
Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review

 ,
, 
 ,
,  ,
,  and
and .jpg)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
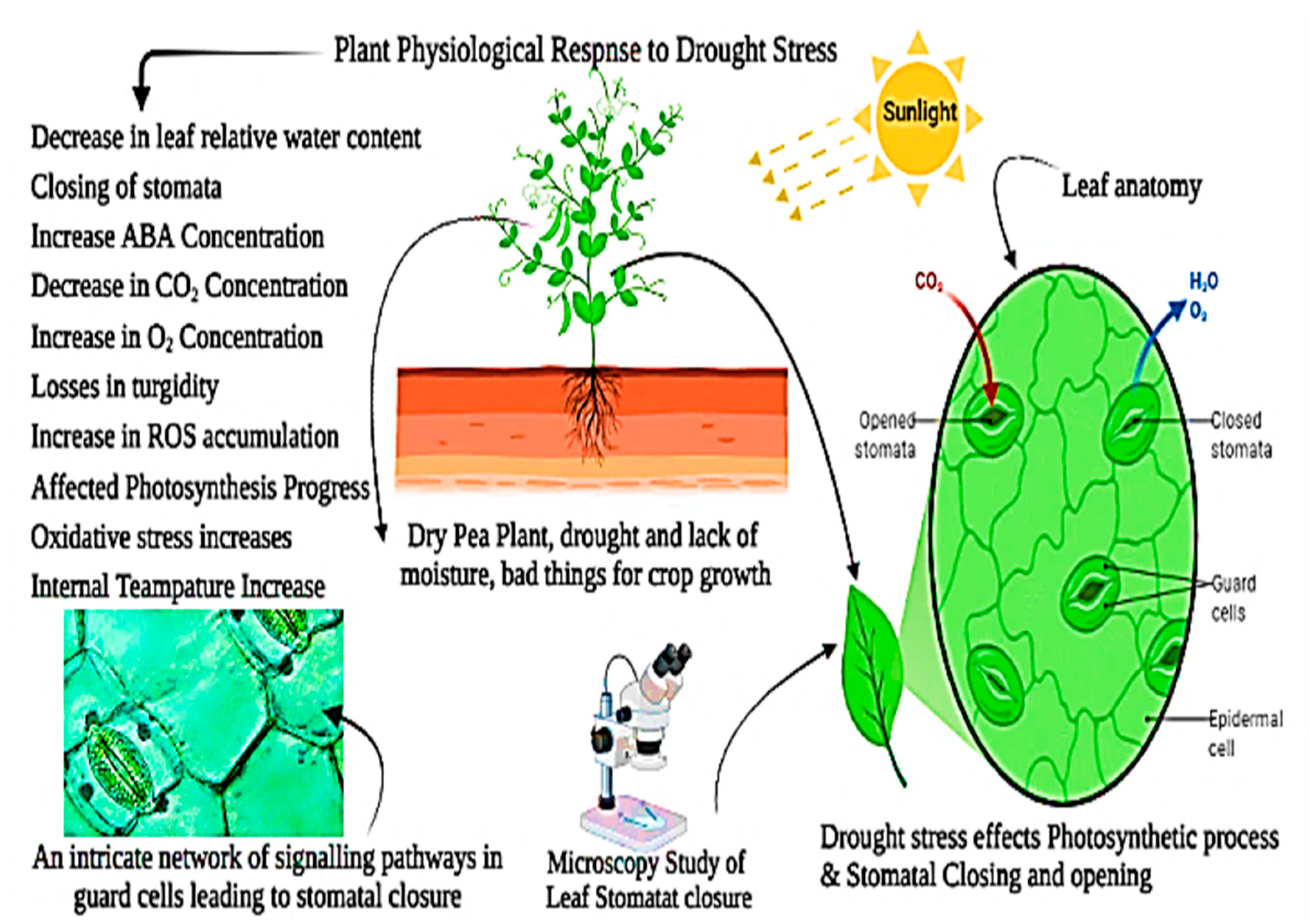{kind=link}
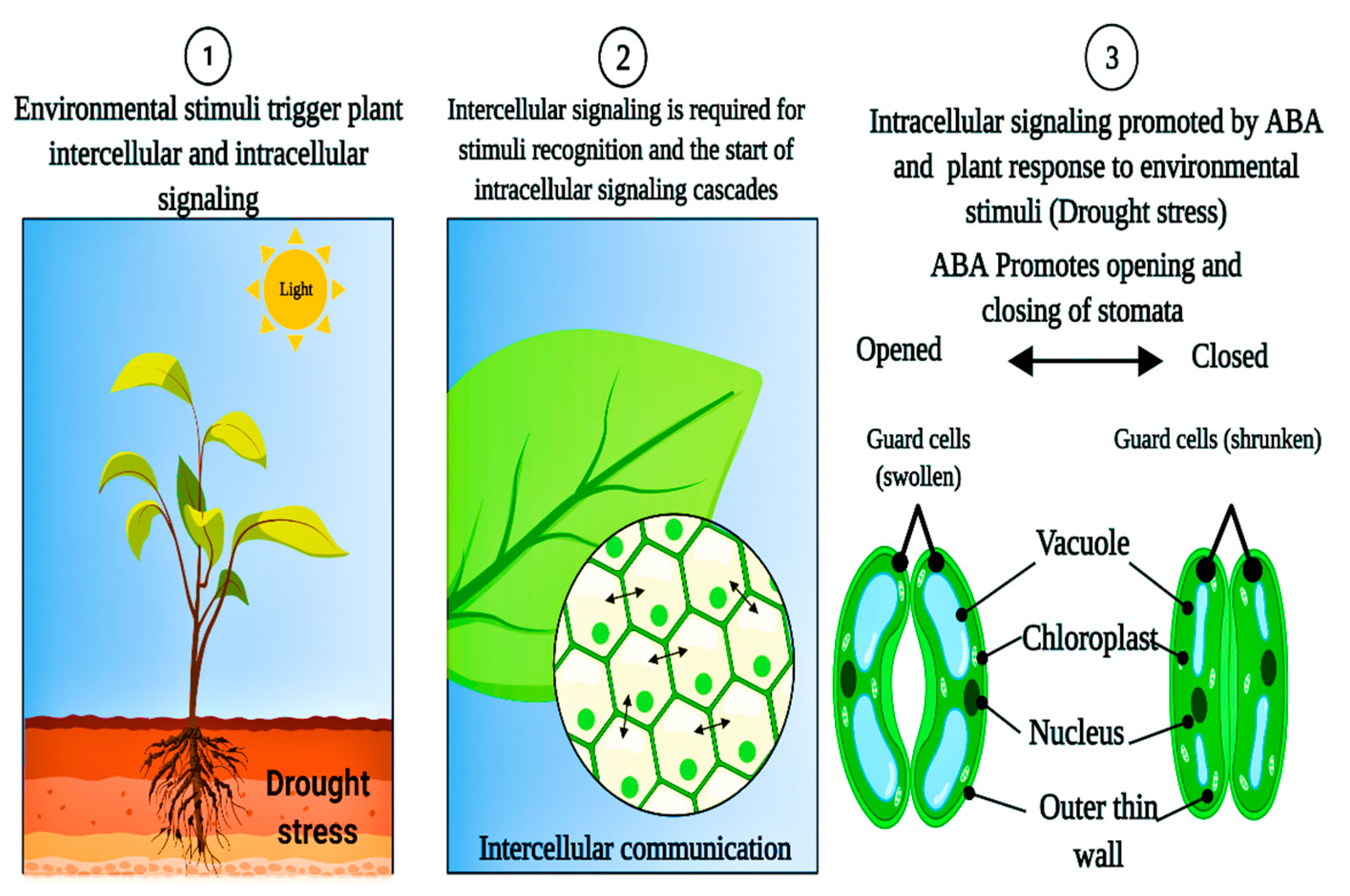{kind=link}
Abstract
:1. Introduction
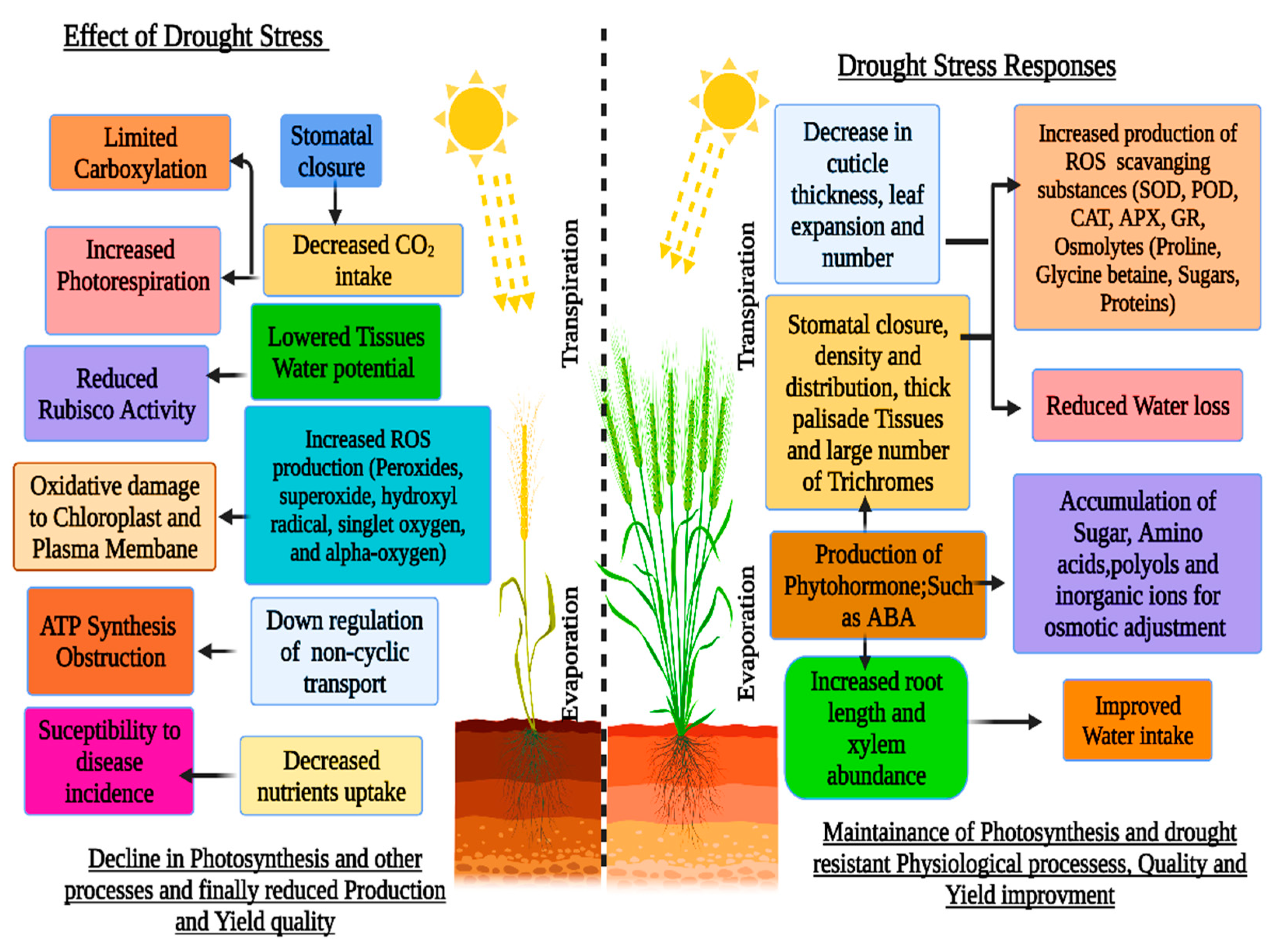
Drought Stress
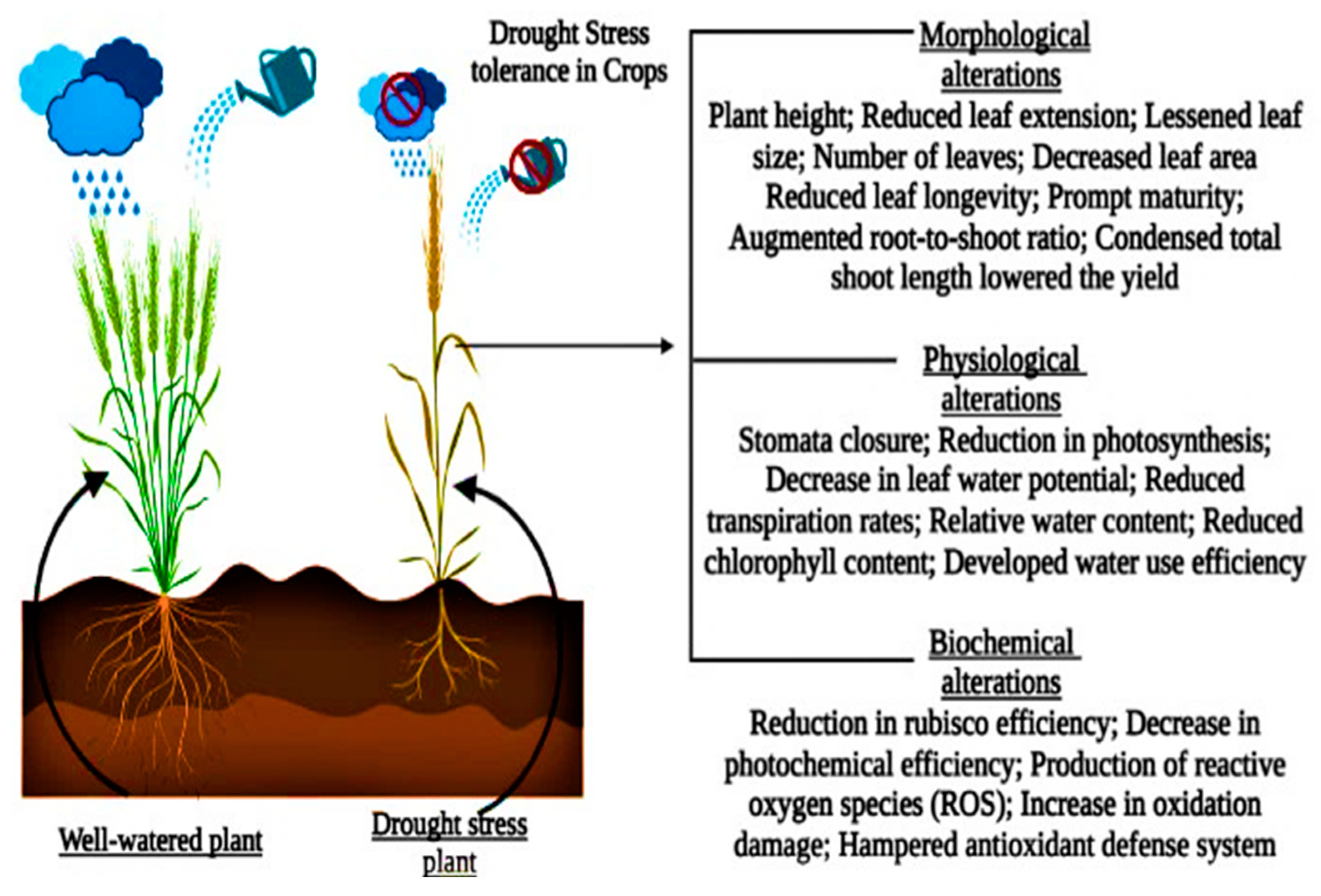
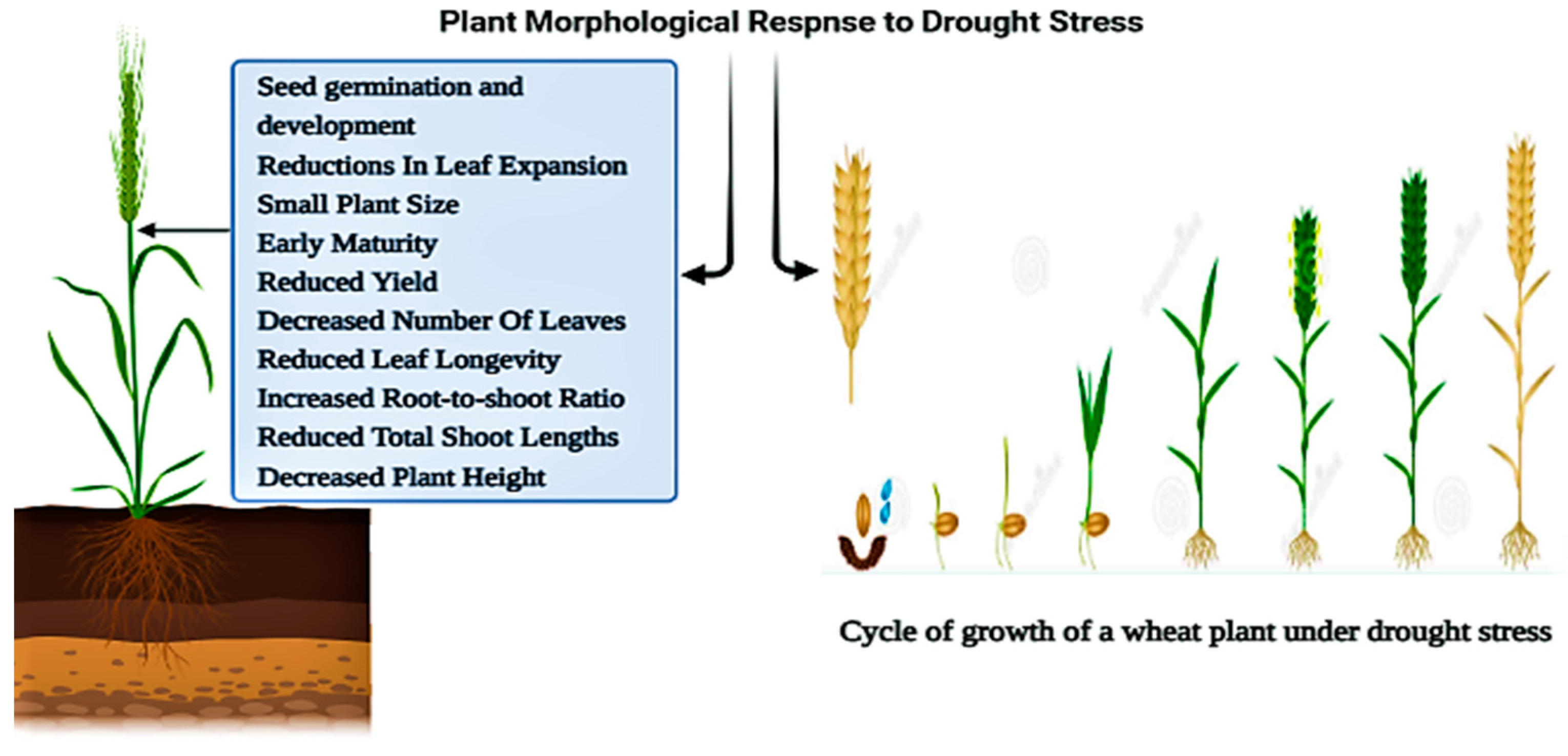
2. Drought-Induced Changes in Plant Morphology
2.1. Early Seed Germination and Flowering
2.2. Plant Morphological Characteristics of Leaves under Drought Stress
2.3. Plants Shoot Morphology and Architecture under Drought Stress
2.4. Plant Root Morphology and Architecture under Drought Stress
2.5. Yield
3. Physiological and Biochemical Responses under Drought Stress
3.1. Leaf Relative Water Content (RWC)
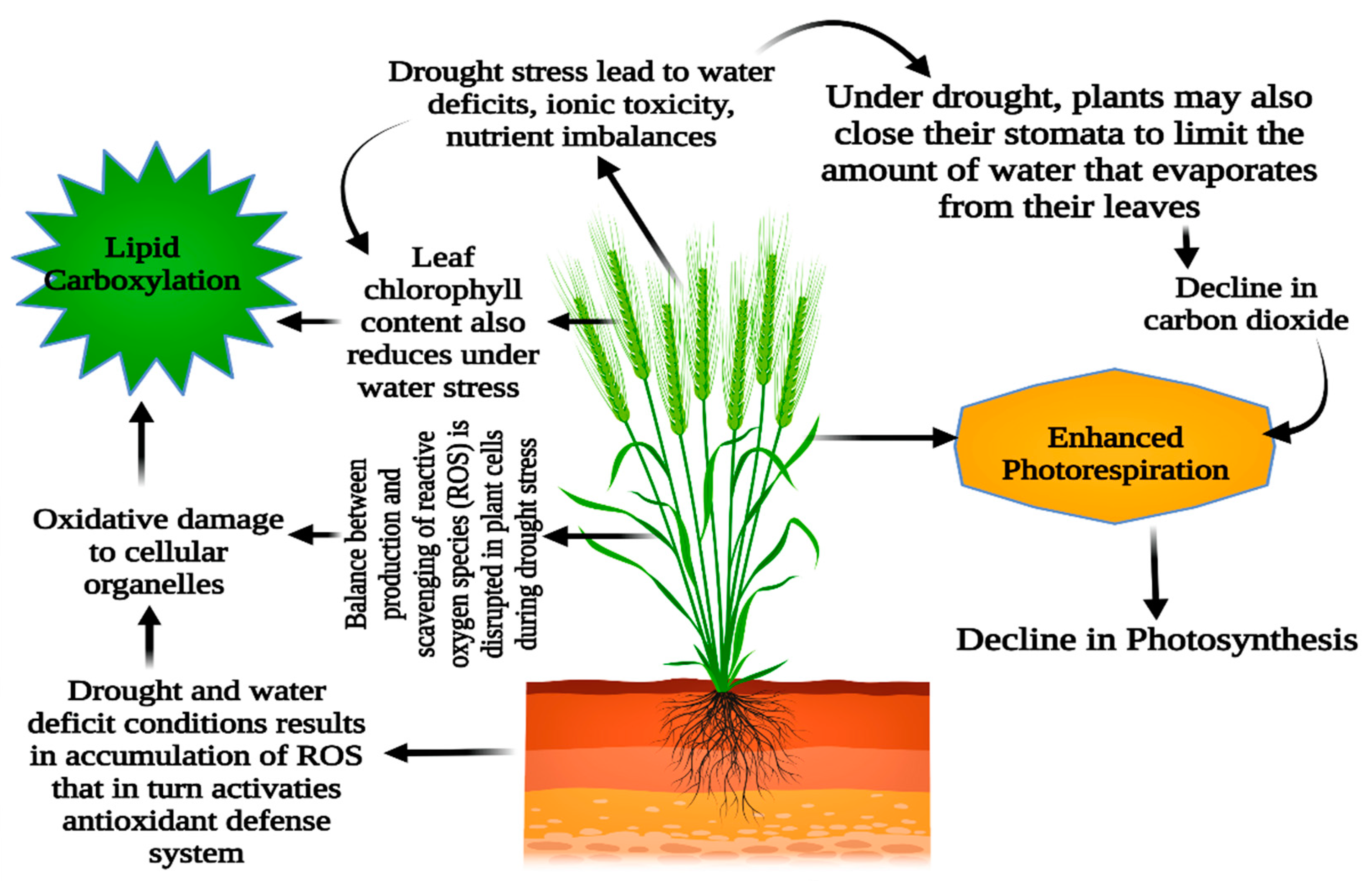
3.2. Effect of Drought Stress Conditions on Photosynthesis and Stomatal Aperture
3.3. Carotenoids
3.4. Cell Size, Cell Membrane Stability, and Respiration
4. Biochemical Responses under Drought Stress Conditions
4.1. Reactive Oxygen Species (ROS)
