
Abscisic Acid and Chitosan Modulate Polyphenol Metabolism and Berry Qualities in the Domestic White-Colored Cultivar Savvatiano

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Climate Conditions
2.2. Berry Ripening and Conventional Must Analysis
2.2.1. Berry Size
2.2.2. Total Soluble Solids
2.2.3. pH and Total Acidity
2.3. Metabolic Changes in Grape Berries
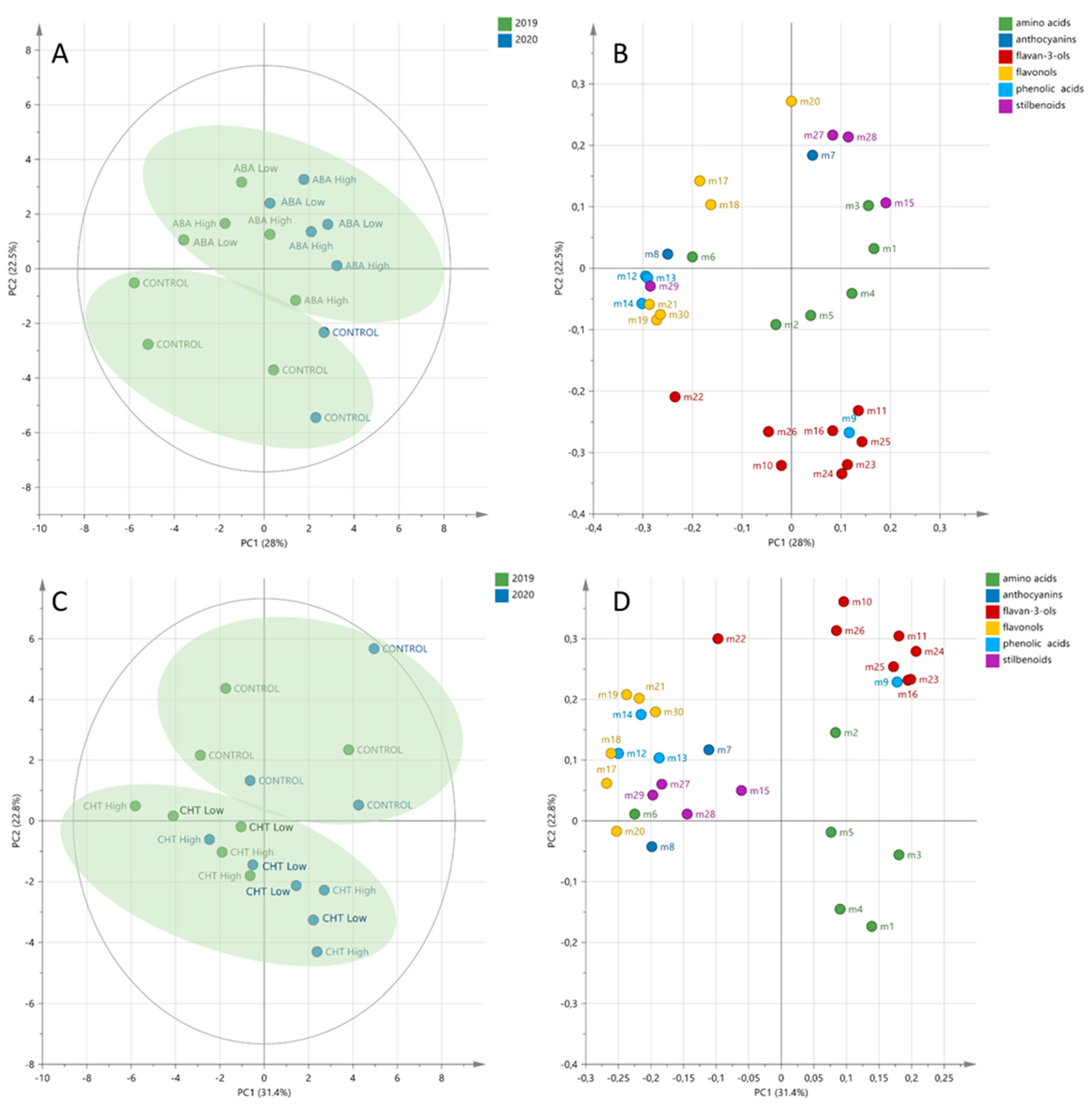
2.3.1. Multivariate Statistics
2.3.2. Univariate Statistics
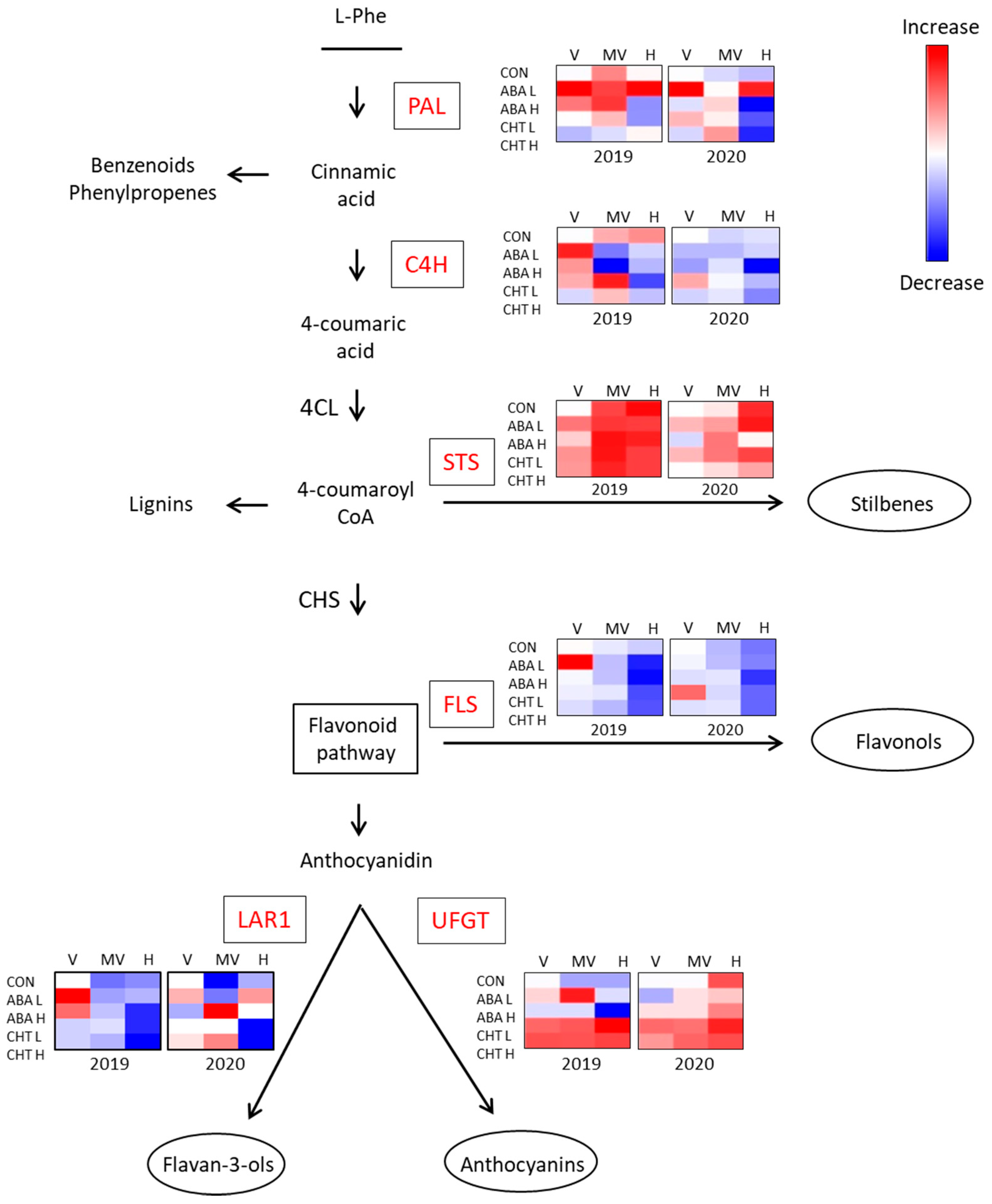
2.4. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Sampling and Physicochemical Determination in Grapes
4.3. UPLC–MS-Based Metabolic Profiling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No | Compound Assignement | RT (min) | Compound Class | Molecular Ion Adducts ES+ | In Source Fragment ES+ | Molecular Ion Adducts ES− | In Source Fragment ES− | λmax (nm) | References |
|---|---|---|---|---|---|---|---|---|---|
| m1 | L-proline | 1.01 | Amino acid | 116 [M+H]+ | 225, 275 | Standard | |||
| m10 | citric acid | 1.43 | Organic acid | 191 [M-H]− | 173 [M-H2O-H]− 111 [173-O=C(OH)2]− | 200 | Standard | ||
| m5 | L-tyrosine | 1.51 | Amino acid | 182 [M+H]+ | 207 | Standard | |||
| m2 | L-leucine | 1.61 | Amino acid | 132 [M+H]+ | 86 [M-CH(CH3)2-2H+H]+ (immonium ion) | 201, 232 | Standard [75] | ||
| m3 | L-isoleucine | 1.74 | Amino acid | 132 [M+H]+ | 86 [M-CH(CH3)2-2H+H]+ (immonium ion) | 210, 268 | Standard [75] | ||
| m9 | gallic acid | 1.94 | Phenolic acid | 169 [M-H]− | 125 [M-CO2]− | 210, 271 | Standard [76] | ||
| m4 | L-phenylalanine | 2.72 | Amino acid | 166 [M+H]+ | 148 [M-H2O+H]+ 120 [M-H2O-CO+H]+ | 200, 280 | Standard | ||
| m13 | caftaric acid | 3.63 | Phenolic acid | 311 [M-H]− | 179 [M - tartaric acid]− 149 [tartaric acid - H]− 135 [caffeic acid - COO]− | 200, 229, 328 | Standard [77] [78] [79] | ||
