
Variation in the Main Health-Promoting Compounds and Antioxidant Activity of Different Edible Parts of Purple Flowering Stalks (Brassica campestris var. purpuraria) and Green Flowering Stalks (Brassica campestris var. campestris)


Abstract
:1. Introduction
2. Results
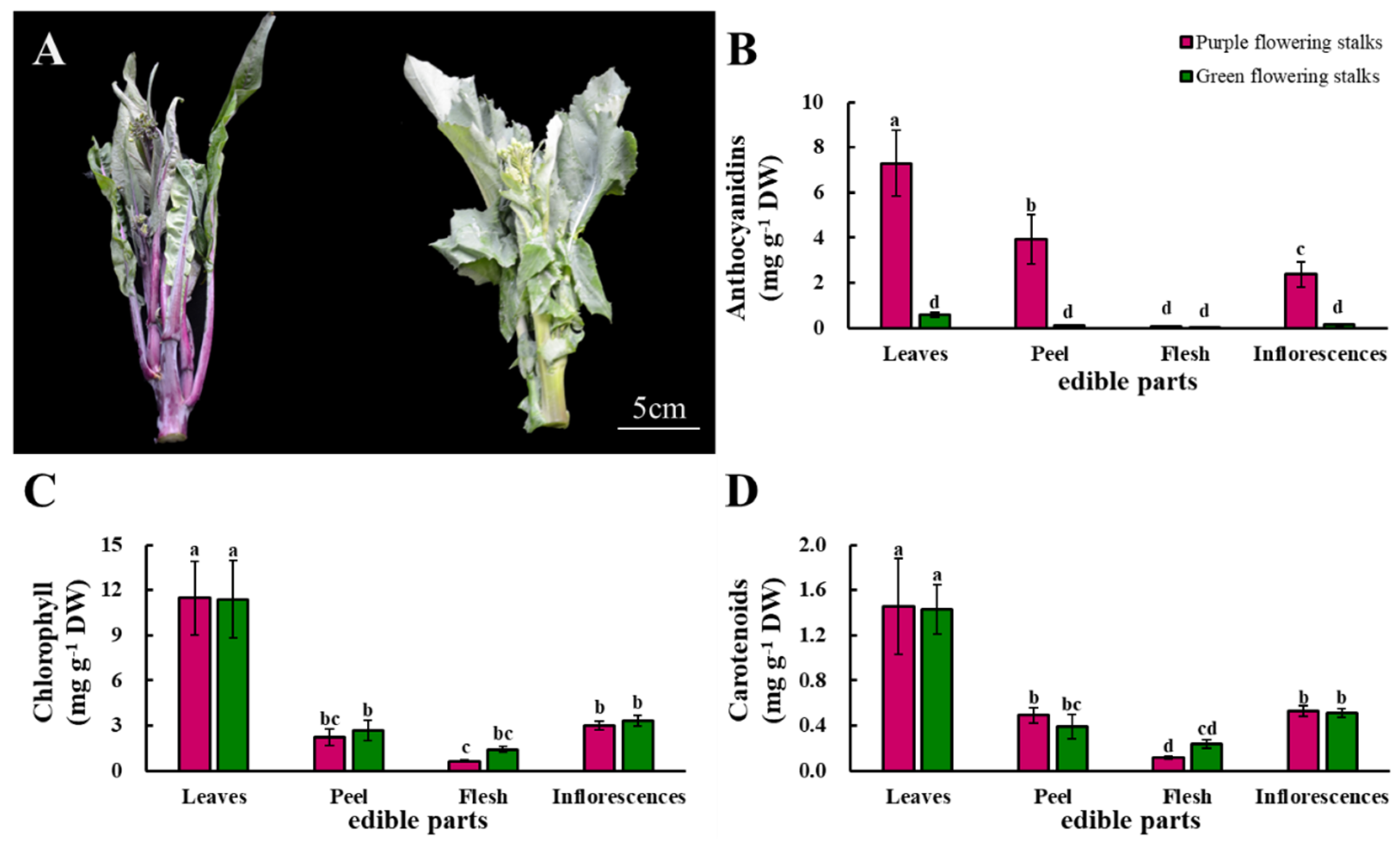
2.1. Anthocyanidins, Chlorophyll, and Carotenoids
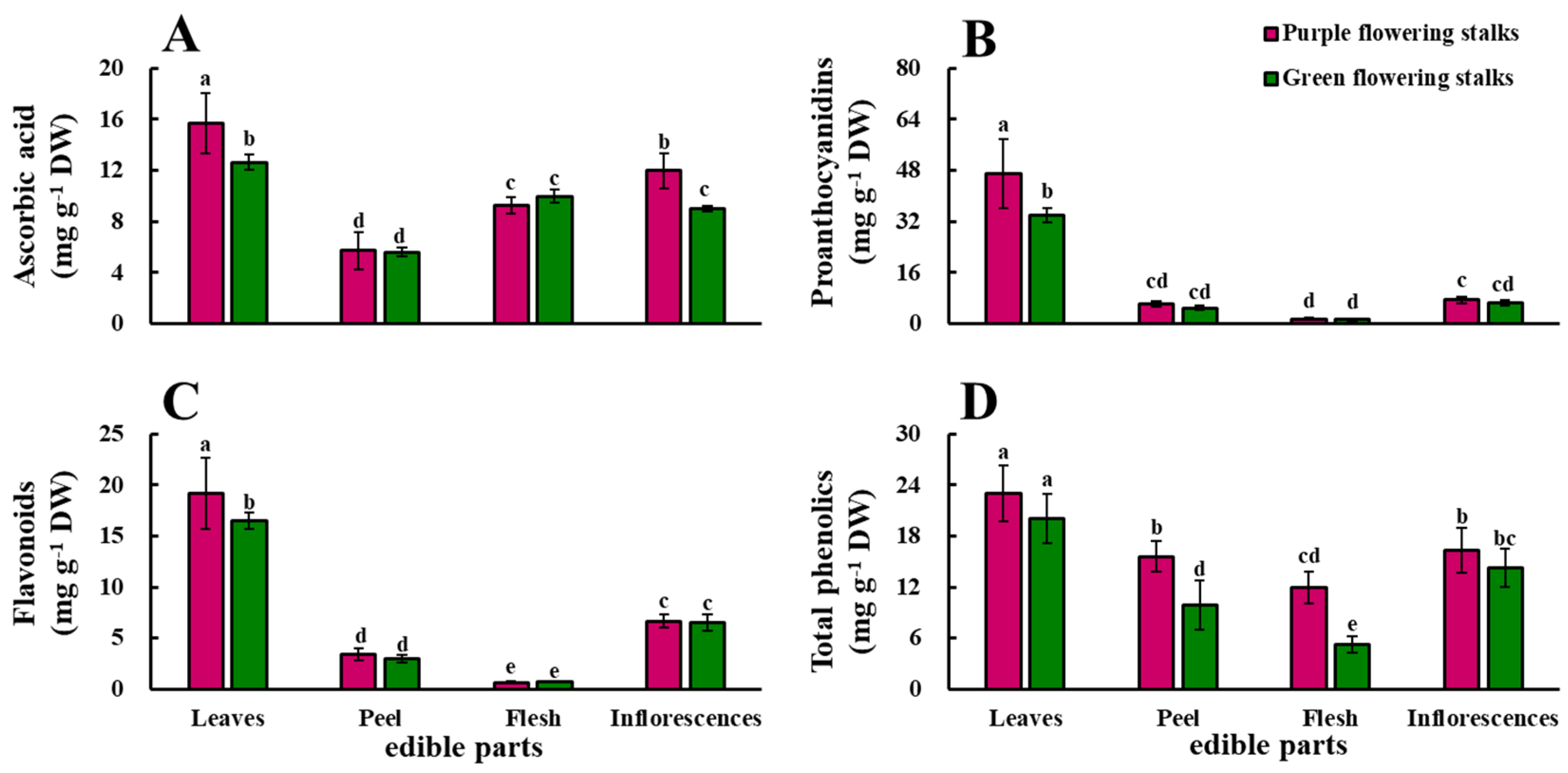
2.2. Ascorbic Acid
2.3. Proanthocyanidins, Flavonoids, and Total Phenolics
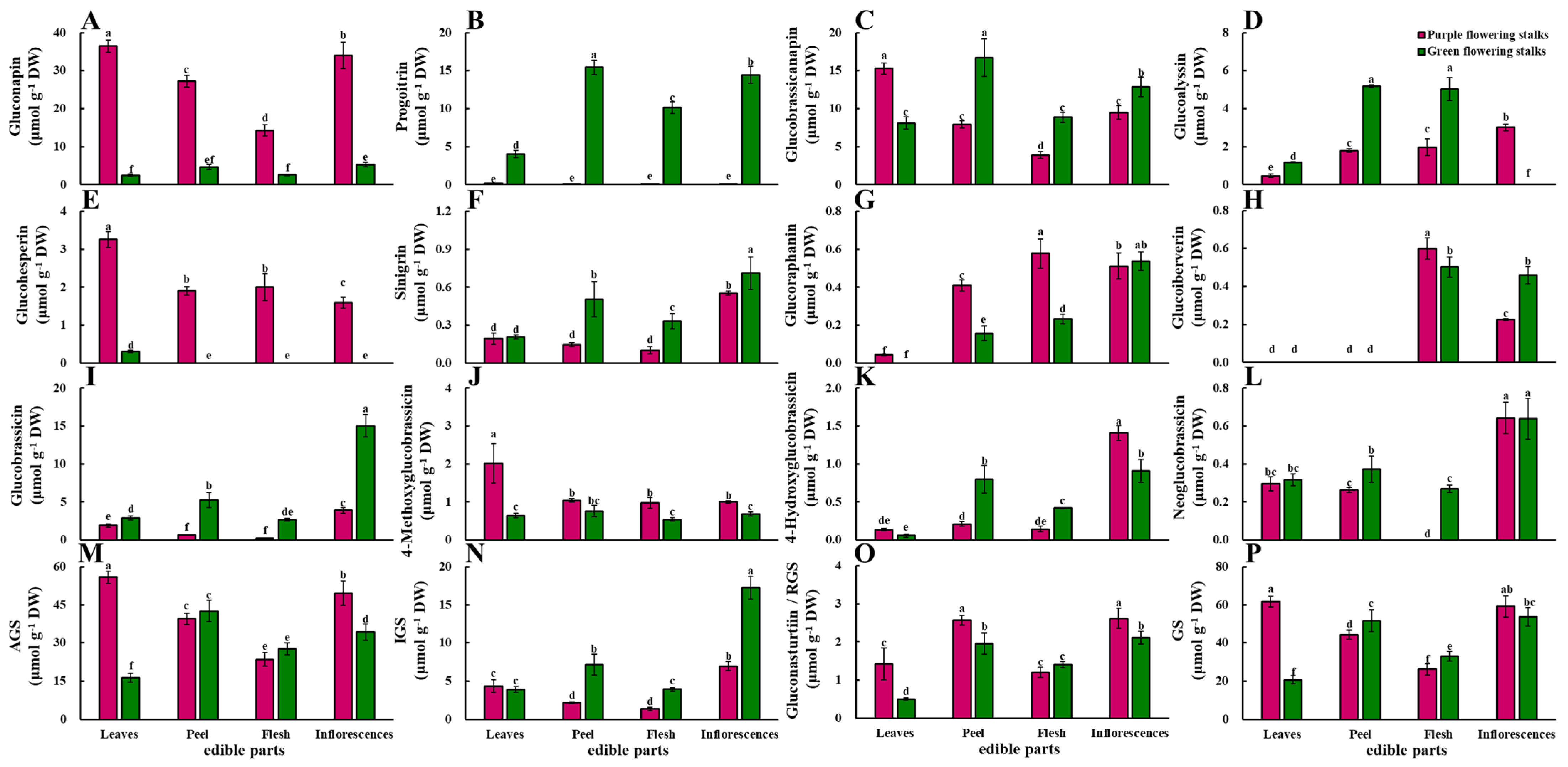
2.4. Glucosinolates
2.4.1. Aliphatic Glucosinolates
2.4.2. Indolic Glucosinolates
2.4.3. Aromatic Glucosinolate
2.4.4. Total Glucosinolate Content
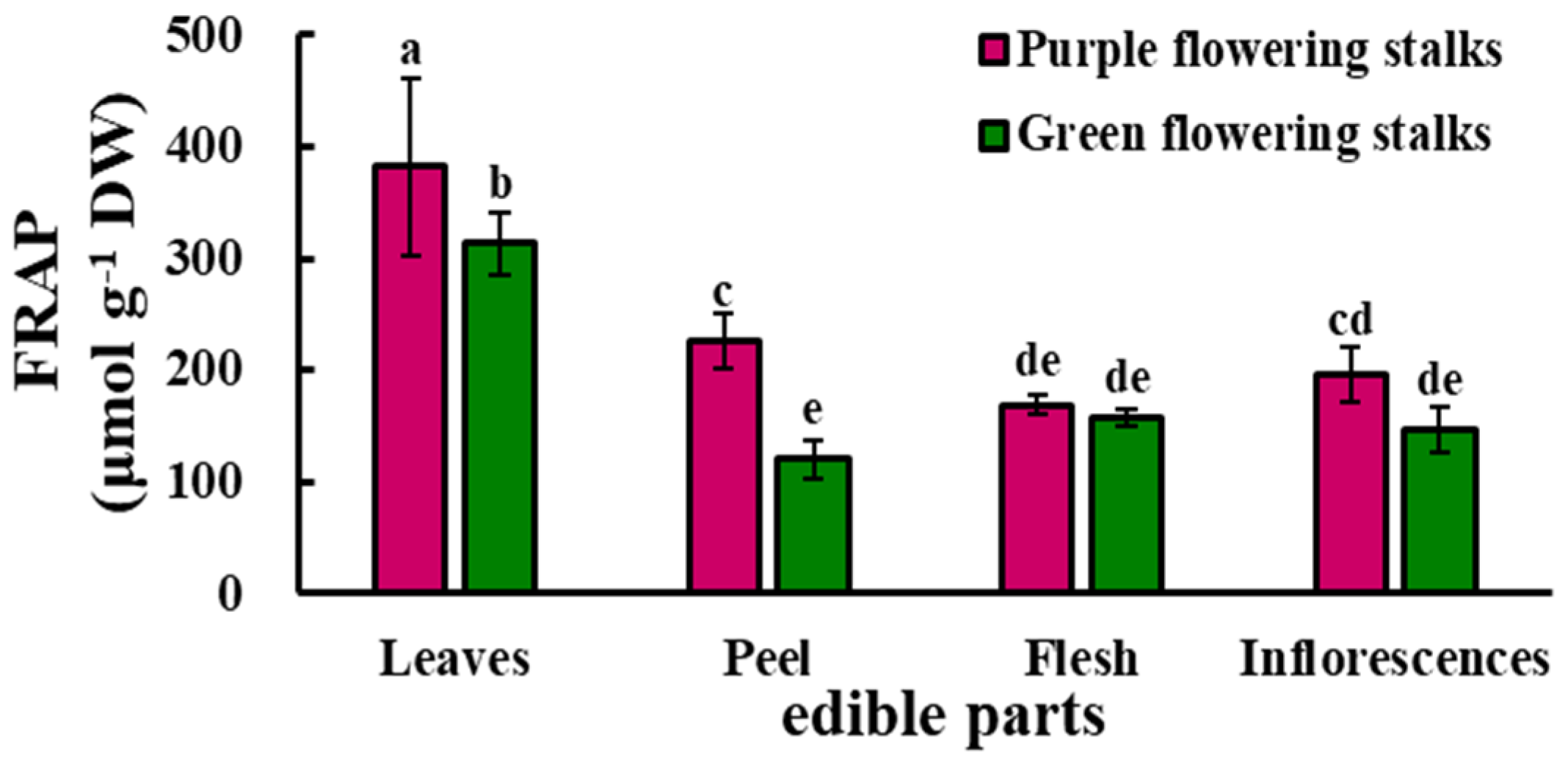
2.5. Antioxidant Capacity
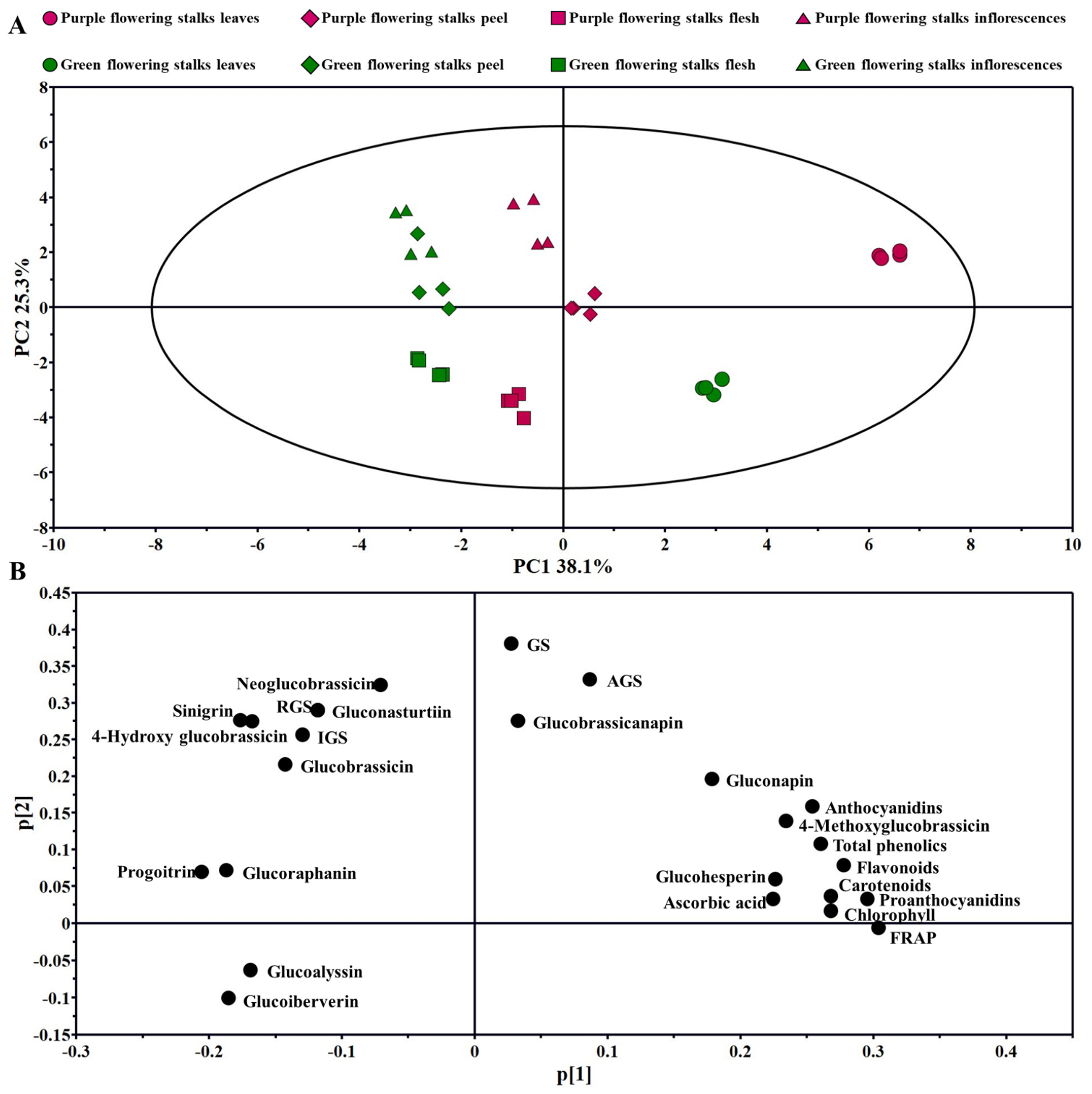
2.6. PCA
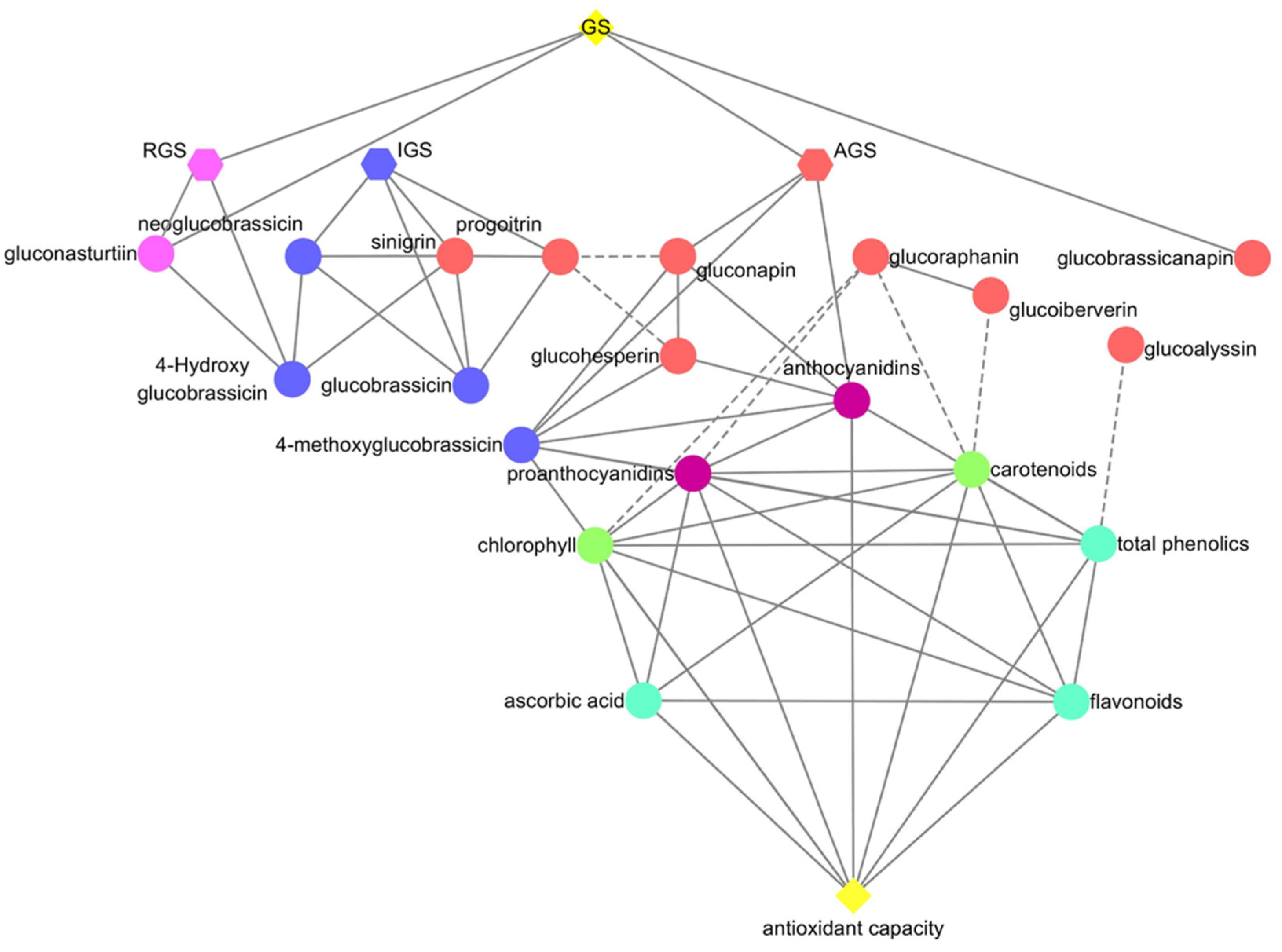
2.7. Correlation Analysis
2.8. Variance Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Anthocyanidin Content
4.3. Chlorophyll Content
4.4. Carotenoid Content
4.5. Ascorbic Acid Content
4.6. Proanthocyanidin Content
4.7. Flavonoid Content
4.8. Total Phenolic Content
4.9. Ferric Reducing Antioxidant Power (FRAP)
4.10. Glucosinolate Composition and Content
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghareaghajlou, N.; Hallaj-Nezhadi, S.; Ghasempour, Z. Red cabbage anthocyanins: Stability, extraction, biological activities and applications in food systems. Food Chem. 2021, 365, 130482. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.H.; Yuan, H.; Cao, H.B.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid Metabolism in Plants: The Role of Plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfer-Marquez, U.M.; Barros, R.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Reddy, C.V.K.; Sreeramulu, D.; Raghunath, M. Antioxidant activity of fresh and dry fruits commonly consumed in India. Food Res. Int. 2010, 43, 285–288. [Google Scholar] [CrossRef]