4.2. Total Soluble Phenolic, Antioxidatant Enzymatic, and Osmolyte Regulation under Drought Stress Conditions
5. Improvement of Drought Tolerance Using Molecular Tools
6. Phytohormonal Modulation under Drought Stress
6.1. Salicylic Acid
6.2. Cytokinin and Auxin
6.3. Gibberellins
6.4. Abscise Acid
6.5. Ethylene
6.6. Jasmonates Acid (JAs)
7. Conclusions
8. Concluding Remarks and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amna; Ali, B.; Azeem, M.A.; Qayyum, A.; Mustafa, G.; Ahmad, M.A.; Javed, M.T.; Chaudhary, H.J. Bio-Fabricated Silver Nanoparticles: A Sustainable Approach for Augmentation of Plant Growth and Pathogen Control. In Sustainable Agriculture Reviews 53; Springer: Berlin/Heidelberg, Germany, 2021; pp. 345–371. [Google Scholar]
- Zainab, N.; Amna; Khan, A.A.; Azeem, M.A.; Ali, B.; Wang, T.; Shi, F.; Alghanem, S.M.; Hussain Munis, M.F.; Hashem, M.; et al. PGPR-Mediated Plant Growth Attributes and Metal Extraction Ability of Sesbania sesban L. in Industrially Contaminated Soils. Agronomy 2021, 11, 1820. [Google Scholar] [CrossRef]
