
Is Tempranillo Blanco Grapevine Different from Tempranillo Tinto Only in the Color of the Grapes? An Updated Review

 , ,
, , 
Abstract
:1. An Introduction to Tempranillo Blanco, a New Variety That Originated from Tempranillo Tinto
2. The Fruit-Bearing Cutting Technique as a Tool to Investigate Differences between Tempranillo Blanco and Tinto
3. The Temperature Gradient Greenhouses Simulate Future Climate Conditions for Growing Tempranillo Blanco and Tinto
4. Growth, Water Use, and Production of Tempranillo Blanco and Tinto Fruit-Bearing Cuttings under Simulated Climate Change Conditions
5. Grape Quality of Tempranillo Blanco and Tinto Fruit-Bearing Cuttings under Simulated Climate Change Conditions
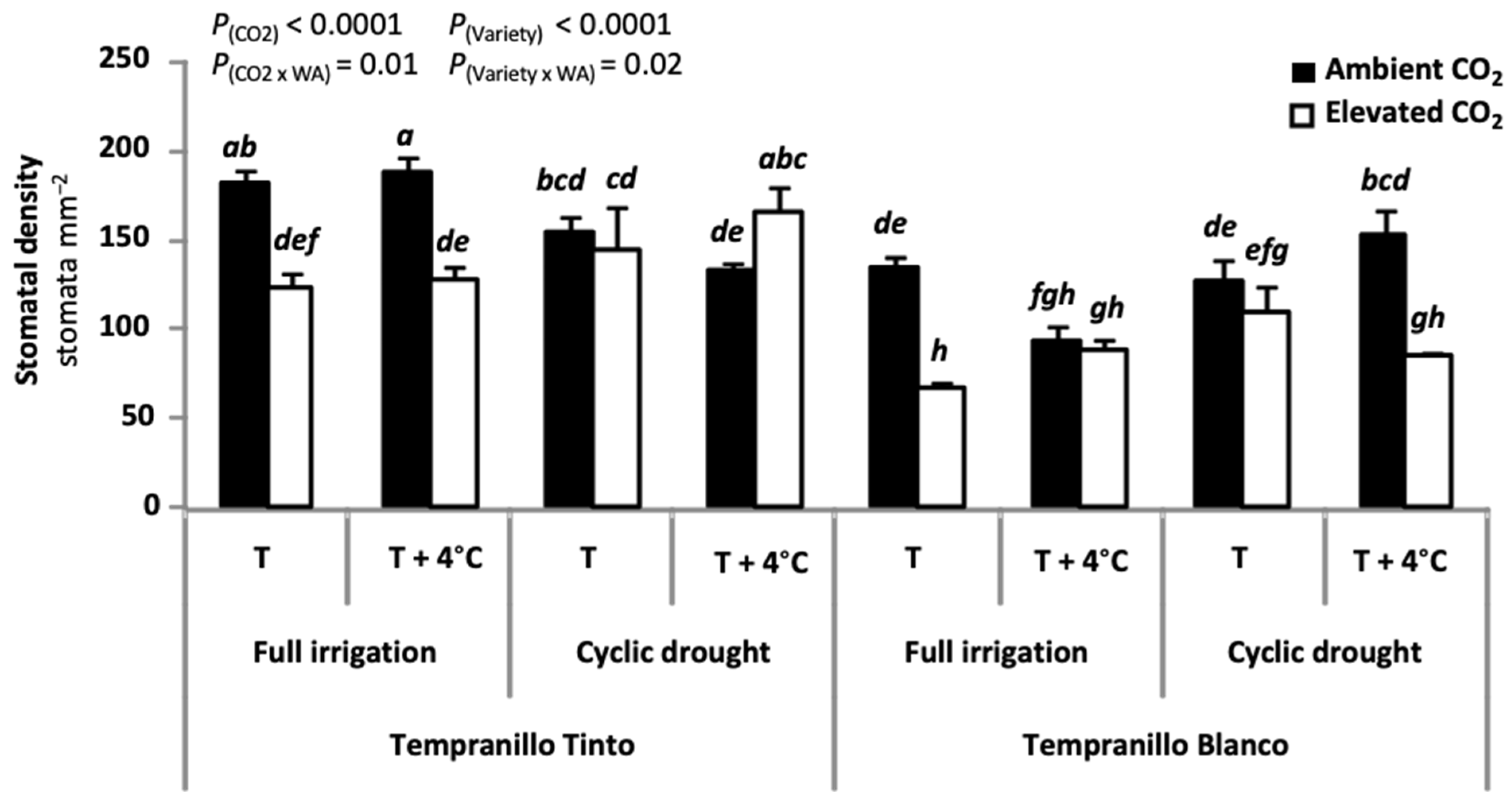
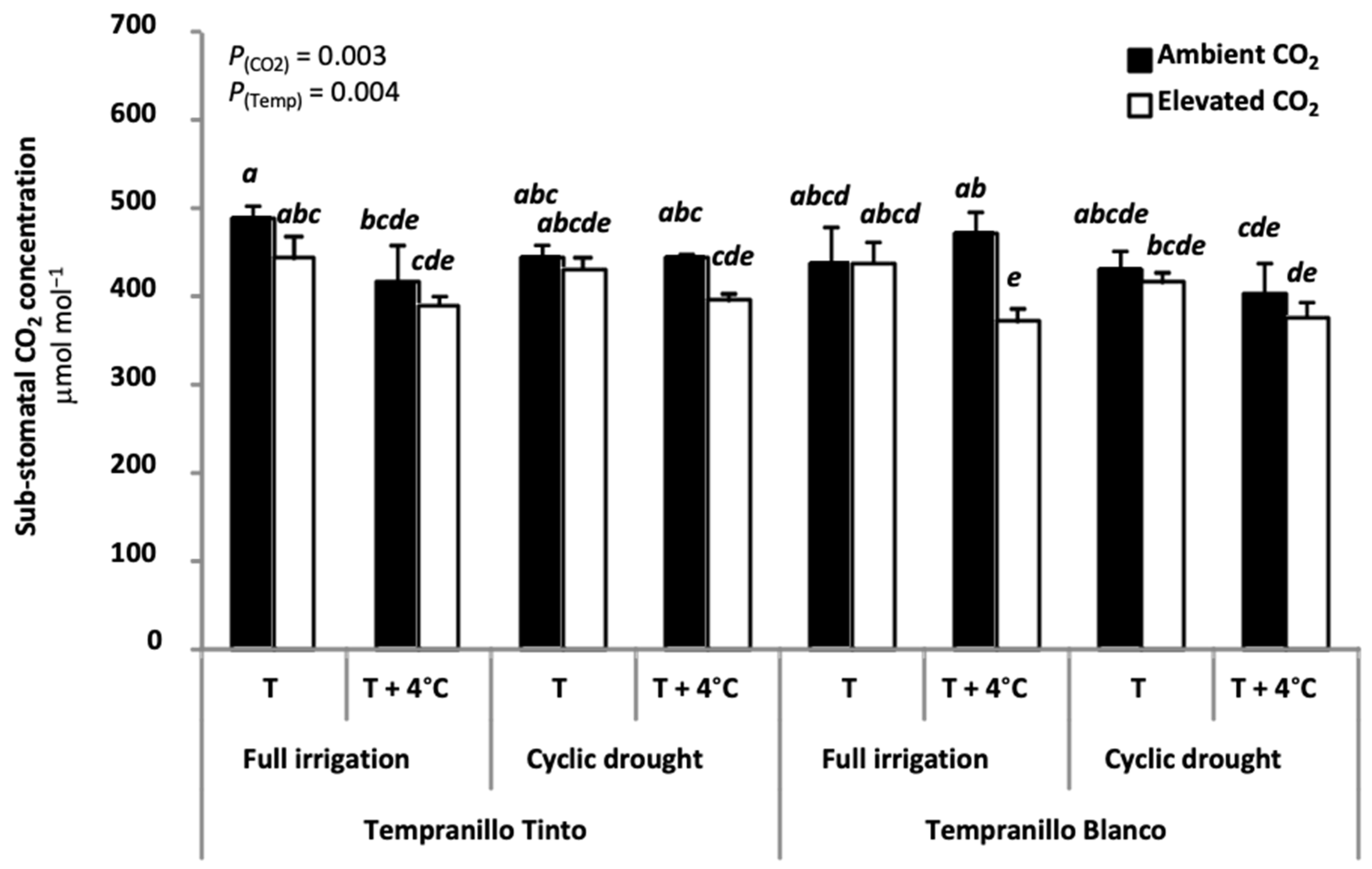
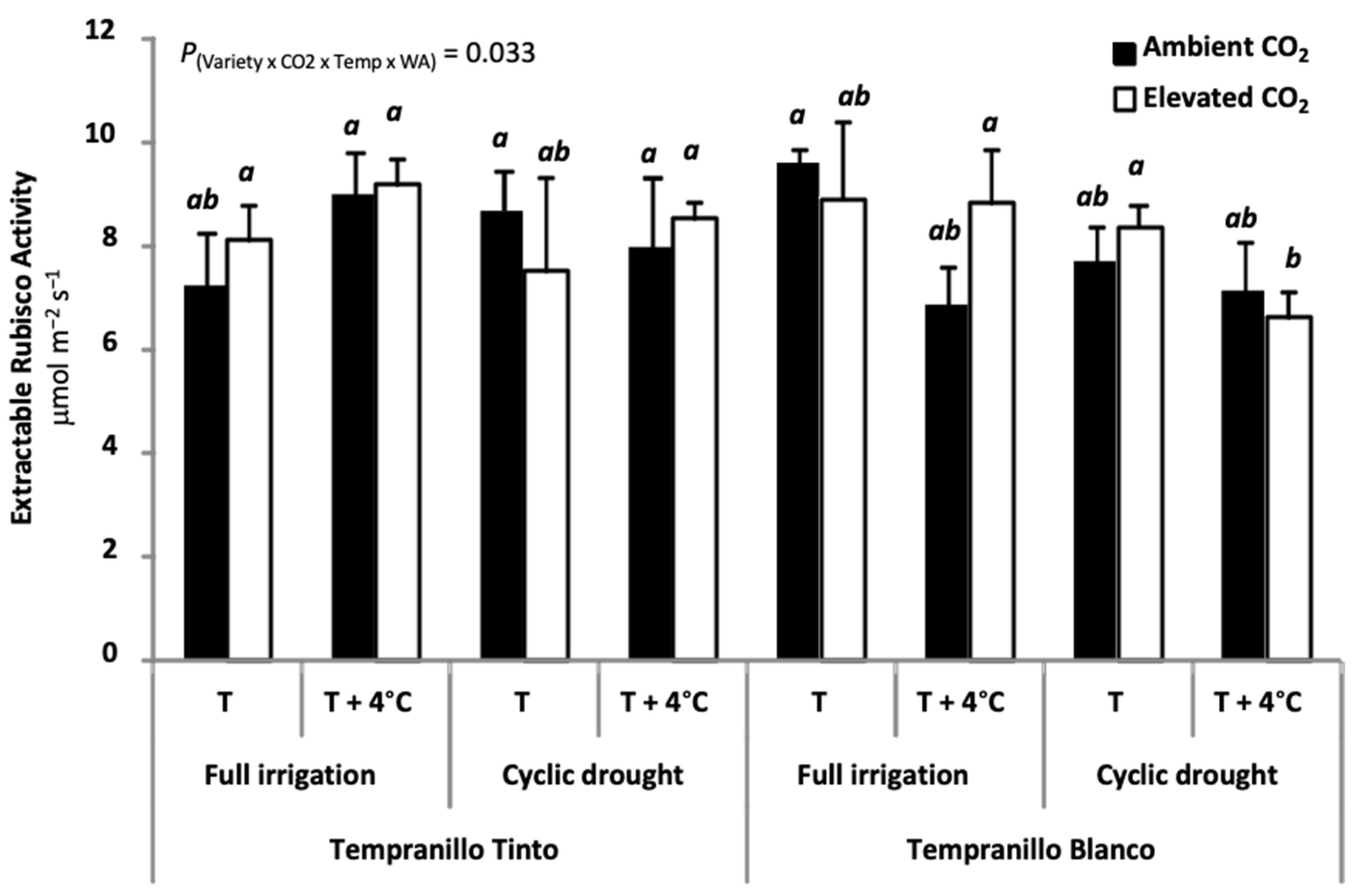
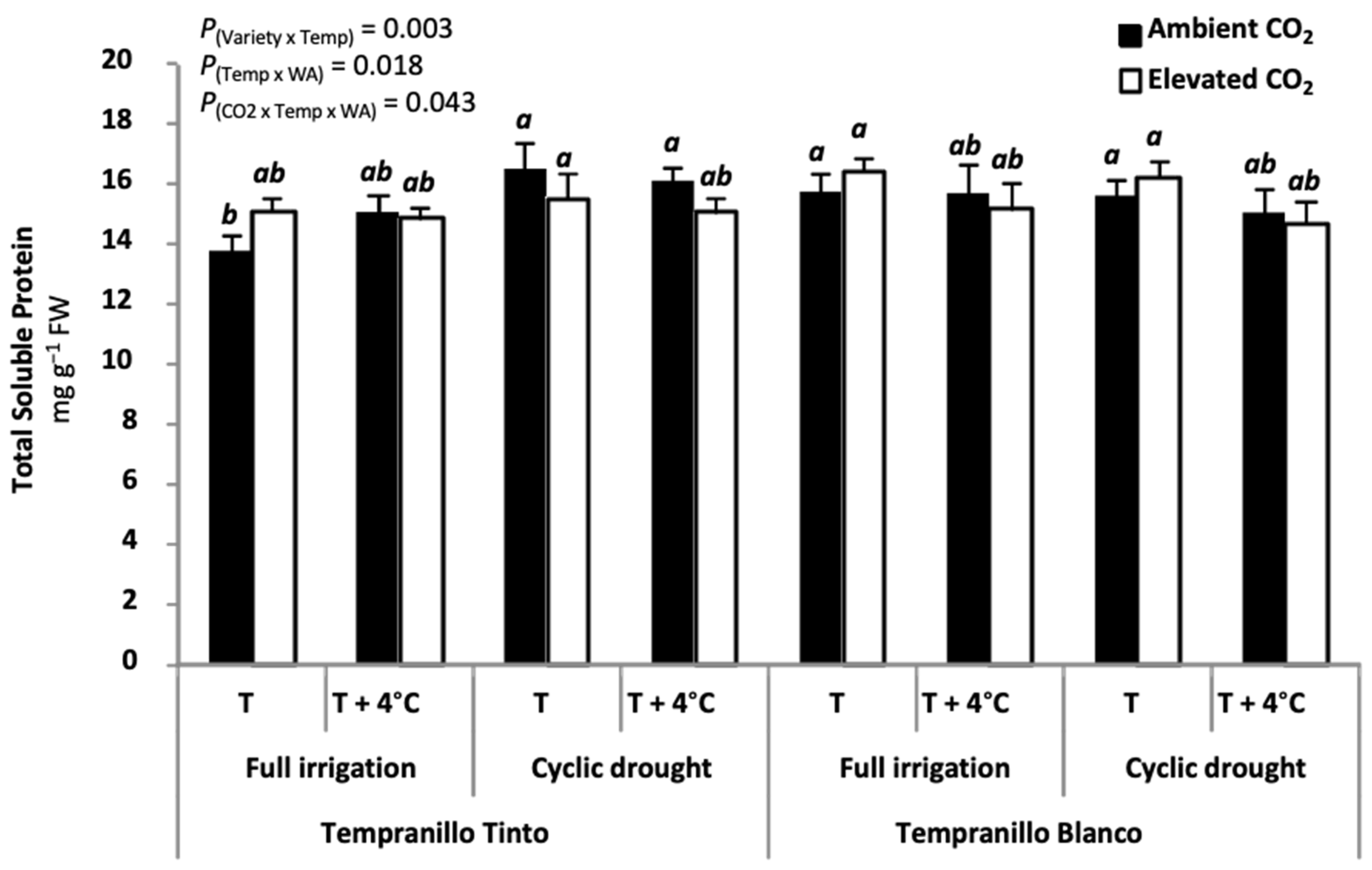
6. Physiology of Tempranillo Blanco and Tinto: Gas Exchange Properties and Photosynthetic Acclimation
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez, J.; Vicente, T.; Martínez, T.; Chavarri, J.B.; García-Escudero, E. Una Nueva Variedad Blanca para la DOCa. Rioja: El Tempranillo Blanco. In Proceedings of the XXIX Congreso Mundial de la Viña y el Vino (OIV), Logroño, Spain, 25–30 June 2006. [Google Scholar]
- Carbonell-Bejerano, P.; Royo, C.; Torres-Pérez, R.; Grimplet, J.; Fernández, L.; Franco-Zorrilla, J.M.; Lijavetzky, D.; Baroja, E.; Martínez, J.; García-Escudero, E.; et al. Catastrophic unbalanced genome rearrangements cause somatic loss of berry color in grapevine. Plant Physiol. 2017, 175, 786–801. [Google Scholar] [CrossRef] [Green Version]
- Tello, J.; Royo, C.; Baroja, E.; García-Escudero, E.; Martínez-Zapater, J.M.; Carbonell-Bejerano, P. Reduced gamete viability associated to somatic genome rearrangements increases fruit set sensitivity to the environment in Tempranillo Blanco grapevine cultivar. Sci. Hortic. 2021, 290, 110497. [Google Scholar] [CrossRef]
- Balda, P.; Martínez de Toda, F. Variedades Minoritarias de vid en La Rioja. Consejería de Agricultura. 2017. Available online: https://www.larioja.org/agricultura/es/publicaciones-agricultura/monografias/variedades-minoritarias-vid-rioja (accessed on 6 June 2022).
- Zinelabidine, L.H.; Torres-Pérez, R.; Grimplet, J.; Baroja, E.; Ibañéz, S.; Carbonell-Bejerano, P.; Martínez-Zapater, J.M.; Ibáñez, J.; Tello, J. Genetic variation and association analyses identify genes linked to fruit set-related traits in grapevine. Plant Sci. 2021, 306, 110875. [Google Scholar] [CrossRef]
