
Integrative Taxonomy Reveals a New Species of the Genus Lejeunea (Marchantiophya: Lejeuneaceae) from Peninsular Malaysia

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
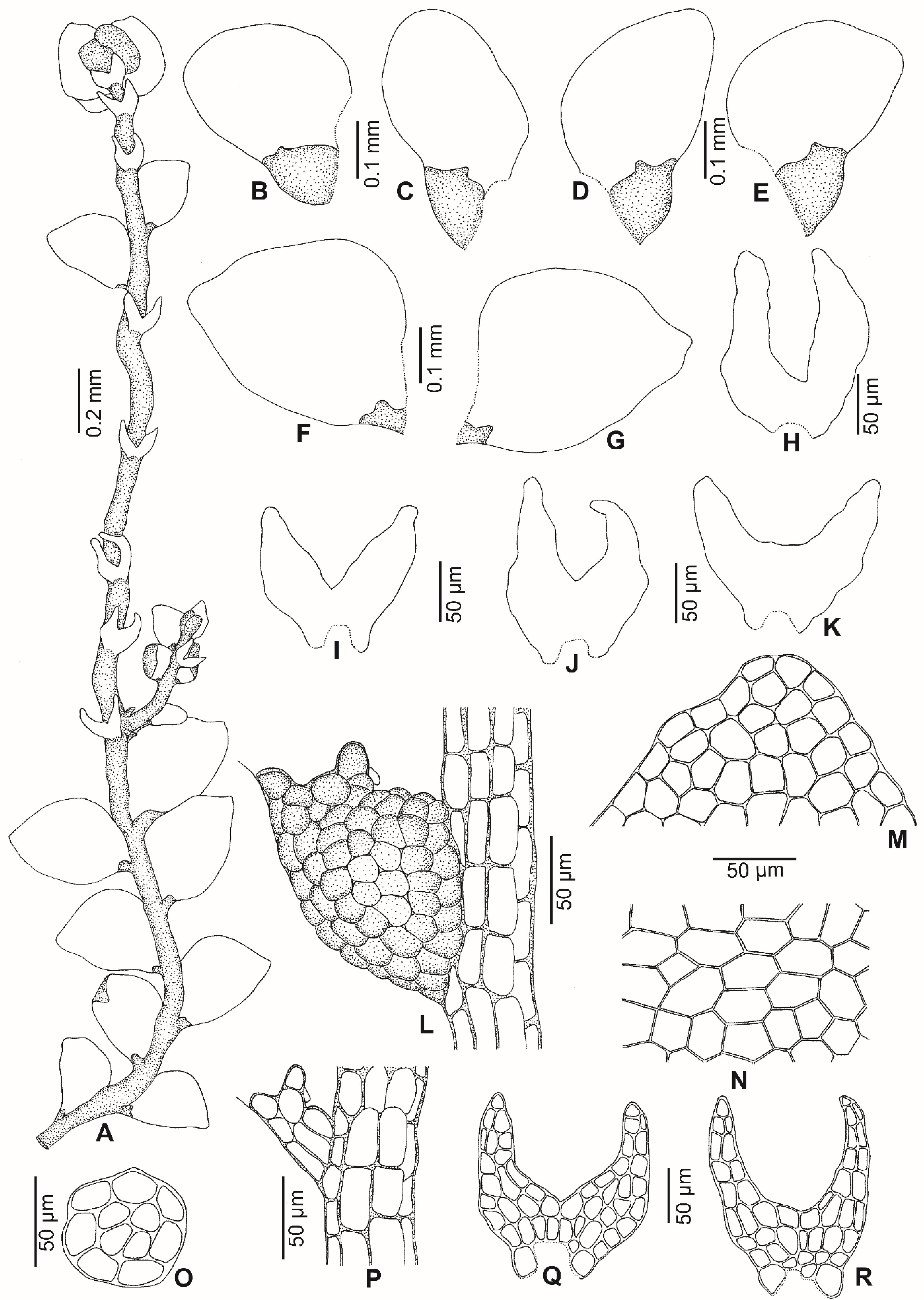
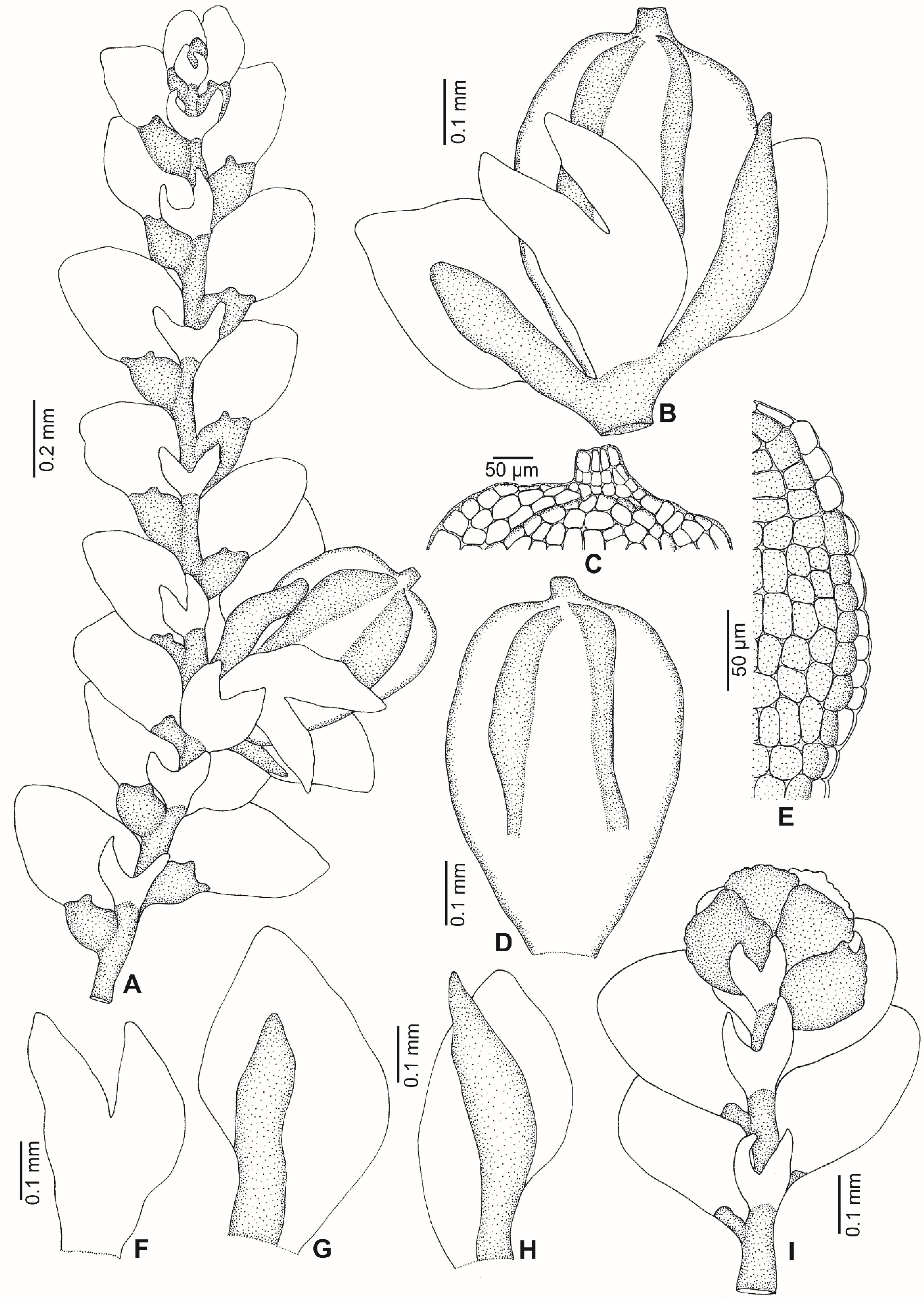
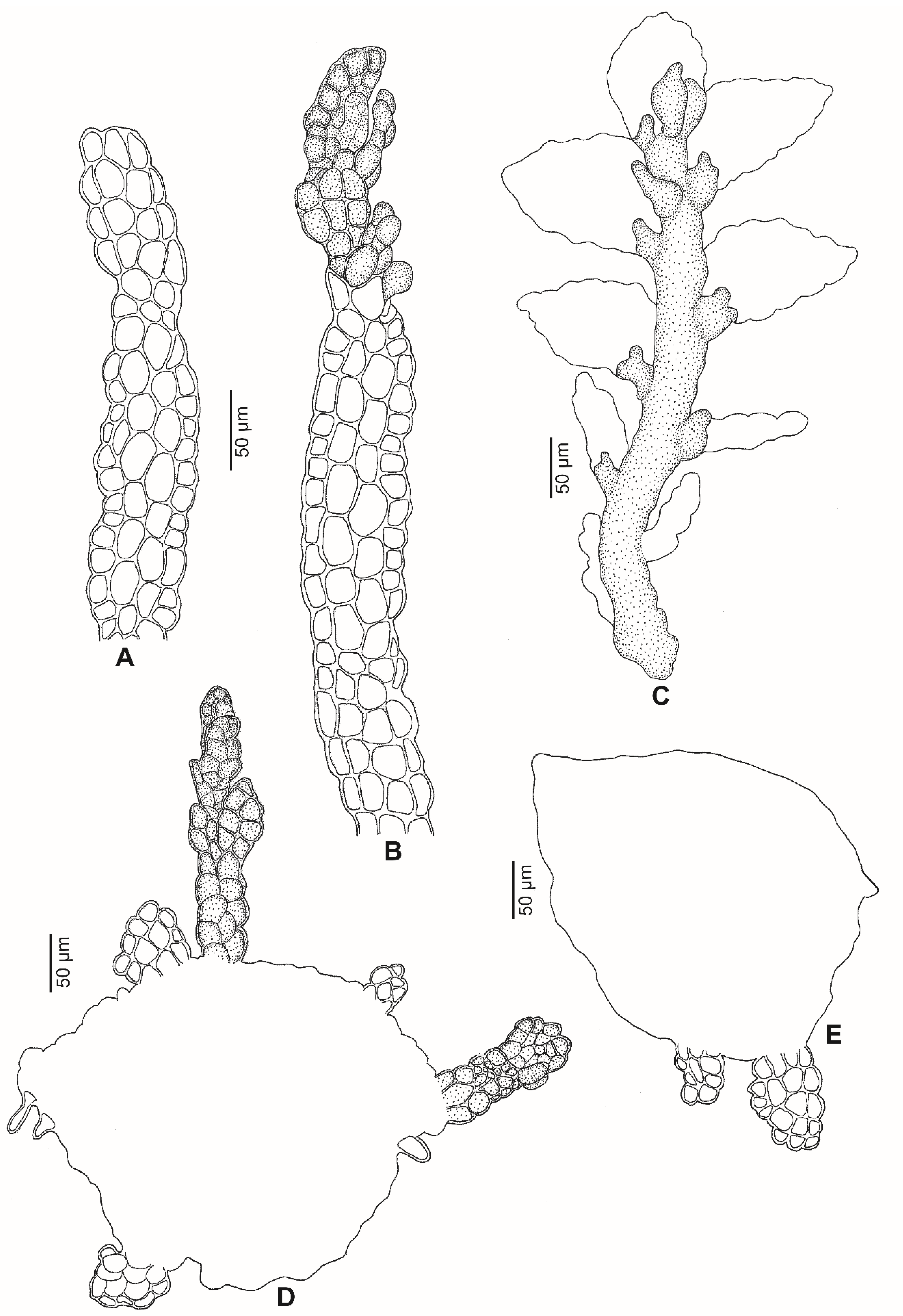
2.1. Morphology
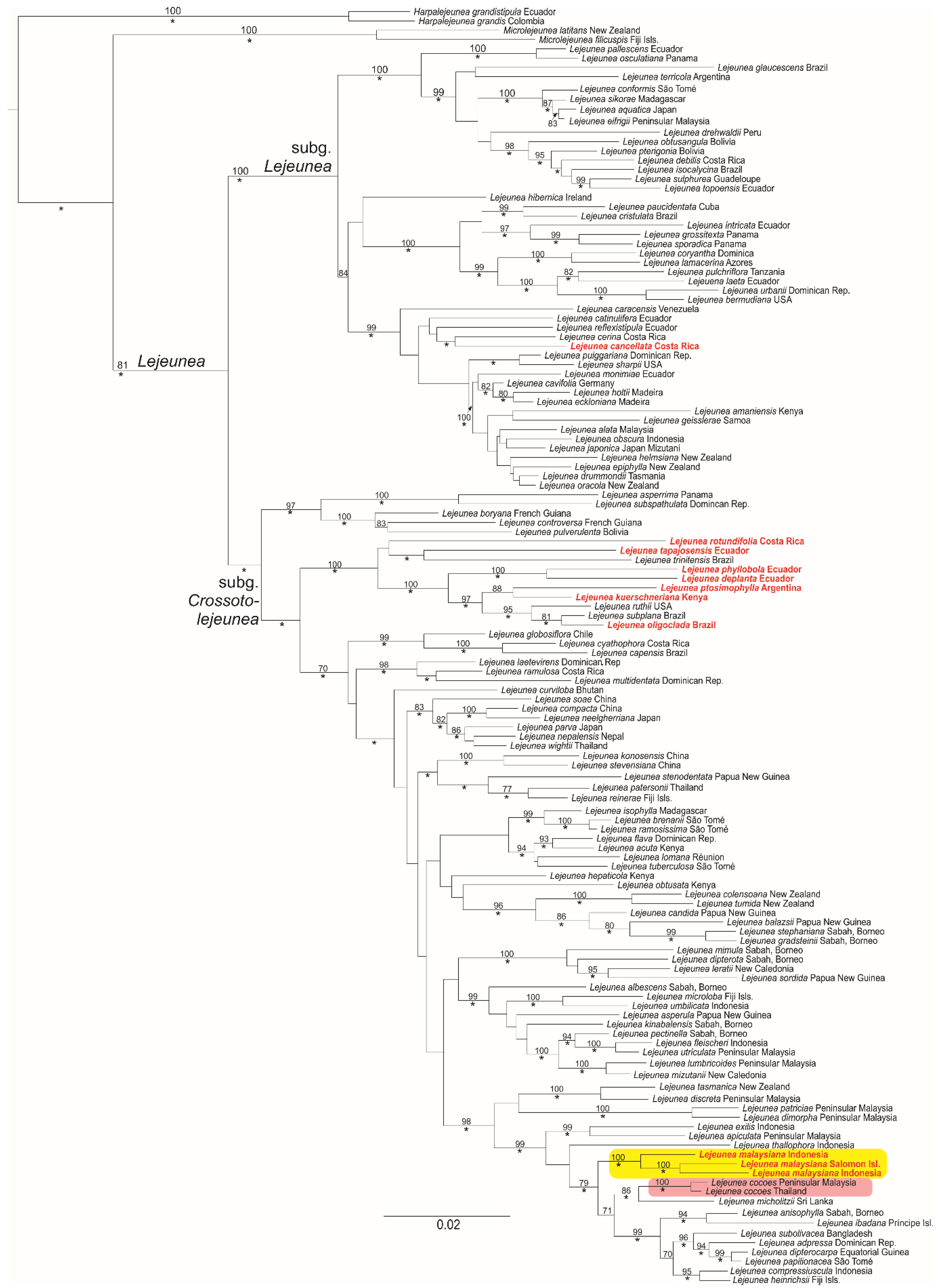
2.2. Molecular Phylogeny
2.3. Taxonomic Description
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Morphological Investigation
4.2. DNA Extraction, PCR Amplification, Sequencing and Alignment
4.3. Phylogenetic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, G.E.; Gradstein, S.R. Guide to the Genera of Liverworts and Hornworts of Malaysia; Hattori Botanical Laboratory: Tokyo, Japan, 2021; p. 234. [Google Scholar]
- Lee, G.E.; Pócs, T.; Gradstein, S.R.; Damanhuri, A.; Latiff, A. Abundant but neglected: Past and present of liverwort (Marchantiophyta) studies in Malaysia. Cryptog. Bryol. 2018, 93, 89–91. [Google Scholar] [CrossRef]
- Cheah, Y.H.; Yong, K.T. New records of Bazzania species (Marchantiophyta: Lepidoziaceae) in Peninsular Malaysia with identification key. Cryptog. Bryol. 2016, 37, 199–210. [Google Scholar] [CrossRef]
- Pesiu, E.; Sarimi, S.; Syafie, A.; Koid, C.W.; Ghazaly, M.; Norhazrina, N.; Pócs, T.; Lee, G.E. First floristic study on epiphyllous bryophytes of the state Terengganu, Peninsular Malaysia. Check List 2021, 17, 1404–1419. [Google Scholar] [CrossRef]