- Sancho, L.; Yahia, E.M.; Gonzalez-Aguilar, G.A. Identification and quantification of phenols, carotenoids, and vitamin C from papaya (Carica papaya L., cv. Maradol) fruit determined by HPLC-DAD-MS/MS-ESI. Food Res. Int. 2011, 44, 1284–1291. [Google Scholar] [CrossRef]
- Travica, N.; Ried, K.; Sali, A.; Scholey, A.; Hudson, I.; Pipingas, A. Vitamin C status and cognitive function: A systematic review. Nutrients 2017, 9, 960. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.Y.; Pan, H.B.; Jiang, H.; Wang, M.T.; Ye, X.Q.; Chen, S.G. Extraction and identification of proanthocyanidins from the leaves of persimmon and loquat. Food Chem. 2022, 372, 130780. [Google Scholar] [CrossRef]
- Leng, C.Q.; Zhang, Y.X.; Wang, M.; Wang, P.; Gu, Z.X.; Yang, R.Q. Dynamic variation of glucosinolates and isothiocyanates in broccoli sprouts during hydrolysis. Sci. Hortic. 2019, 255, 128–133. [Google Scholar] [CrossRef]
- Wang, J.S.; Yu, H.F.; Zhao, Z.Q.; Sheng, X.G.; Shen, Y.S.; Gu, H.H. Natural variation of glucosinolates and their breakdown products in Broccoli (Brassica oleracea var. italica) seeds. J. Agric. Food Chem. 2019, 67, 12528–12537. [Google Scholar] [CrossRef]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Chang, J.Q.; Wang, M.Y.; Jian, Y.; Zhang, F.; Zhu, J.; Wang, Q.M.; Sun, B. Health-promoting phytochemicals and antioxidant capacity in different organs from six varieties of Chinese kale. Sci. Rep. 2019, 9, 20344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.Z.; Wu, H.X.; Tollefsbol, T.O. Combined broccoli sprouts and green tea polyphenols contribute to the prevention of estrogen receptor-negative mammary cancer via cell cycle arrest and inducing apoptosis in HER2/neu Mice. J. Nutr. 2021, 151, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Juge, N.; Mithen, R.F.; Traka, M. Molecular basis for chemoprevention by sulforaphane: A comprehensive review. Cell. Mol. Life Sci. 2007, 64, 1105–1127. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.D.; Olsen, C.E.; Halkier, B.A. Production of the cancer-preventive glucoraphanin in tobacco. Mol. Plant 2010, 3, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Tian, Y.X.; Jiang, M.; Yuan, Q.; Chen, Q.; Zhang, Y.; Luo, Y.; Zhang, F.; Tang, H.R. Variation in the main health-promoting compounds and antioxidant activity of whole and individual edible parts of baby mustard (Brassica juncea var. gemmifera). RSC Adv. 2018, 8, 33845. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.; Tian, Y.X.; Chen, Q.; Zhang, Y.; Luo, Y.; Wang, Y.; Li, M.Y.; Gong, R.G.; Wang, X.R.; Zhang, F. Variations in the glucosinolates of the individual edible parts of three stem mustards (Brassica juncea). R. Soc. Open Sci. 2019, 6, 182054. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.E.; Shang, X.; Assefa, A.D.; Keum, Y.S.; Saini, R.K. Metabolite profiling of green, green/red, and red lettuce cultivars: Variation in health beneficial compounds and antioxidant potential. Food Res. Int. 2018, 105, 361–370. [Google Scholar] [CrossRef]
- Sharma, R.J.; Gupta, R.C.; Singh, S.; Bansal, A.K.; Singh, I.P. Stability of anthocyanins- and anthocyanidins-enriched extracts, and formulations of fruit pulp of Eugenia jambolana (‘jamun’). Food Chem. 2016, 190, 808–817. [Google Scholar] [CrossRef]
- Liu, S.C.; Lin, J.T.; Hu, C.C.; Shen, B.Y.; Chen, T.Y.; Chang, Y.L.; Shih, C.H.; Yang, D.J. Phenolic compositions and antioxidant attributes of leaves and stems from three inbred varieties of Lycium chinense Miller harvested at various times. Food Chem. 2017, 215, 284–291. [Google Scholar] [CrossRef]