- Martínez, J.; López, E.; Baroja, E.; Pérez, D.; Chávarri, J.D.; García-Escudero, E. Evaluación Agronómica y Enológica de la Variedad Tempranillo Blanco (Vitis vinifera L.) y de Otras Variedades Minoritarias Blancas de la DO Ca. Rioja. In Proceedings of the Jornada del Grupo de Viticultura y Enología, Logroño, Spain, 19–20 November 2014. [Google Scholar]
- Mullins, M.G. Test-plant for investigations of the physiology of fruiting in Vitis vinifera L. Nature 1966, 209, 419–420. [Google Scholar] [CrossRef]
- Morales, F.; Antolín, M.C.; Aranjuelo, I.; Goicoechea, N.; Pascual, I. From vineyards to controlled environments in grapevine research: Investigating responses to climate change scenarios using fruit-bearing cuttings. Theor. Exp. Plant Physiol. 2016, 28, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Ollat, N.; Geny, L.; Soyer, J. Les boutures fructifères de vigne: Validation d’un modele d’étude du développement de la physiologie de la vigne. I. Caractéristiques de l’appareil végétatif. J. Int. Sci. Vigne Vin 1998, 32, 1–9. [Google Scholar]
- Morales, F.; Pascual, I.; Sánchez-Díaz, M.; Aguirreolea, J.; Irigoyen, J.J.; Goicoechea, N.; Antolín, M.C.; Oyarzun, M.; Urdiain, A. Methodological advances: Using greenhouses to simulate climate change scenarios. Plant Sci. 2014, 226, 30–40. [Google Scholar] [CrossRef]
- Rawson, H.M.; Gifford, R.M.; Condon, B.N. Temperature-gradient chambers for research on global environment change. 1. Portable chambers for research on short-stature vegetation. Plant Cell Environ. 1995, 18, 1048–1054. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Irigoyen, J.J.; Pascual, I.; Morales, F. Simulating the impact of climate change (elevated CO2 and temperature, and water deficit) on the growth of red and white Tempranillo grapevine in three consecutive growing seasons (2013–2015). Agric. Water Manag. 2018, 202, 220–230. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Pascual, I.; Irigoyen, J.J.; Morales, F. Future CO2, warming and water deficit impact white and red Tempranillo grapevine: Photosynthetic acclimation to elevated CO2 and biomass allocation. Physiol. Plantarum 2021, 172, 1779–1794. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Mekni, I.; Santesteban, H.; Pascual, I.; Morales, F.; Irigoyen, J.J. Effects of climate change including elevated CO2 concentration, temperature and water deficit on growth, water status, and yield quality of grapevine (Vitis vinifera L.) cultivars. Agric. Water Manag. 2015, 159, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Parra, C. Vid y Cambio Climático. Estudio del Proceso de Maduración de la Baya en Esquejes Fructíferos de Tempranillo en Respuesta a la Interacción de CO2 Elevado, Estrés Hídrico y Temperatura Elevada. Ph.D. Thesis, University of Navarra, Pamplona, Spain, 2011. [Google Scholar]