- Sarimi, M.S.; Pócs, T.; Lee, G.E. Data to the Malaysian liverwort flora, III: New Lejeuneaceae records from Sabah and Peninsular Malaysia. Cryptog. Bryol. 2021, 42, 249–267. [Google Scholar] [CrossRef]
- Katagiri, T.; Suleiman, M.; Deguchi, H. Taxonomic studies of the Trichocoleaceae in Southeast Asia. II. A new species of Eotrichocolea from Malaysia. Bryologist 2012, 115, 518–522. [Google Scholar] [CrossRef]
- Furuki, T.; Suleiman, M. Diplophyllum kinabaluense (Scapaniaceae, Marchantiophyta) sp. nov. from Mt. Kinabalu, Malaysian Borneo. J. Jpn. Bot. 2016, 91, 340–344. [Google Scholar]
- Sukkharak, P. Studies on the genus Mastigolejeunea (Marchantiophyta: Lejeuneaceae): Mastigolejeunea gradsteinii Sukkharak sp. nov. J. Bryol. 2014, 36, 56–60. [Google Scholar] [CrossRef]
- Lee, G.E.; Bechteler, J.; Pócs, T.; Schäfer-Verwimp, A.; Heinrichs, J. Molecular and morphological evidence for an intercontinental range of the liverwort Lejeunea pulchriflora (Marchantiophyta: Lejeuneaceae). Org. Divers. Evol. 2016, 16, 13–21. [Google Scholar] [CrossRef]
- Zhu, R.L.; Shu, L.; Mustapeng, A.M.A.; Suleiman, M. Thiersianthus (Marchantiophyta: Lejeuneaceae) a new genus from lowland rainforest in Borneo. Bryologist 2017, 120, 511–520. [Google Scholar] [CrossRef]