| m6 | L-tryptophan | 3.82 | Amino acid | 205 [M+H]+ | 219, 269 | Standard | |||
| m22 | procyanidinB1 | 4.26 | Flavan-3-ol | 579 [M+H]+ | 427 [M+H-C8H8O3]+ (RDA) 291 [M+H-(epi)catechin]+ (QM) | 577 [M-H]− | 425 [M-H-C8H8O3]− (RDA) 407 [M-H-C8H8O3-H2O]− 289 [M-H-(epi)catechin]− (QM) | 280, 313 | Standard [77] [80] |
| m24 | procyanidinB3 | 4.58 | Flavan-3-ol | 579 [M+H]+ | 427 [M+H-C8H8O3]+ (RDA) 291 [M+H-(epi)catechin]+ (QM) | 577 [M-H]− | 425 [M-H-C8H8O3]− (RDA) 407 [M-H-C8H8O3-H2O]− 289 [M-H-(epi)catechin]− (QM) | 200, 275sh | Standard [77] [80] |
| m12 | coutaric acid | 4.66 | Phenolic acid | 295 [M-H]− | 163 [coumaric acid - H]− 149 [tartaric acid - H]− 119 [coumaric acid - COO]− | 205, 311 | Standard [81] | ||
| m10 | catechin | 4.77 | Flavan-3-ol | 291 [M+H]+ | 289 [M-H]− | 271 [M-H-H2O]− 245 [M-H-CO2]− 205 [M - A ring]− 203 [M-H-CO2-C2H2O]− | 229, 278 | Standard [82] | |
| m14 | fertaric acid | 4.98 | Phenolic acid | 325 [M-H]− | 193 [ferulic acid - H]− 149 [tartaric acid - H]− 134 [ferulic acid - COO - CH3]− | 221, 262, 340 | Standard [77] [78] [83] | ||
| m25 | procyanidinB4 | 5.2 | Flavan-3-ol | 579 [M+H]+ | 427 [M+H-C8H8O3]+ (RDA) 291 [M+H-(epi)catechin]+ (QM) | 577 [M-H]− | 425 [M-H-C8H8O3]− (RDA) 407 [M-H-C8H8O3-H2O]− 289 [M-H-(epi)catechin]− (QM) | 202, 264, 362sh | Standard [77] [80] |
| m23 | procyanidinB2 | 5.35 | Flavan-3-ol | 579 [M+H]+ | 427 [M+H-C8H8O3]+ (RDA) 291 [M+H-(epi)catechin]+ (QM) | 577 [M-H]− | 425 [M-H-C8H8O3]− (RDA) 407 [M-H-C8H8O3-H2O]− 289 [M-H-(epi)catechin]− (QM) | 200, 278 | Standard [77] [80] |
| m11 | epicatechin | 5.91 | Flavan-3-ol | 291 [M+H]+ | 289 [M-H]− | 271 [M-H-H2O]− 245 [M-H-CO2]− 205 [M - A ring]− 203 [M-H-CO2-C2H2O]− | 229, 278 | Standard [82] | |
| m26 | procyanidin gallate | 6.51 | Flavan-3-ol | 731 [M+H]+ | 507 | 729 [M-H]− | 505, 523, 577 [M-H-galloyl]− | 206, 276 | |