- Erice, G.; Pérez-Bueno, M.L.; Pineda, M.; Barón, M.; Aroca, R.; Calvo-Polanco, M. Determining plant water relations. In Advances in Plant Ecophysiology Techniques; Sánchez-Moreiras, A., Reigosa, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar] [CrossRef]
- Mullins, M.G.; Bouquet, A.; Williams, L.E. Biology of the Grapevine; Cambridge University Press: New York, NY, USA, 1992. [Google Scholar]
- Williams, L.E.; Matthews, M.A. Grapevine. In Irrigation of Agricultural Crops, Agronomy Monograph No. 30; Stewart, B.A., Nielsen, D.R., Eds.; ASA-CSSA-SSSA: Madison, WI, USA, 1990; pp. 1019–1055. [Google Scholar]
- Chaves, M.M.; Pereira, J.S. Water stress, CO2 and climate change. J. Exp. Bot. 1992, 43, 1131–1139. [Google Scholar] [CrossRef]
- Houel, C.; Martin-Magniette, M.L.; Nicolas, S.D.; Lacombe, T.; Le Cunff, L.; Franck, D.; Torregrosa, L.; Conéjéro, G.; Lalet, S.; This, P.; et al. Genetic variability of berry size in the grapevine (Vitis vinifera L.). Aust. J. Grape Wine Res. 2013, 19, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Bindi, M.; Fibbi, L.; Miglietta, F. Free Air CO2 Enrichment (FACE) of grapevine (Vitis vinifera L.): II. Growth and quality of grape and wine in response to elevated CO2 concentrations. Eur. J. Agron. 2001, 14, 145–155. [Google Scholar] [CrossRef]
- Bindi, M.; Fibbi, L.; Gozzini, B.; Orlandini, S.; Miglietta, F. Modelling the impact of future climate scenarios on yield and yield variability of grapevine. Clim. Res. 1996, 7, 213–224. [Google Scholar] [CrossRef]
- Schultz, H.R. Climate change and viticulture: A European perspective on climatology, carbon dioxide and UV-B effects. Aust. J. Grape Wine Res. 2000, 6, 2–12. [Google Scholar] [CrossRef]
- Salazar-Parra, C.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Morales, F. Photosynthetic response of Tempranillo grapevine to climate change scenarios. Ann. Appl. Biol. 2012, 161, 277–292. [Google Scholar] [CrossRef]
- Salazar-Parra, C.; Aranjuelo, I.; Pascual, I.; Erice, G.; Sanz-Sáez, A.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Araus, J.L.; Morales, F. Carbon balance, partitioning and photosynthetic acclimation in fruit-bearing grapevine (Vitis vinifera L. cv. Tempranillo) grown under simulated climate change (elevated CO2, elevated temperature and moderate drought) scenarios in temperature gradient greenhouses. J. Plant Physiol. 2015, 174, 97–109. [Google Scholar] [PubMed] [Green Version]
- Da Silva, J.R.; Patterson, A.E.; Rodrigues, W.P.; Campostrini, E.; Griffin, K.L. Photosynthetic acclimation to elevated CO2 combined with partial rootzone drying results in improved water use efficiency, drought tolerance and leaf carbon balance of grapevines (Vitis labrusca). Environ. Exp. Bot. 2017, 134, 82–95. [Google Scholar] [CrossRef]