- Zhu, R.L.; Bi, X.F.; Shu, L. Mohamedia, a new genus of Lejeuneaceae (Marchantiophyta) from Oceania and tropical Asia. Bryologist 2019, 122, 84–97. [Google Scholar] [CrossRef]
- Lee, G.E. A systematic revision of the genus Lejeunea (Marchantiophyta: Lejeuneaceae) in Malaysia. Cryptog. Bryol. 2013, 34, 381–484. [Google Scholar] [CrossRef]
- Lee, G.E.; Gradstein, S.R.; Pesiu, E.; Norhazrina, N. An updated checklist of liverworts and hornworts of Malaysia. Phytokeys 2022, 199, 29–111. [Google Scholar] [CrossRef]
- Gradstein, S.R. The liverworts and hornworts of Colombia and Ecuador. Mem. N. Y. Bot. Gard. 2020, 121, 1–723. [Google Scholar]
- Lee, G.E.; Gradstein, S.R. Distribution and habitats of Lejeunea species from Malaysia. Pol. Bot. J. 2013, 58, 59–69. [Google Scholar]
- Lee, G.E.; Condamine, F.L.; Bechteler, J.; Pérez-Escobar, O.A.; Scheben, A.; Schäfer-Verwimp, A.; Pócs, T.; Heinrichs, J. An ancient tropical origin, dispersal via land bridges and Miocene diversification explain the subcosmopolitan disjunctions of the liverwort genus Lejeunea. Sci. Rep. 2020, 10, 14123. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWRs) Threatened and Endemic to Italy: Urgent Actions for Protection and Use. Biology 2022, 11, 193. [Google Scholar] [CrossRef]