- Li, H.; Xia, Y.; Liu, H.; Guo, H. Nutritional values, beneficial effects, and food applications of broccoli (Brassica oleracea var. italica Plenck). Trends Food Sci. Technol. 2022, 119, 288–308. [Google Scholar] [CrossRef]
- Mitharwal, S.; Kumar, A.; Chauhan, K.; Taneja, N.K. Nutritional, phytochemical composition and potential health benefits of taro (Colocasia esculenta L.) leaves: A review. Food Chem. 2022, 383, 132406. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Xu, T.; Tian, Y.; Zhong, Y.; Lu, B. Health benefits and phenolic compounds of Moringa oleifera leaves: A comprehensive review. Phytomedicine 2021, 93, 153771. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Ko, H.C.; Baek, H.J.; Cho, S.M.; Jang, H.H.; Lee, Y.M.; Kim, J.B. Identification and quantification of glucosinolates in Korean leaf mustard germplasm (Brassica juncea var. integrifolia) by liquid chromatography–electrospray ionization/tandem mass spectrometry. Eur. Food Res. Technol. 2016, 242, 1479–1484. [Google Scholar] [CrossRef] [Green Version]
- Giuffrida, F.; Agnello, M.; Mauro, R.P.; Ferrante, A.; Leonardi, C. Cultivation under salt stress conditions influences postharvest quality and glucosinolates content of fresh-cut cauliflower. Sci. Hortic. 2018, 236, 166–174. [Google Scholar] [CrossRef]
- Lee, M.K.; Chun, J.H.; Dong, H.B.; Chung, S.O.; Kim, S.J. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp. pekinensis) and their antioxidant activity. LWT Food Sci. Technol. 2014, 58, 93–101. [Google Scholar] [CrossRef]
- Sun, B.; Liu, N.; Zhao, Y.; Yan, H.; Wang, Q. Variation of glucosinolates in three edible parts of Chinese kale (Brassica alboglabra Bailey) varieties. Food Chem. 2011, 124, 941–947. [Google Scholar] [CrossRef]
- Helland, H.S.; Leufven, A.; Bengtsson, G.B.; Skaret, J.; Lea, P.; Wold, A. Storage of fresh-cut swede and turnip in modified atmosphere: Effects on vitamin C, sugars, glucosinolates and sensory attributes. Postharvest Biol. Technol. 2016, 111, 150–160. [Google Scholar] [CrossRef]