| m22 | myricetin-3-O-glucoside | 7.03 | Flavonol | 479 [M-H]− | 317 [M-H-glucose] | 206, 356 | [83] | ||
| m7 | cyanidin-3-O-(6-O-acetyl)-glucoside | 7.16 | Anthocyanin diOH | 493 [M+H]+ | 511 [M+H+H2O]+ 287 [M+H-glucose-acethyl]+ | 202, 264, 325 | Standard | ||
| m16 | catechin gallate | 7.73 | Flavan-3-ol | 441 | 289, 169, 125 | 207, 280 | [78] | ||
| m20 | quercetin-3-O-glucuronide | 8.02 | Flavonol | 479 [M+H]+ | 303 [M+H-glucuronic acid]+ | 477 [M-H]− | 301 [M-H-glucuronic acid]− 955 [2M-H]− | 256, 359 | Standard |
| m19 | quercetin-3-O-glucoside | 8.14 | Flavonol | 465 [M+H]+ | 303 [M+H-glucose]+ | 477; 463; 478; 941; 955 | 477; 463; 301; 478; 941; 955 | 205, 273, 251 | Standard |
| m28 | E-piceatannol | 8.77 | stilbenoid DP1 | 244 [M+H]+ | 242 [M-H]− | 207, 283 | [78] & [84] | ||
| m30 | kaempferol-3-O-rutinoside | 8.75 | Flavonol | 595 [M+H]+ | 449 [M-rhamnose+H]+ 287 [M-rutin+H]+ | 593 [M-H]− 609 [M+O]+ | 447 [M-rhamnose-H]− 285 [M-rutin-H]− 301, 271 | 224, 264, 345 | Standard |
| m15 | E-piceid | 9.42 | Stilbenoid glucoside | 389 [M-H]− | 227, 185 | 200, 218, 221 | Standard | ||
| m8 | peonidin-3-O-(6-p-coumaroyl-glucoside)) | 9.79 | Anthocyanin diOH | 609 [M+H]+ | 607 [M-H]− | 205, 283 | Standard | ||
| m27 | E-resveratrol | 11.13 | stilbenoid DP1 | 229 [M+H]+ | 227 [M-H]− | 143, 185 | 203, 279 | Standard | |
| m29 | E-ε-viniferin | 12.53 | Stilbenoid DP2 | 455 [M+H]+ | 453 [M-H]− | 347, 359, 225 | 225sh, 323 | Standard [84] |
4.4. RNA Extraction and Gene Expression Analysis by RT-qPCR
4.5. Statistical Analysis
4.6. Meteorological Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hellenic Statistical Authority, Research of Viticulture. Available online: https://www.statistics.gr/el/statistics/-/publication/SPG63 (accessed on 13 May 2022).