- Hussain, S.Q.; Rasheed, M.; Saleem, M.H.; Ahmed, Z.I.; Hafeez, A.; Jilani, G.; Alamri, S.; Hashem, M.; Ali, S. Salt tolerance in maize with melatonin priming to achieve sustainability in yield on salt affected soils. Pak. J. Bot. 2022, 55, 1. [Google Scholar] [CrossRef]
- Yahaya, M.A.; Shimelis, H. Drought Stress in Sorghum: Mitigation Strategies, Breeding Methods and Technologies—A Review. J. Agron. Crop. Sci. 2022, 208, 127–142. [Google Scholar] [CrossRef]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef]
- McDowell, N.G.; Sapes, G.; Pivovaroff, A.; Adams, H.D.; Allen, C.D.; Anderegg, W.R.L.; Arend, M.; Breshears, D.D.; Brodribb, T.; Choat, B.; et al. Mechanisms of Woody-Plant Mortality under Rising Drought, CO2 and Vapour Pressure Deficit. Nat. Rev. Earth Environ. 2022, 3, 294–308. [Google Scholar] [CrossRef]
- Pepe, M.; Crescente, M.F.; Varone, L. Effect of Water Stress on Physiological and Morphological Leaf Traits: A Comparison among the Three Widely-Spread Invasive Alien Species Ailanthus Altissima, Phytolacca Americana, and Robinia Pseudoacacia. Plants 2022, 11, 899. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology (Basel) 2022, 11, 155. [Google Scholar] [CrossRef]