- Shi, H.; Zhao, Y.; Sun, J.; Yu, L.; Chen, P. Chemical profiling of glucosinolates in cruciferous vegetables-based dietary supplements using ultra-high performance liquid chromatography coupled to tandem high resolution mass spectrometry. J. Food Compos. Anal. 2017, 61, 67–72. [Google Scholar] [CrossRef]
- Koritsas, V.M.; Lewis, J.A.; Fenwick, G.R. Glucosinolate responses of oilseed rape, mustard and kale to mechanical wounding and infestation by cabbage stem flea beetle (Psylliodes chrysocephala). Ann. Appl. Biol. 1991, 118, 209–221. [Google Scholar] [CrossRef]
- Gu, H.; Wang, J.; Zhao, Z.; Sheng, X.; Yu, H.; Huang, W. Characterization of the appearance, health-promoting compounds, and antioxidant capacity of the florets of the loose-curd cauliflower. Int. J. Food Prop. 2015, 18, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Picchi, V.; Migliori, C.; Lo Scalzo, R.; Campanelli, G.; Ferrari, V.; Di Cesare, L.F. Phytochemical content in organic and conventionally grown Italian cauliflower. Food Chem. 2012, 130, 501–509. [Google Scholar] [CrossRef]
- Wang, J.S.; Gu, H.; Yu, H.; Zhao, Z.; Sheng, X.; Zhang, X. Genotypic variation of glucosinolates in broccoli (Brassica oleracea var. italica) florets from China. Food Chem. 2012, 133, 735–741. [Google Scholar] [CrossRef]
- Wang, J.S.; Zhao, Z.; Sheng, X.; Yu, H.; Gu, H. Influence of leaf-cover on visual quality and health-promoting phytochemicals in loose-curd cauliflower florets. LWT Food Sci. Technol. 2015, 61, 177–183. [Google Scholar] [CrossRef]