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate Change, Wine, and Conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [Green Version]
- Keller, M. Managing Grapevines to Optimise Fruit Development in a Challenging Environment: A Climate Change Primer for Viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–69. [Google Scholar] [CrossRef]
- Litskas, V.; Mandoulaki, A.; Vogiatzakis, I.N.; Tzortzakis, N.; Stavrinides, M. Sustainable Viticulture: First Determination of the Environmental Footprint of Grapes. Sustainability 2020, 12, 8812. [Google Scholar] [CrossRef]
- Dufour, M.C.; Lambert, C.; Bouscauta, J.; Merillon, J.M.; Corio-Costet, M.F. Benzothiadiazole-primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Delaunois, B.; Farace, G.; Jeandet, P.; Clément, C.; Baillieul, F.; Dorey, S.; Cordelier, S. Elicitors as Alternative Strategy to Pesticides in Grapevine? Current Knowledge on Their Mode of Action from Controlled Conditions to Vineyard. Environ Sci. Pollut. Res. 2014, 21, 4837–4846. [Google Scholar] [CrossRef]
- De Bona, G.S.; Vincenzi, S.; De Marchi, F.; Angelini, E.; Bertazzon, N. Chitosan induces delayed grapevine defense mechanisms and protects grapevine against Botrytis cinerea. J. Plant Dis. Prot. 2021, 128, 8. [Google Scholar] [CrossRef]
- Dong, T.; Zheng, T.; Fu, W.; Guan, L.; Jia, H.; Fang, J. The Effect of Ethylene on the Color Change and Resistance to Botrytis cinerea Infection in ‘Kyoho’ Grape Fruits. Foods 2020, 9, 892. [Google Scholar] [CrossRef]
- Vitalini, S.; Ruggiero, A.; Rapparini, F.; Neri, L.; Tonni, M.; Iriti, M. The application of chitosan and benzothiadiazole in vineyard (Vitis vinifera L. cv Groppello Gentile) changes the aromatic profile and sensory attributes of wine. Food Chem. 2014, 162, 192–205. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Cerdán, G.T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 15, 213–221. [Google Scholar] [CrossRef]
- Villalobos-Gonzalez, L.; Peña-Neira, A.; Ibanez, F.; Pastenes, C. Long term effect of abscisic acid (ABA) on the grape berry phenylpropanoids pathway: Gene expression and metabolic content. Plant Physiol. Biochem. 2016, 105, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Koyama, R.; Roberto, S.R.; de Souza, R.T.; Borges, W.F.S.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous Abscisic Acid Promotes Anthocyanin Biosynthesis and Increased Expression of Flavonoid Synthesis Genes in Vitis vinifera X Vitis labrusca Table Grapes in a Subtropical Region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, V.; Unlubayir, M.; Teixeira, A.; Geros, H.; Lanoue, A. Calcium and methyl jasmonate cross-talk in the secondary metabolism of grape cells. Plant Physiol Biochem 2021, 165, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. Int. J. Mol. Sci. 2020, 21, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.L.; Inês, A.; Oliveira, A.A.; Falco, V. Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) Tissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, J.A.; Saucier, C.; Glories, Y. Grape and Wine Phenolics: History and Perspective. Am. J. Enol. Vitic. 2006, 57, 239–248. [Google Scholar]
- Teixeira, A.; Eiras-Dias, J.; Castellarin, S.; Gerós, H. Berry phenolics of grapevine under challenging environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Guo, A.; Wang, H. Mechanisms of Oxidative Browning of Wine. Food Chem. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Bishayee, A.; Barnes, K.F.; Bhatia, D.; Darvesh, A.S.; Carroll, R.T. Resveratrol suppresses oxidative stress and inflammatory response in diethylnitrosamine-initiated rat hepatocarcinogenesis. Cancer Prev. Res. 2010, 3, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Gülçin, İ. Antioxidant properties of resveratrol: A structure-activity insight. Innov. Food Sci. Emerg. Technol. 2010, 11, 210–218. [Google Scholar] [CrossRef]
- Wu, J.M.; Hsieh, T. Resveratrol: A cardioprotective substance. Ann. N. Y. Acad. Sci. 2011, 1215, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T.; Szkudelska, K. Resveratrol and diabetes: From animal to human studies. Biochim. Biophys. Acta 2015, 1852, 1145–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aluyen, J.K.; Ton, Q.N.; Tran, T.; Yang, A.E.; Gottlieb, H.B.; Bellanger, R.A. Resveratrol: Potential as anticancer agent. J. Diet. Suppl. 2012, 9, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.A. Anti-aging properties of resveratrol: Review and report of a potent new antioxidant skin care formulation. J. Cosmet. Dermatol. 2008, 7, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O. Phenolics and ripening in grape berries. Am. J. Enol. Vitic. 2006, 57, 249–256. [Google Scholar]
- Aleixandre-Tudo, J.L.; Toit, W.D. The Role of UV-Visible Spectroscopy for Phenolic Compounds Quantification in Winemaking. In Frontiers and New Trends in the Science of Fermented Food and Beverages; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Sacchi, K.L.; Bisson, L.F.; Adams, D.O. A Review of the Effect of Winemaking Techniques on Phenolic Extraction in Red Wines. Am J Enol Vitic. 2005, 56, 197–206. [Google Scholar]
- Belhadj, A.; Saigne, C.; Telef, N.; Cluzet, S.; Bouscaut, J.; Corio-Costet, M.F.; Mérillon, J.M. Methyl jasmonate induces defense responses in grapevine and triggers protection against Erysiphe necator. J. Agric. Food Chem. 2006, 54, 9119–9125. [Google Scholar] [CrossRef]