- Sofy, M.R.; Aboseidah, A.A.; Heneidak, S.A.; Ahmed, H.R. ACC Deaminase Containing Endophytic Bacteria Ameliorate Salt Stress in Pisum Sativum through Reduced Oxidative Damage and Induction of Antioxidative Defense Systems. Environ. Sci. Pollut. Res. 2021, 28, 40971–40991. [Google Scholar] [CrossRef]
- Perveen, S.; Hussain, S.A. Methionine-Induced Changes in Growth, Glycinebetaine, Ascorbic Acid, Total Soluble Proteins and Anthocyanin Contents of Two Zea mays L. Varieties under Salt Stress. J. Anim. Plant Sci. 2020, 31, 131–142. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Sumaira; Hafeez, A.; Afridi, M.S.; Khan, S.; Zaib-Un-Nisa; Ullah, I.; Amaral Júnior, A.T.; et al. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and Its Actions during the Drought Stress in Plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Wang, X.; Saleem, M.H.; Azeem, M.A.; Afridi, M.S.; Nadeem, M.; Ghazal, M.; Batool, T.; Qayyum, A.; Alatawi, A.; et al. Bacillus mycoides PM35 Reinforces Photosynthetic Efficiency, Antioxidant Defense, Expression of Stress-Responsive Genes, and Ameliorates the Effects of Salinity Stress in Maize. Life 2022, 12, 219. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Nie, L.; Chen, Y.Y.; Wu, C.; Xiong, D.; Saud, S.; Hongyan, L.; Cui, K.; Huang, J.; Bogati, K.; et al. The Solanum Melongena COP1LIKE Manipulates Fruit Ripening and Flowering Time in Tomato (Solanum lycopersicum). Front. Plant Sci. 2021, 10, 369–382. [Google Scholar] [CrossRef]
- Chai, Q.; Gan, Y.; Zhao, C.; Xu, H.L.; Waskom, R.M.; Niu, Y.; Siddique, K.H.M. Regulated Deficit Irrigation for Crop Production under Drought Stress. A Review. Agron. Sustain. Dev. 2016, 36, 3. [Google Scholar] [CrossRef] [Green Version]
- Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Fahad, S.; Datta, R.; Islam, A.K.M.A. Drought Stress in Grain Legumes: Effects, Tolerance Mechanisms and Management. Agronomy 2021, 11, 2374. [Google Scholar] [CrossRef]
- Alam, H.; Khattak, J.Z.; Ksiksi, T.S.; Saleem, M.H.; Fahad, S.; Sohail, H.; Ali, Q.; Zamin, M.; El-Esawi, M.A.; Saud, S. Negative impact of long-term exposure of salinity and drought stress on native Tetraena mandavillei L. Physiol. Plant. 2021, 172, 1336–1351. [Google Scholar] [CrossRef]