- Qian, H.; Sun, B.; Miao, H.; Cai, C.; Xu, C.; Wang, Q. Variation of glucosinolates and quinone reductase activity among different varieties of Chinese kale and improvement of glucoraphanin by metabolic engineering. Food Chem. 2015, 168, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Xu, K.; Zhu, Y.; Wang, F.; Xiao, J.; Guo, L. Calcium affects glucoraphanin metabolism in broccoli sprouts under ZnSO4 stress. Food Chem. 2021, 334, 127520. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J.; Lambrix, V.M.; Reichelt, M.; Gershenzon, J.; Mitchell-Olds, T. Gene duplication in the diversification of secondary metabolism: Tandem 2-oxoglutarate-dependent dioxygenases control glucosinolate biosynthesis in Arabidopsis. Plant Cell 2001, 13, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Augustine, R.; Bisht, N.C. Biofortification of oilseed Brassica juncea with the anti-cancer compound glucoraphanin by suppressing GSL-ALK gene family. Sci. Rep. 2015, 5, 18005. [Google Scholar] [CrossRef] [Green Version]
- Paśko, P.; Okoń, K.; Krośniak, M.; Prochownik, E.; Żmudzki, P.; Kryczyk-Kozioł, J.; Zagrodzki, P. Interaction between iodine and glucosinolates in rutabaga sprouts and selected biomarkers of thyroid function in male rats. J. Trace Elem. Med. Biol. 2018, 46, 110–116. [Google Scholar] [CrossRef]
- Sun, B.; Di, H.; Zhang, J.; Xia, P.; Huang, W.; Jian, Y.; Zhang, C.; Zhang, F. Effect of light on sensory quality, health-promoting phytochemicals and antioxidant capacity in post-harvest baby mustard. Food Chem. 2021, 339, 128057. [Google Scholar] [CrossRef]
- Deng, M.; Qian, H.; Chen, L.; Sun, B.; Chang, J.; Miao, H.; Cai, C.; Wang, Q. Influence of pre-harvest red light irradiation on main phytochemicals and antioxidant activity of Chinese kale sprouts. Food Chem. 2017, 222, 1–5. [Google Scholar] [CrossRef]
- KIM, S.J.; Ishii, G. Glucosinolate profiles in the seeds, leaves and roots of rocket salad (Eruca sativa Mill.) and anti-oxidative activities of intact plant powder and purified 4-methoxyglucobrassicin. Soil Sci. Plant Nutr. 2006, 52, 394–400. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S.; Rengasamy, K.R. Profiling of nutritionally important metabolites in green/red and green perilla (Perilla frutescens Britt.) cultivars: A comparative study. Ind. Crop. Prod. 2020, 151, 112441. [Google Scholar] [CrossRef]
