04 +a | 191.52 ± 3.42 +a | 5.98 ± 0.08 *b |
| 2.5 | 39.84 ± 5.66 a | 0.41 ± 0.07 +a | 200.21 ± 5.25 +ab | 4.71 ± 0.09 *c |
| 5 | 35.92 ± 4.32 ab | 0.72 ± 0.11 b | 270.42 ± 8.61 *c | 4.86 ± 0.07 *c |
| 15 | 26.52 ± 3.65 +c | 0.76 ± 0.06 b | 312.71 ± 6.05 *d | 4.66 ± 0.04 *c |
| BBM trial | ||||
| Control | 44.76 ± 0.92 | 0.44 ± 0.11 | 128.51 ± 10.66 | 2.68 ± 0.07 |
| 0.25 | 52.67 ± 1.01 *a | 0.85 ± 0.09 *a | 202.12 ± 12.54 *a | 3.35 ± 0.08 *a |
| 0.75 | 56.72 ± 1.25 *b | 1.24 ± 0.04 *b | 254.24 ± 8.42 *b | 4.14 ± 0.06 *b |
| 2.5 | 46.74 ± 0.63 *c | 0.79 ± 0.08 *a | 179.13 ± 5.57 *c | 3.89 ± 0.09 *b |
| 5 | 46.84 ± 1.05 c | 0.81 ± 0.08 *a | 186.02 ± 14.05 *ac | 3.84 ± 0.06 *bc |
| 15 | 42.95 ± 1.48 d | 0.75± 0.06 *a | 164.64 ± 7.33 *ac | 2.95± 0.09 d |
| E2, ng/L | Control | 0.25 | 0.75 | 2.5 | 5 | 15 |
|---|---|---|---|---|---|---|
| Lemna minor | ||||||
| FWW | 36.12 ± 1.06 | 34.77 ± 0.76 a | 38.95 ± 0.48 *b | 40.41 ± 0.57 *b | 38.92 ± 0.91 *b | 39.68 ± 0.66 *b |
| RFWW | 31.73 ± 0.72 | 40.74 ± 1.21 *a | 41.12 ± 0.43 *a | 36.02 ± 0.79 *b | 32.82 ± 1.04 c | 32.51 ± 0.72 c |
| BBM | 24.75 ± 0.66 | 25.34 ± 0.26 a | 25.28 ± 1.01 a | 26.25 ± 0.76 a | 25.78 ± 0.86 a | 27.78 ± 0.58 *b |
| Scenedesmus quadricauda | ||||||
| FWW | 30.11 ± 0.56 | 37.22 ± 1.03 *a | 43.35 ± 0.19 *b | 46.76 ± 0.63 *c | 44.52 ± 1.52 *bc | 40.14 ± 1.06 *d |
| RFWW | 25.88 ± 0.69 | 29.94 ± 0.21 *a | 36.02 ± 0.48 *b | 39.93 ± 0.55 *c | 43.76 ± 1.04 *d | 29.57 ± 1.48 *a |
| BBM | 20.38 ± 0.21 | 21.84 ± 0.75 a | 31.08 ± 0.61 *b | 36.25 ± 0.65 *c | 38.78 ± 0.34 *d | 41.88 ± 0.71 *e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozlova, T.A.; Levin, D.B. Effect of 17β-Estradiol on Growth and Biosynthesis of Microalgae Scenedesmus quadricauda (CPCC-158) and Duckweed Lemna minor (CPCC-490) Grown in Three Different Media. Plants 2022, 11, 1669. https://doi.org/10.3390/plants11131669
Kozlova TA, Levin DB. Effect of 17β-Estradiol on Growth and Biosynthesis of Microalgae Scenedesmus quadricauda (CPCC-158) and Duckweed Lemna minor (CPCC-490) Grown in Three Different Media. Plants. 2022; 11(13):1669. https://doi.org/10.3390/plants11131669
Chicago/Turabian StyleKozlova, Tatiana A., and David B. Levin. 2022. "Effect of 17β-Estradiol on Growth and Biosynthesis of Microalgae Scenedesmus quadricauda (CPCC-158) and Duckweed Lemna minor (CPCC-490) Grown in Three Different Media" Plants 11, no. 13: 1669. https://doi.org/10.3390/plants11131669