- Leakey, A.D.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef]
- Flexas, J.; Carriquí, M.; Coopman, R.E.; Gago, J.; Galmés, J.; Martorell, S.; Morales, F.; Díaz-Espejo, A. Stomatal and mesophyll conductances to CO2 in different plant groups: Underrated factors for predicting leaf photosynthesis responses to climate change? Plant Sci. 2014, 226, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Moutinho-Pereira, J.M.; Gonçalves, B.; Bacelar, E.; Boaventura Cunha, J.; Coutinho, J.; Correia, C.M. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Physiological and yield attributes. Vitis 2009, 48, 159–165. [Google Scholar]
- Rogiers, S.Y.; Hardie, W.J.; Smith, J.P. Stomatal density of grapevine leaves (Vitis vinifera L.) responds to soil temperature and atmospheric carbon dioxide. Aust. J. Grape Wine Res. 2011, 17, 147–152. [Google Scholar] [CrossRef]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Calfapietra, C.; McCarthy, H.R.; Norby, R.J.; Francesca Cotrufo, M. Elevated CO2 increases tree-level intrinsic water use efficiency: Insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites. New Phytol. 2013, 197, 544–554. [Google Scholar] [CrossRef] [Green Version]
- Kizildeniz, T.; Pascual, I.; Irigoyen, J.J.; Morales, F. Using fruit-bearing cuttings of grapevine and temperature gradient greenhouses to evaluate effects of climate change (elevated CO2 and temperature, and water deficit) on the cv. red and white Tempranillo. Yield and must quality in three consecutive growing seasons (2013–2015). Agric. Water Manag. 2018, 202, 299–310. [Google Scholar]
- Poni, S.; Gatti, M.; Palliotti, A.; Dai, Z.W.; Duchêne, E.; Truong, T.-T.; Ferrara, G.; Matarrese, A.M.S.; Gallotta, A.; Bellincontro, A.; et al. Grapevine quality: A multiple choice issue. Sci. Hortic. 2018, 234, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vučetić, V.; Dai, Z.; Luedeling, E.; Van Leeuwen, C.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of grapevine phenology to water availability, temperature and CO2 concentration. Front. Environ. Sci. 2016, 4, 48. [Google Scholar] [CrossRef]
- Arrizabalaga-Arriazu, M.; Gomès, E.; Morales, F.; Irigoyen, J.J.; Pascual, I.; Hilbert, G. High temperature and elevated carbon dioxide modify berry composition of different clones of grapevine (Vitis vinifera L.) cv. Tempranillo. Front. Plant Sci. 2020, 11, 603687. [Google Scholar] [CrossRef]