- Iriti, M.; Rossoni, M.; Borgo, M.; Faoro, F. Benzothiadiazole enhances resveratrol and anthocyanin biosynthesis in grapevine, meanwhile improving resistance to Botrytis cinerea. J. Agric. Food Chem. 2004, 52, 4406–4413. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; Tommaso, G.; Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Improving phenolic and chromatic characteristics of Monastrell, Merlot and Syrah wines by using methyl jasmonate and benzothiadiazole.J. Int. Sci. Vigne Vin 2017, 51, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, A.R.; Mulinacci, N.; Valletta, A.; Innocenti, M.; Pasqua, G. Effects of elicitors on the production of resveratrol and viniferins in cell cultures of Vitis vinifera L. cv Italia. J. Agric. Food Chem. 2011, 59, 9094–9101. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-García, Y.; López-Roca, J.M.; Bautista-Ortín, A.B.; Gil-Muñoz, R.; Gómez-Plaza, E. Effect of combined use of benzothiadiazoleand methyl jasmonate on volatile compounds of Monastrell wine. Am. J. Enol. Vitic. 2014, 65, 238–243. [Google Scholar] [CrossRef]
- Peppi, M.C.; Walker, M.A.; Fidelibus, M.W. Application of abscisic acid rapidly upregulated UFGT gene expression and improved color of grape berries. Vitis J. Grapevine Res. 2008, 47, 11–14. [Google Scholar]
- Silva, V.; Singh, R.K.; Gomes, N.; Soares, B.G.; Silva, A.; Falco, V.; Capita, R.; Alonso-Calleja, C.; Pereira, J.E.; Amaral, J.S.; et al. Comparative Insight upon Chitosan Solution and Chitosan Nanoparticles Application on the Phenolic Content, Antioxidant and Antimicrobial Activities of Individual Grape Components of Sousão Variety. Antioxidants 2020, 9, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deytieux, C.; Gagné, S.; L’Hyvernay, A.; Donèche, B.; Geny, L. Possible roles of both abscisic acid and indol acetic acid in controlling grape berry ripening process. J. Int. Sci. Vigne. Vin. 2007, 41, 141–148. [Google Scholar] [CrossRef]
- Jeong, S.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, M. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins. Plant Sci. 2004, 167, 247–252. [Google Scholar] [CrossRef]
- Ju, Y.-L.; Liu, M.; Zhao, H.; Meng, J.-F.; Fang, Y.-L. Effect of Exogenous Abscisic Acid and Methyl Jasmonate on Anthocyanin Composition, Fatty Acids, and Volatile Compounds of Cabernet Sauvignon (Vitis Vinifera L.) Grape Berries. Molecules 2016, 21, 1354. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Gamboa, G.; Pérez-Álvarez, E.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Changes on grape volatile composition through elicitation with methyl jasmonate, chitosan, and a yeast extract in Tempranillo (Vitis vinifera L.) grapevines. Sci. Hortic. 2019, 244, 257–262. [Google Scholar] [CrossRef]
- Sandhu, A.K.; Gray, D.J.; Lu, J.; Gu, L. Effects of exogenous abscisic acid on antioxidant capacities, anthocyanins, and flavonol contents of Muscadine grape (Vitis rotundifolia) skins. Food Chem. 2011, 126, 982–988. [Google Scholar] [CrossRef]
- Peppi, M.C.; Fidelibus, M.W.; Dokoozlian, N. Abscisic acid application timing and concentration affect firmness, pigmentation, and color of ‘Flame Seedless’ grapes. Hort. Sc. 2006, 41, 1440–1445. [Google Scholar] [CrossRef] [Green Version]
- Peppi, M.C.; Fidelibus, M.W.; Dokoozlian, N.K. Application timing and concentration of abscisic acid affect the quality of ‘Redglobe’ grapes. J. Hortic. Sci. Biotechnol. 2007, 82, 304–310. [Google Scholar] [CrossRef]
- Ferrara, G.; Mazzeo, A.; Matarrese, A.M.S.; Pacucci, C.; Punzi, R.; Faccia, M.; Trani, A.; Gambacorta, G. Application of abscisic acid (S-ABA) and sucrose to improve colour, anthocyanin content and antioxidant activity of cv. Crimson Seedless grape berries. J. Grape Wine Res. 2015, 21, 18–29. [Google Scholar] [CrossRef]
- Reynolds, A.; Robbins, N.; Lee, H.S.; Kotsaki, E. Impacts and interactions of abscisic acid and gibberellic acid on sovereign Coronation and Skookum seedless table grapes. Am. J. Enol. Vitic. 2016, 67, 327–338. [Google Scholar] [CrossRef]
- Ranjbaran, E.; Gholami, M.; Jensen, M. Improvement of grape phenolic content and antioxidant activity by pre-harvest application of methyl jasmonate. Acta Hortic. 2020, 1292, 405–410. [Google Scholar] [CrossRef]
- Cataldo, E.; Salvi, L.; Paoli, F.; Fucile, M.; Mattii, G.B. Effect of Agronomic Techniques on Aroma Composition of White Grapevines: A Review. Agronomy 2021, 11, 2027. [Google Scholar] [CrossRef]
- González, R.; González, M.-R.; Martín, P. Abscisic Acid and Ethephon Treatments Applied to ‘Verdejo’ White Grapes Affect the Quality of Wine in Different Ways. Sci. Agric. (Piracicaba Braz.) 2018, 75, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic Acid Is a Major Regulator of Grape Berry Ripening Onset: New Insights into ABA Signaling Network. Front. Plant Sci. 2017, 8, 1093. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Khalil-Ur-Rehman, M.; Wei, L.L.; Nieuwenhuizen, N.J.; Zheng, H.; Tao, J.M. Effect of Thidiazuron on Terpene Volatile Constituents and Terpenoid BiosynthesisPathway Gene Expression of Shine Muscat (Vitis labrusca × V. vinifera) Grape Berries. Molecules 2020, 25, 2578. [Google Scholar] [CrossRef]