- Medic, A.; Jakopic, J.; Hudina, M.; Solar, A.; Veberic, R. Identification and quantification of the major phenolic constituents in Juglans regia L. peeled kernels andpellicles, using HPLC–MS/MS. Food Chem. 2021, 352, 129404. [Google Scholar] [CrossRef] [PubMed]
- Prakash, O.; Baskaran, R.; Kudachikar, V.B. Characterization, quantification of free, esterified and bound phenolics in Kainth (Pyrus pashia Buch.-Ham. Ex D.Don) fruit pulp by UPLC-ESI-HRMS/ MS and evaluation of their antioxidant activity. Food Chem. 2019, 299, 125114. [Google Scholar] [CrossRef]
- Ming, H.N.; Wang, Q.; Wu, Y.; Liu, H.M.; Zheng, L.M.; Zhang, G.F. Transcriptome analysis reveals the mechanism of anthocyanidins biosynthesis during grains development in purple corn (Zea mays L.). J. Plant Physiol. 2021, 257, 153328. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | VV/VP | VE/VP | VVE/VP |
|---|---|---|---|
| Anthocyanidins | 0.402 ** | 0.313 ** | 0.232 ** |
| Chlorophyll | 0.002 | 0.925 ** | 0.001 |
| Carotenoids | 0.000 | 0.902 ** | 0.006 |
| Ascorbic acid | 0.042 ** | 0.806 ** | 0.062 ** |
| Proanthocyanidins | 0.013 * | 0.917 ** | 0.026 * |
| Flavonoids | 0.004 | 0.959 ** | 0.007 |
| Total phenolics | 0.148 ** | 0.685 ** | 0.029 |
| Gluconapin | 0.787 ** | 0.111 ** | 0.092 ** |
| Progoitrin | 0.740 ** | 0.126 ** | 0.127 ** |
| Glucobrassicanapin | 0.091 ** | 0.328 ** | 0.521 ** |
| Glucoalyssin | 0.079 ** | 0.420 ** | 0.483 ** |
| Glucohesperin | 0.830 ** | 0.108 ** | 0.048 ** |
| Sinigrin | 0.190 ** | 0.642 ** | 0.083 ** |
| Glucoraphanin | 0.123 ** | 0.728 ** | 0.119 ** |
| Glucoiberverin | 0.005 ** | 0.921 ** | 0.062 ** |