- Arrizabalaga-Arriazu, M.; Gomès, E.; Morales, F.; Irigoyen, J.J.; Pascual, I.; Hilbert, G. Impact of 2100-projected air temperature, carbon dioxide, and water scarcity on grape primary and secondary metabolites of different Vitis vinifera cv. Tempranillo clones. J. Agric. Food. Chem. 2021, 69, 6172–6185. [Google Scholar] [CrossRef]
- Acevedo De la Cruz, A.; Hilbert, G.; Rivière, C.; Mengin, V.; Ollat, N.; Bordenave, L.; Decroocq, S.; Delaunay, J.-C.; Delrot, S.; Merillon, J.-M.; et al. Anthocyanin identification and composition of wild Vitis spp. accessions by using LC–MS and LC–NMR. Anal. Chim. Acta 2012, 732, 145–152. [Google Scholar] [CrossRef]
- Hilbert, G.; Temsamani, H.; Bordenave, L.; Pedrot, E.; Chaher, N.; Cluzet, S.; Delaunay, J.-C.; Ollat, N.; Delrot, S.; Merillon, J.-M.; et al. Flavonol profiles in berries of wild Vitis accessions using liquid chromatography coupled to mass spectrometry and nuclear magnetic resonance spectrometry. Food Chem. 2015, 169, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Drake, B.G.; Gonzalez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.D.; Tjoelker, M.G.; Ellsworth, D.S.; Reich, P.B. Leaf gas exchange responses of 13 praire grassland species to elevated CO2 and increased nitrogen supply. New Phytol. 2001, 150, 405–418. [Google Scholar] [CrossRef] [Green Version]
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising atmospheric carbon dioxide: Plants face the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef]
- Oliveira, V.F.; Zaidan, L.B.P.; Braga, M.R.; Aidar, M.P.M.; Carvalho, M.A.M. Elevated CO2 atmosphere promotes plant growth and inulin production in the cerrado species Vernonia herbacea. Funct. Plant Biol. 2010, 37, 223–231. [Google Scholar] [CrossRef]
- Campbell, W.J.; Allen, L.H., Jr.; Bowes, G. Response of soybean canopy photosynthesis to CO2 concentration, light, and temperature. J. Exp. Bot. 1990, 41, 427–433. [Google Scholar] [CrossRef]
- Maroco, J.P.; Breia, E.; Faria, T.; Pereira, J.S.; Chaves, M.M. Effects of long-term exposure to elevated CO2 and N fertilization on the development of photosynthetic capacity and biomass accumulation in Quercus suber L. Plant Cell Environ. 2002, 25, 105–113. [Google Scholar] [CrossRef]
- Jifon, J.L.; Wolfe, D.W. Photosynthetic acclimation to elevated CO2 in Phaseolus vulgaris L. is altered by growth response to nitrogen supply. Glob. Chang. Biol. 2002, 8, 1018–1027. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Pérez, P.; Hernández, L.; Irigoyen, J.J.; Zita, G.; Martínez-Carrasco, R.; Sánchez-Díaz, M. The response of nodulated alfalfa to water supply, temperature and elevated CO2: Photosynthetic down-regulation. Physiol. Plantarum 2005, 123, 348–358. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Pardo, A.; Biel, C.; Savé, R.; Azcón-Bieto, J.; Nogués, S. Leaf carbon management in slow-growing plants exposed to elevated CO2. Glob. Chang. Biol. 2009, 15, 97–109. [Google Scholar] [CrossRef]