- Renner, M.A.M.; Brown, E.A.; Wardle, G.M. The Lejeunea tumida species group is positively polyphyletic (Lejeuneaceae: Jungermanniopsida). Austral. Syst. Bot. 2011, 24, 10–18. [Google Scholar] [CrossRef]
- Lee, G.E.; Bechteler, J.; Pócs, T.; Schäfer-Verwimp, A. Molecular and morphological evidence for a new species of liverwort, Lejeunea heinrichsii (Marchantiophyta: Lejeuneaceae) from Taveuni, Fiji. J. Syst. Evol. 2019, 57, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Bastos, C.J.P.; Gradstein, S.R. The genus Lejeunea Lib. (Lejeuneaceae, Marchantiophyta) in Brazil. Phytotaxa 2020, 453, 55–107. [Google Scholar] [CrossRef]
- Pócs, T.; Piippo, S.; Mizutani, M. Bryophyte flora of the Huon Peninsular, Papua New Guinea. LVI. Preliminary contributions on Lejeuneaceae (Hepaticae) 2. Ann. Bot. Fenn. 1995, 32, 259–268. [Google Scholar]
- Reiner-Drehwald, M.E.; Goda, A. Revision of the genus Crossotolejeunea (Lejeuneaceae: Hepaticae). J. Hattori Bot. Lab. 2000, 89, 1–54. [Google Scholar]
- Zhu, R.L.; So, M.L. Epiphyllous liverworts of China. Beih. Nova Hedwigia. 2001, 121, 1–418. [Google Scholar]
- Zhu, R.L.; Grolle, R. Taxonomy and distribution of Lejeunea exilis (Lejeuneaceae, Hepaticae). Ann. Bot. Fenn. 2003, 40, 101–106. [Google Scholar]
- Lee, G.E.; Pócs, T. Additions to the distribution of some Palaeotropic Lejeunea (Marchantiophyta) species, with the description of Lejeunea konratii sp. nov. from Fiji. Phytotaxa 2018, 349, 31–38. [Google Scholar] [CrossRef]