| Glucobrassicin | 0.289 ** | 0.504 ** | 0.191 ** |
| 4-Methoxyglucobrassicin | 0.413 ** | 0.221 ** | 0.228 ** |
| 4-Hydroxy glucobrassicin | 0.006 * | 0.768 ** | 0.196 ** |
| Neoglucobrassicin | 0.060 ** | 0.811 ** | 0.070 ** |
| Total aliphatic glucosinolates | 0.218 ** | 0.258 ** | 0.480 ** |
| Total indolic glucosinolates | 0.210 ** | 0.595 ** | 0.173 ** |
| Gluconasturtiin | 0.106 ** | 0.735 ** | 0.084 ** |
| Total aromatic glucosinolate | 0.106 ** | 0.735 ** | 0.084 ** |
| Total glucosinolates | 0.073 ** | 0.437 ** | 0.438 ** |
| Antioxidant capacity | 0.107 ** | 0.750 ** | 0.036 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Di, H.; Cheng, W.; Ren, G.; Luo, S.; Ma, J.; Ma, W.; Lian, H.; Li, X.; Huang, Z.; et al. Variation in the Main Health-Promoting Compounds and Antioxidant Activity of Different Edible Parts of Purple Flowering Stalks (Brassica campestris var. purpuraria) and Green Flowering Stalks (Brassica campestris var. campestris). Plants 2022, 11, 1664. https://doi.org/10.3390/plants11131664
Wang Y, Di H, Cheng W, Ren G, Luo S, Ma J, Ma W, Lian H, Li X, Huang Z, et al. Variation in the Main Health-Promoting Compounds and Antioxidant Activity of Different Edible Parts of Purple Flowering Stalks (Brassica campestris var. purpuraria) and Green Flowering Stalks (Brassica campestris var. campestris). Plants. 2022; 11(13):1664. https://doi.org/10.3390/plants11131664
Chicago/Turabian StyleWang, Yating, Hongmei Di, Wenjuan Cheng, Guanru Ren, Sha Luo, Jie Ma, Wei Ma, Huashan Lian, Xiaomei Li, Zhi Huang, and et al. 2022. "Variation in the Main Health-Promoting Compounds and Antioxidant Activity of Different Edible Parts of Purple Flowering Stalks (Brassica campestris var. purpuraria) and Green Flowering Stalks (Brassica campestris var. campestris)" Plants 11, no. 13: 1664. https://doi.org/10.3390/plants11131664