- Deluc, L.G.; Quilici, D.R.; Decendit, A.; Grimplet, J.; Wheatley, M.D.; Schlauch, K.A.; Mérillon, J.-M.; Cushman, J.C.; Cramer, G.R. Water deficit alters differentially metabolic pathways affecting important flavor and quality traits in grape berries of Cabernet Sauvignon and Chardonnay. BMC Genom. 2009, 10, 212. [Google Scholar] [CrossRef] [Green Version]
- Vannozzi, A.; Dry, I.B.; Fasoli, M.; Zenoni, S.; Lucchin, M. Genome-wide analysis of the grapevine stilbene synthase multigenic family: Genomic organization and expression profiles upon biotic and abiotic stresses. BMC Plant Biol. 2012, 12, 130. [Google Scholar] [CrossRef]
- Czemmel, S.; Stracke, R.; Weisshaar, B.; Cordon, N.; Harris, N.N.; Walker, A.R.; Robinson, S.P.; Bogs, J. The grapevine R2R3-MYB transcription factor VvMYBF1 regulates flavonol synthesis in developing grape berries. Plant Physiol. 2009, 151, 1513–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boss, P.K.; Davies, C.; Robinson, S.P. Expression of anthocyanin biosynthesis pathway genes in red and white grapes. Plant Mol. Biol. 1996, 32, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocco, M.; Mercenaro, L.; Lo Cascio, M.; Nieddu, G. Effects of Vine Water Status and Exogenous Abscisic Acid on Berry Composition of Three Red Wine Grapes Grown under Mediterranean Climate. Horticulturae 2020, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Arapitsas, P.; Oliveira, J.; Mattivi, F. Do White Grapes Really Exist? Food Res. Int. 2015, 69, 21–25. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Minute, F.; Giotto, F.; Filipe-Ribeiro, L.; Cosme, F.; Nunes, F.M. Is Pinking Susceptibility Index a Good Predictor of White Wines Pinking Phenomena? Food Chem. 2022, 386, 132861. [Google Scholar] [CrossRef]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.J.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. Cell Mol. Biol. 2007, 49, 772–785. [Google Scholar] [CrossRef]
- Ferreira, V.; Matus, J.T.; Pinto-Carnide, O.; Carrasco, D.; Arroyo-García, R.; Castro, I. Genetic Analysis of a White-to-Red Berry Skin Color Reversion and Its Transcriptomic and Metabolic Consequences in Grapevine (Vitis Vinifera Cv. ‘Moscatel Galego’). BMC Genom. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Tran-scriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 67. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lüscher, J.; Brillante, L.; Kurtural, S.K. Flavonol profile is a reliable indicator to assess canopy architecture and the exposure of red wine grapes to solar radiation. Front. Plant Sci. 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Balint, G.; Reynolds, A.G. Impact of exogenous abscisic acid on vine physiology and grape composition of Cabernet Sauvignon. Am. J. Enol. Vitic. 2013, 64, 74–87. [Google Scholar] [CrossRef]
- Andrea-Silva, J.; Cosme, F.; Ribeiro, L.F.; Moreira, A.S.P.; Malheiro, A.C.; Coimbra, M.A.; Domingues, M.R.M.; Nunes, F.M. Origin of the Pinking Phenomenon of White Wines. J. Agric. Food Chem. 2014, 62, 5651–5659. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, Q.; Xi, B.; Dai, H. Study on the regulation of anthocyanin biosynthesis by exogenous abscisic acid in grapevine. Sci. Hortic. 2019, 2, 294–301. [Google Scholar] [CrossRef]
- Degu, A.; Ayenew, B.; Cramer, G.R.; Fait, A. Polyphenolic responses of grapevine berries to light, temperature, oxidative stress, abscisic acid and jasmonic acid show specific developmental-dependent degrees of metabolic resilience to perturbation. Food Chem. 2016, 212, 828–836. [Google Scholar] [CrossRef]
- Freitas, P.M.; López-Gálvez, F.; Tudela, J.A.; Gil, M.I.; Allende, A. Postharvest treatment of table grapes with ultraviolet-C and chitosan coating preserves quality and increases stilbene content. Postharvest Biol. Technol. 2015, 105, 51–57. [Google Scholar] [CrossRef]
- Ferri, M.; Tassoni, A.; Franceschetti, M.; Righetti, L.; Naldrett, M.J.; Bagni, N. Chitosan treatment induces changes of protein expression profile and stilbene distribution in Vitis vinifera cell suspensions. Proteomics 2009, 9, 610–624. [Google Scholar] [CrossRef]
- Mihai, R.; Cristina, S.; Helepciuc, F.; Brezeanu, A.; Stoian, G. Biotic and Abiotic Elicitors Induce Biosynthesis and Accumulation of Resveratrol with Antitumoral Activity in the Long - Term Vitis Vinifera L. Callus Cultures. Rom. Biotechnol. Lett. 2011, 16, 7. [Google Scholar]
- Righetti, L.; Franceschetti, M.; Ferri, M.; Tassoni, A.; Bagni, N. Resveratrol production in Vitis vinifera cell suspensions treated with several elicitors. Caryologia 2007, 60, 169–171. [Google Scholar] [CrossRef] [Green Version]
- Valletta, A.; Iozia, L.M.; Leonelli, F. Impact of Environmental Factors on Stilbene Biosynthesis. Plants 2021, 10, 90. [Google Scholar] [CrossRef]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth stages of the grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp.vinifera). Codes and descriptions according to the extended BBCH scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- OIV. Compendium of International Methods of Analysis of Wines and Musts; Methods: OIVMA–AS313–01; OIV–MA–AS2–10; International Organisation of Vine and Wine: Paris, France, 2018; Volume 2, Available online: http://www.oiv.int/oiv/info/enmethodes (accessed on 12 May 2022).