- Erice, G.; Irigoyen, J.J.; Pérez, P.; Martínez-Carrasco, R.; Sánchez-Díaz, M. Effect of elevated CO2, temperature and drought on dry matter partitioning and photosynthesis before and after cutting of nodulated alfalfa. Plant Sci. 2006, 170, 1059–1067. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Irigoyen, J.J.; Goicoechea, N.; Antolín, M.C.; Pascual, I.; Sánchez-Díaz, M.; Aguirreolea, J.; Morales, F. Growth, photosynthetic acclimation and yield quality in legumes grown under climate change simulations: An updated survey. Plant Sci. 2014, 226, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Aranjuelo, I.; Irigoyen, J.J.; Sánchez-Díaz, M. Effect of elevated temperature and water availability on CO2 exchange and nitrogen fixation on nodulated alfalfa plants. Environ. Exp. Bot. 2007, 59, 99–108. [Google Scholar] [CrossRef]
- Arrizabalaga-Arriazu, M.; Morales, F.; Irigoyen, J.J.; Hilbert, G.; Pascual, I. Growth performance and carbon partitioning of grapevine Tempranillo clones under simulated climate change scenarios: Elevated CO2 and temperature. J. Plant Physiol. 2020, 252, 153226. [Google Scholar] [CrossRef] [PubMed]
- Leibar, U.; Aizpurua, A.; Unamunzaga, O.; Pascual, I.; Morales, F. How will climate change influence grapevine cv. Tempranillo photosynthesis under different soil textures? Photosynth. Res. 2015, 124, 199–215. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Davey, P.A.; Bernacchi, C.J.; Dermody, O.C.; Heaton, E.A.; Moore, D.J.; Morgan, P.B.; Naidu, S.L.; Ra, H.S.Y.; Zhu, X.G.; et al. A meta-analysis of elevated [CO2] effects on soybean (Glycine max) physiology, growth and yield. Glob. Chang. Biol. 2002, 8, 695–709. [Google Scholar] [CrossRef]
- Urban, O. Physiological impacts of elevated CO2 concentration ranging from molecular to whole plant responses. Photosynthetica 2003, 41, 9–20. [Google Scholar] [CrossRef]
- Sánchez-Díaz, M.; Irigoyen, J.J.; Gómez-Casanovas, N.; Pardo, A.; Azcón-Bieto, J. El cambio climático global. Efecto previsible del CO2 sobre los vegetales. In La Ecofisiología Vegetal. Una Ciencia de Síntesis; Reigosa, M., Pedrol, N., Sánchez-Moreiras, A., Eds.; Universidad de Vigo: Vigo, Spain, 2004. [Google Scholar]