- Ghafar, M.A.; Akram, N.A.; Saleem, M.H.; Wang, J.; Wijaya, L.; Alyemeni, M.N. Ecotypic Morphological and Physio-Biochemical Responses of Two Differentially Adapted Forage Grasses, Cenchrus ciliaris L. and Cyperus arenarius Retz. to Drought Stress. Sustainability 2021, 13, 8069. [Google Scholar] [CrossRef]
- Azeem, M.; Haider, M.Z.; Javed, S.; Saleem, M.H.; Alatawi, A. Drought Stress Amelioration in Maize (Zea mays L.) by Inoculation of Bacillus spp. Strains under Sterile Soil Conditions. Agriculture 2022, 12, 50. [Google Scholar] [CrossRef]
- Ma, Y.; Vosátka, M.; Freitas, H. Editorial: Beneficial Microbes Alleviate Climatic Stresses in Plants. Front. Plant Sci. 2019, 10, 595. [Google Scholar] [CrossRef]
- Grzesiak, M.T.; Hordyńska, N.; Maksymowicz, A.; Grzesiak, S.; Szechyńska-Hebda, M. Variation among Spring Wheat (Triticum aestivum L.) Genotypes in Response to the Drought Stress. Ii—Root System Structure. Plants 2019, 8, 584. [Google Scholar] [CrossRef] [Green Version]