- Billet, K.; Delanoue, G.; Arnault, I.; Besseau, S.; Oudin, A.; Courdavault, V.; Marchand, P.A.; Giglioli-Guivarc’h, N.; Guérin, L.; Lanoue, A. Vineyard evaluation of stilbenoid-rich grape cane extracts against downy mildew: A large-scale study. Pest Manag. Sci. 2018, 75, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Vecchi, M.M.; Wen, D. Distinguishing between Leucine and Isoleucine by Integrated LC–MS Analysis Using an Orbitrap Fusion Mass Spectrometer. Anal. Chem. 2016, 88, 10757–10766. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, A.; Kumar, B. Identification and characterization of phenolics and terpenoids from ethanolic extracts of “Phyllanthus” species by HPLC-ESI-QTOF-MS/MS. J. Pharm. Anal. 2017, 7, 214–222. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Arapitsas, P.; Stefanini, M.; Flick, G.; Mattivi, F. Analysis of the phenolic composition of fungus-resistant grape varieties cultivated in Italy and Germany using UHPLC-MS/MS. J. Mass Spectrom. 2014, 49, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Vrhovsek, U.; Masuero, D.; Gasperotti, M.; Franceschi, P.; Caputi, L.; Viola, R.; Mattivi, F. A versatile targeted metabolomics method for the rapid quantification of multiple classes of phenolics in fruits and beverages. J. Agric. Food Chem. 2012, 60, 8831–8840. [Google Scholar] [CrossRef]
- Khoza, B.S.; Gbashi, S.; Steenkamp, P.A.; Njobeh, P.B.; Madala, N.E. Identification of hydroxylcinnamoyl tartaric acid esters in ”Bidens pilosa” by UPLC-tandem mass spectrometry. South Afr. J. Bot. 2016, 103, 95–100. [Google Scholar] [CrossRef]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Lambert, M.; Meudec, E.; Verbaere, A.; Mazerolles, G.; Wirth, J.; Masson, G.; Sommerer, N. A high-throughput UHPLC-QqQ-MS method for polyphenol profiling in rosé wines. Molecules 2015, 20, 7890–7914. [Google Scholar] [CrossRef]
- Stoggl, W.M.; Huck, C.W.; Bonn, G.K. Structural elucidation of catechin and epicatechin in sorrel leaf extracts using liquid-chromatography coupled to diode array-, fluorescence-, and mass spectrometric detection. J. Sep. Sci. 2004, 27, 524–528. [Google Scholar] [CrossRef]
- Sobral, F.; Calhelha, R.C.; Barros, L.; Dueñas, M.; Tomás, A.; Santos-Buelga, C.; Vilas-Boas, M.; Ferreira, I.C.F.R. Flavonoid Composition and Antitumor Activity of Bee Bread Collected in Northeast Portugal. Molecules 2017, 22, 248. [Google Scholar] [CrossRef] [Green Version]
- Moss, R.; Mao, Q.; Taylor, D.; Saucier, C. Investigation of monomeric and oligomeric wine stilbenoids in red wines by ultra-high-performance liquid chromatography/electrospray ionization quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alatzas, A.; Theocharis, S.; Miliordos, D.-E.; Leontaridou, K.; Kanellis, A.K.; Kotseridis, Y.; Hatzopoulos, P.; Koundouras, S. The effect of water deficit on two Greek Vitis vinifera L. cultivars: Physiology, grape composition and gene expression during berry development. Plants 2021, 10, 1947. [Google Scholar] [CrossRef] [PubMed]
- Lagouvardos, K.; Kotroni, V.; Bezes, A.; Koletsis, I.; Kopania, T.; Lykoudis, S.; Mazarakis, N.; Papagiannaki, K.; Vougioukas, S. The Automatic Weather Stations NOANN Network of the National Observatory of Athens: Operation and Database. Geosci. Data J. 2017, 4, 4–16. [Google Scholar] [CrossRef]




| 2019 | |||||
|---|---|---|---|---|---|
| Stage | Treatment | Weight/Berry (g) | Total Soluble Solids (°Brix) | pH | Total Acidity (Tart. Ac. g/L) |
| Veraison | Control | 2.22 ± 0.29 b | 14.26 ± 0.75 b | 2.94 ± 0.22 a | 5.42 ± 0.24 b |
| ABA Low | 2.50 ± 0.17 ab | 15.16 ± 0.40 a | 2.74 ± 0.01 a | 5.40 ± 0.07 b | |
| ABA High | 2.38 ± 0.15 ab | 15.06 ± 0.40 ab | 2.91 ± 0.15 a | 5.15 ± 0.15 b | |
| CHT Low | 2.62 ± 0.08 a | 15.33 ± 0.37 a | 2.89 ± 0.05 a | 5.32 ± 0.11 b | |
| CHT High | 2.62 ± 0.14 a | 15.60 ± 0.02 a | 2.78 ± 0.02 a | 5.82 ± 0.18 a | |