- Bastos, C.J.P.; Reiner-Drehwald, M.E.; Schäfer-Verwimp, A. A new species of the genus Lejeunea Lib. (Marchantiophyta, Lejeuneaceae) from Brazil. Phytotaxa 2017, 326, 71–76. [Google Scholar] [CrossRef]
- Pócs, T. On some new or less known Lejeunea (Lejeuneaceae, Jungermanniopsida) species in tropical Africa. East African Bryophytes, XXVII. Beih. Nova Hedwigia. 2010, 138, 99–116. [Google Scholar]
- Mizutani, M. Lejeunea from the Himalayan Region. J. Hattori Bot. Lab. 1971, 34, 445–457. [Google Scholar]
- Zhu, R.L.; So, M.L. Additions of Lejeuneaceae taxa to the hepatic flora of Yunnan, China. Ann. Bot. Fenn. 1999, 36, 219–229. [Google Scholar]
- Reiner-Drehwald, M.E.; Schäfer-Verwimp, A. On Inflatolejeunea, Lejeunea species with eplicate perianths and Lejeunea talamancensis sp. nov. from Costa Rica (Lejeuneaceae). Nova Hedwigia. 2008, 87, 387–420. [Google Scholar] [CrossRef]
- Bechteler, J.; Lee, G.E.; Schäfer-Verwimp, A.; Pócs, T.; Peralta, D.F.; Renner, M.A.M.; Schneider, H.; Heinrichs, J. Toward a monophyletic classification of Lejeuneaceae IV: Reinstatement of Allorgella, transfer of Microlejeunea aphanella to Vitalianthus and refinements of the subtribal classification. Pl. Syst. Evol. 2016, 302, 187–201. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Borsch, T.; Quandt, D. Mutational dynamics and phylogenetic utility of noncoding chloroplast DNA. Pl. Syst. Evol. 2009, 282, 169–199. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Daribba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Mason-Gamer, R.J.; Kellogg, E.A. Testing for phylogenetic conflict among molecular data sets in the tribe Triticeae (Gramineae). Syst. Biol. 1996, 45, 524–545. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Larget, B.; Simon, D.L. Markov chain Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, G.E.; Bechteler, J.; Pócs, T.; Schäfer-Verwimp, A.; Tang, H.Y.; Chia, P.W. Integrative Taxonomy Reveals a New Species of the Genus Lejeunea (Marchantiophya: Lejeuneaceae) from Peninsular Malaysia. Plants 2022, 11, 1642. https://doi.org/10.3390/plants11131642
Lee GE, Bechteler J, Pócs T, Schäfer-Verwimp A, Tang HY, Chia PW. Integrative Taxonomy Reveals a New Species of the Genus Lejeunea (Marchantiophya: Lejeuneaceae) from Peninsular Malaysia. Plants. 2022; 11(13):1642. https://doi.org/10.3390/plants11131642
Chicago/Turabian StyleLee, Gaik Ee, Julia Bechteler, Tamás Pócs, Alfons Schäfer-Verwimp, Hung Yung Tang, and Poh Wai Chia. 2022. "Integrative Taxonomy Reveals a New Species of the Genus Lejeunea (Marchantiophya: Lejeuneaceae) from Peninsular Malaysia" Plants 11, no. 13: 1642. https://doi.org/10.3390/plants11131642