- Kamal, N.M.; Gorafi, Y.S.A.; Abdelrahman, M.; Abdellatef, E.; Tsujimoto, H. Stay-Green Trait: A Prospective Approach for Yield Potential, and Drought and Heat Stress Adaptation in Globally Important Cereals. Int. J. Mol. Sci. 2019, 20, 5837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and Biochemical Changes during Drought and Recovery Periods at Tillering and Jointing Stages in Wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Boudiar, R.; Casas, A.M.; Gioia, T.; Fiorani, F.; Nagel, K.A.; Igartua, E. Effects of Low Water Availability on Root Placement and Shoot Development in Landraces and Modern Barley Cultivars. Agronomy 2020, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, S.; Khatoon, Z.; Amna; Ahmad, I.; Muneer, M.A.; Kamran, M.A.; Ali, J.; Ali, B.; Chaudhary, H.J.; Munis, M.F.H. Bacillus sp. PM31 harboring various plant growth-promoting activities regulates Fusarium dry rot and wilt tolerance in potato. Arch. Agron. Soil Sci. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Singh, B.; Norvell, E.; Wijewardana, C.; Wallace, T.; Chastain, D.; Reddy, K.R. Assessing Morphological Characteristics of Elite Cotton Lines from Different Breeding Programmes for Low Temperature and Drought Tolerance. J. Agron. Crop. Sci. 2018, 204, 467–476. [Google Scholar] [CrossRef]