| Mid Veraison | Control | 2.70 ± 0.16 a | 17.13 ± 0.55 a | 3.14 ± 0.07 ab | 4.75 ± 0.11 a |
| ABA Low | 2.72 ± 0.11 a | 17.33 ± 0.20 a | 3.22 ± 0.06 ab | 4.32 ± 0.11 c | |
| ABA High | 2.32 ± 0.14 b | 17.63 ± 0.77 a | 3.26 ± 0.11 a | 4.60 ± 0.08 ab | |
| CHT Low | 2.44 ± 0.06 b | 17.20 ± 0.51 a | 3.11 ± 0.01 b | 4.77 ± 0.04 a | |
| CHT High | 2.07 ± 0.14 c | 17.80 ± 0.30 a | 3.21 ± 0.04 ab | 4.55 ± 0.11 b | |
| Harvest | Control | 2.30 ± 0.06 ab | 17.13 ± 0.55 a | 3.14 ± 0.07 ab | 4.75 ± 0.11 a |
| ABA Low | 2.55 ± 0.30 a | 17.33 ± 0.20 a | 3.22 ± 0.06 ab | 4.32 ± 0.11 c | |
| ABA High | 2.08 ± 0.13 b | 17.63 ± 0.77 a | 3.26 ± 0.11 a | 4.60 ± 0.08 ab | |
| CHT Low | 2.55 ± 0.19 a | 17.20 ± 0.51 a | 3.11 ± 0.01 b | 4.77 ± 0.04 a | |
| CHT High | 2.05 ± 0.17 b | 17.80 ± 0.30 a | 3.21 ± 0.04 ab | 4.55 ± 0.11 b | |
| 2020 | |||||
| Veraison | Control | 2.85 ± 0.11 a | 17.76 ± 0.28 a | 3.32 ± 0.09 ab | 5.30 ± 0.34 ab |
| ABA Low | 2.74 ± 0.02 a | 17.16 ± 0.50 ab | 3.21 ± 0.04 b | 5.55 ± 0.15 ab | |
| ABA High | 2.81 ± 0.11 a | 17.56 ± 0.83 ab | 3.73 ± 0.10 b | 5.15 ± 0.22 b | |
| CHT Low | 2.74 ± 0.26 a | 17.26 ± 0.41 ab | 3.29 ± 0.03 ab | 5.15 ± 0.17 b | |
| CHT High | 2.68 ± 0.08 a | 16.7 ± 0.55 b | 3.27 ± 0.04 ab | 5.70 ± 0.15 a | |
| Mid Veraison | Control | 2.47 ± 0.13 a | 19.33 ± 0.21 a | 3.36 ± 0.02 ab | 3.7 ± 0.1 b |
| ABA Low | 2.90 ± 0.04 a | 18.13 ± 0.46 c | 3.40 ± 0.02 a | 3.70 ± 0.0 ba | |
| ABA High | 2.72 ± 0.07 a | 18.63 ± 0.55 bc | 3.36 ± 0.05 ab | 3.90 ± 0.19 ab | |
| CHT Low | 2.61 ± 0.43 a | 19.06 ± 0.21 ab | 3.32 ± 0.02 b | 3.7b ± 0.19 ab | |
| CHT High | 2.77 ± 0.22 a | 18.81 ± 0.34a bc | 3.36 ± 0.03 ab | 4.17 ± 0.04 a | |
| Harvest | Control | 2.94 ± 0.22 bc | 20.33 ± 0.20 a | 3.42 ± 0.03 a | 4.35 ± 0.15 d |
| ABA Low | 3.08 ± 0.05 ab | 18.03 ± 0.45 b | 3.21 ± 0.07 c | 4.50 ± 0.08 cd | |
| ABA High | 3.26 ± 0.14 a | 19.23 ± 0.55 ab | 3.23 ± 0.07 c | 4.85 ± 0.01 cd | |
| CHT Low | 2.80 ± 0.14 c | 19.40 ± 1.57 ab | 3.36 ± 0.08 b | 4.76 ± 0.0 cb | |
| CHT High | 3.13 ± 0.11 ab | 19.16 ± 1.07 ab | 3.35 ± 0.03 b | 5.01 ± 0.15 a | |
| Gene Name | NCBI Accession Number | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|---|
| VviPAL | XM_010660093.2 | GTGAGGGAAGAACTGGGAGC | TTGTCACACTCTTCACCGGG | [86] |
| VviC4H | XM_002266202.3 | GAACCACCTGAACCTCTCCG | ATCCGAACTCCACTCCCTGA | [86] |
| VviSTS | X_76892 | ATCGAAGATCACCCACCTTG | CTTAGCGGTTCGAAGGACAG | [6] |
| VviFLS | XM_002285803.4 | TGGGGTTAGGTCTGGGAGAG | AACCTGCAAGCCCTGAACTT | [61] |
| VviUFGT | NM_001397857.1 | TGGTGGCTGACGCATTCAT | CCCCATCTCTGCTGCCATATC | [13] |
| VviLAR1 | NM_001280958.1 | CAGGAGGCTATGGAGAAGATAC | ACGCTTCTCTCTGTACATGTTG | [61] |
| VviEF1a | XM_002284888.3 | GAACTGGGTGCTTGATAGGC | AACCAAAATATCCGGAGTAAAAGA | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miliordos, D.E.; Alatzas, A.; Kontoudakis, N.; Kouki, A.; Unlubayir, M.; Gémin, M.-P.; Tako, A.; Hatzopoulos, P.; Lanoue, A.; Kotseridis, Y. Abscisic Acid and Chitosan Modulate Polyphenol Metabolism and Berry Qualities in the Domestic White-Colored Cultivar Savvatiano. Plants 2022, 11, 1648. https://doi.org/10.3390/plants11131648
Miliordos DE, Alatzas A, Kontoudakis N, Kouki A, Unlubayir M, Gémin M-P, Tako A, Hatzopoulos P, Lanoue A, Kotseridis Y. Abscisic Acid and Chitosan Modulate Polyphenol Metabolism and Berry Qualities in the Domestic White-Colored Cultivar Savvatiano. Plants. 2022; 11(13):1648. https://doi.org/10.3390/plants11131648
Chicago/Turabian StyleMiliordos, Dimitrios Evangelos, Anastasios Alatzas, Nikolaos Kontoudakis, Angeliki Kouki, Marianne Unlubayir, Marin-Pierre Gémin, Alexandros Tako, Polydefkis Hatzopoulos, Arnaud Lanoue, and Yorgos Kotseridis. 2022. "Abscisic Acid and Chitosan Modulate Polyphenol Metabolism and Berry Qualities in the Domestic White-Colored Cultivar Savvatiano" Plants 11, no. 13: 1648. https://doi.org/10.3390/plants11131648