- Bunce, J.A. Direct and acclimatory responses of stomatal conductance to elevated carbon dioxide in four herbaceous crop species in the field. Glob. Chang. Biol. 2001, 7, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Goicoechea, N.; Baslam, M.; Erice, G.; Irigoyen, J.J. Increased photosynthetic acclimation in alfalfa associated with arbuscular mycorrhizal fungi (AMF) and cultivated in greenhouse under elevated CO2. J. Plant Physiol. 2014, 171, 1774–1781. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Loreto, F.; Delwiche, C.F. High carbon dioxide and sun/shade effects on isoprene emission from oak and aspen tree leaves. Plant Cell Environ. 1991, 14, 333–338. [Google Scholar] [CrossRef]
- Sanz-Sáez, Á.; Erice, G.; Aranjuelo, I.; Aroca, R.; Ruiz-Lozano, J.M.; Aguirreolea, J.; Irigoyen, J.J.; Sánchez-Díaz, M. Photosynthetic and molecular markers of CO2-mediated photosynthetic downregulation in nodulated alfalfa. J. Integ. Plant Biol. 2013, 55, 721–734. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Morales, F.; Abadía, A.; Abadía, J. Chlorophyll fluorescence and photon yield of oxygen evolution in iron-deficient sugar beet (Beta vulgaris L.) leaves. Plant Physiol. 1991, 97, 886–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, F.; Abadía, A.; Abadía, J. Photosynthesis, quenching of chlorophyll fluorescence and thermal energy dissipation in iron-deficient sugar beet leaves. Aust. J. Plant Physiol. 1998, 25, 403–412. [Google Scholar] [CrossRef]
- Geiger, M.; Haake, V.; Ludewig, F.; Sonnewald, U.; Stitt, M. The nitrate and ammonium nitrate supply have a major influence on the response of photosynthesis, carbon metabolism and growth to elevated carbon dioxide in tobacco. Plant Cell Environ. 1999, 22, 1177–1199. [Google Scholar] [CrossRef]
- Mishra, S.; Heckathorn, S.A.; Barua, D.; Wang, D.; Joshi, P.; Hamilton, E.W.; Frantz, J. Interactive effects of elevated CO2 and ozone on leaf thermotolerance in field-grown Glycine max. J. Integ. Plant Biol. 2008, 50, 1396–1405. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytic Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Bota, J.; Tomás, M.; Hernández, E.; García-Escudero, E.; Medrano, H. Exploring the genetic variability in water use efficiency: Evaluation of inter and intra cultivar genetic diversity in grapevines. Plant Sci. 2016, 251, 35–43. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flav (mg g−1 Skin DW) | Flav (mg g−1 Skin DW) | ||||||
|---|---|---|---|---|---|---|---|
| 2013–2015 | 2013 | 2014 | 2015 | ||||
| Tinto | T | ACO2 | FI | 2.61 ± 0.94 abc | 1.54 ± 0.08 bc | 1.79 ± 0.28 bc | 4.49 ± 0.98 ab |
| CD | 2.74 ± 0.76 ab | 2.22 ± 0.15 a | 1.76 ± 0.18 bc | 4.24 ± 0.78 abc | |||
| ECO2 | FI | 2.44 ± 0.68 abc | 1.53 ± 0.32 bc | 2.03 ± 0.42 bc | 3.76 ± 0.61 abc | ||
| CD | 3.06 ± 0.51 a | 2.30 ± 0.41 a | 2.84 ± 0.65 a | 4.04 ± 0.39 abc | |||
| T + 4 °C | ACO2 | FI | 2.47 ± 0.63 abc | 1.52 ± 0.05 bc | 2.20 ± 0.04 ab | 3.67 ± 0.47 a–d | |
| CD | 2.08 ± 0.71 abc | 1.15 ± 0.15 cd | 1.61 ± 0.28 bcd | 3.47 ± 0.92 a–f | |||
| ECO2 | FI | 2.41 ± 0.56 abc | 1.80 ± 0.34 ab | 1.91 ± 0.20 bc | 3.53 ± 0.67 a–e | ||
| CD | 2.69 ± 0.95 ab | 1.94 ± 0.26 ab | 1.56 ± 0.12 b–e | 4.57 ± 0.91 a | |||
| Blanco | T | ACO2 | FI | 0.94 ± 0.25 c | 0.53 ± 0.12 e | 0.89 ± 0.18 def | 1.39 ± 0.14 g |
| CD | 1.89 ± 0.51 abc | 1.48 ± 0.36 bc | 1.29 ± 0.25 c–f | 2.91 ± 0.31 b–g | |||
| ECO2 | FI | 1.14 ± 0.47 bc | 0.49 ± 0.06 e | 0.86 ± 0.05 def | 2.06 ± 0.34 d–g | ||
| CD | 1.33 ± 0.31 abc | 0.79 ± 0.12 de | 1.33 ± 0.31 c–f | 1.85 ± 0.30 fg | |||
| T + 4 °C | ACO2 | FI | 1.22 ± 0.32 bc | 0.64 ± 0.06 de | 1.29 ± 0.12 c–f | 1.74 ± 0.19 g | |
| CD | 0.93 ± 0.24 c | 0.59 ± 0.10 de | 0.81 ± 0.05 ef | 1.39 ± 0.24 g | |||
| ECO2 | FI | 1.14 ± 0.45 bc | 0.56 ± 0.14 de | 0.83 ± 0.15 ef | 2.02 ± 0.37 efg | ||
| CD | 1.32 ± 0.74 bc | 0.43 ± 0.04 e | 0.73 ± 0.15 f | 2.79 ± 0.26 c–g | |||
| Cultivar | 0.000 | <0.0001 | <0.0001 | <0.0001 | |||
| CO2 | 0.788 | 0.838 | 0.662 | 0.559 | |||
| Temp | 0.442 | 0.012 | 0.087 | 0.491 | |||
| Irrigation | 0.491 | 0.011 | 0.876 | 0.255 | |||
| Cultivar × CO2 | 0.746 | 0.017 | 0.161 | 0.585 | |||
| Cultivar × Temp | 0.833 | 0.901 | 0.684 | 0.657 | |||
| Cultivar × Irrigation | 0.874 | 0.862 | 0.679 | 0.701 | |||
| CO2 × Temp | 0.660 | 0.087 | 0.042 | 0.088 | |||
| CO2 × Irrigation | 0.724 | 0.888 | 0.150 | 0.608 | |||
| Temp × Irrigation | 0.384 | 0.001 | 0.005 | 0.964 | |||
| Cultivar × CO2 × Temp | 0.916 | 0.545 | 0.295 | 0.939 | |||
| Cultivar × CO2 × Irrigation | 0.554 | 0.122 | 0.557 | 0.297 | |||
| Cultivar × Temp × Irrigation | 0.872 | 0.782 | 0.790 | 0.457 | |||
| CO2 × Temp × Irrigation | 0.562 | 0.252 | 0.781 | 0.123 | |||
| Cultivar × CO2 × Temp × Irrigation | 0.663 | 0.858 | 0.394 | 0.349 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kizildeniz, T.; Pascual, I.; Hilbert, G.; Irigoyen, J.J.; Morales, F. Is Tempranillo Blanco Grapevine Different from Tempranillo Tinto Only in the Color of the Grapes? An Updated Review. Plants 2022, 11, 1662. https://doi.org/10.3390/plants11131662
Kizildeniz T, Pascual I, Hilbert G, Irigoyen JJ, Morales F. Is Tempranillo Blanco Grapevine Different from Tempranillo Tinto Only in the Color of the Grapes? An Updated Review. Plants. 2022; 11(13):1662. https://doi.org/10.3390/plants11131662
Chicago/Turabian StyleKizildeniz, Tefide, Inmaculada Pascual, Ghislaine Hilbert, Juan José Irigoyen, and Fermín Morales. 2022. "Is Tempranillo Blanco Grapevine Different from Tempranillo Tinto Only in the Color of the Grapes? An Updated Review" Plants 11, no. 13: 1662. https://doi.org/10.3390/plants11131662