- Queiroz, M.S.; Oliveira, C.E.S.; Steiner, F.; Zuffo, A.M.; Zoz, T.; Vendruscolo, E.P.; Silva, M.V.; Mello, B.F.F.R.; Cabral, R.C.; Menis, F.T. Drought Stresses on Seed Germination and Early Growth of Maize and Sorghum. J. Agric. Sci. 2019, 11, 310. [Google Scholar] [CrossRef]
- Wagaw, K. Isolation and Screening of ACC Deaminase-Producing Microbes for Drought Stress Management in Crops. Acad. Res. J. Agri. Sci. Res. 2019, 7, 87–99. [Google Scholar] [CrossRef]
- Islam, M.M.; Kayesh, E.; Zaman, E.; Urmi, T.A.; Haque, M.M. Evaluation of Rice (Oryza sativa L.) Genotypes for Drought Tolerance at Germination and Early Seedling Stage. Agriculturists 2018, 16, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Al-Quraan, N.A.; Al-Ajlouni, Z.I.; Qawasma, N.F. Physiological and Biochemical Characterization of the Gaba Shunt Pathway in Pea (Pisum sativum L.) Seedlings under Drought Stress. Horticulturae 2021, 7, 125. [Google Scholar] [CrossRef]
- Sabagh, A.E.L.; Hossain, A.; Barutçular, C.; Iqbal, M.A.; Islam, M.S.; Fahad, S.; Sytar, O.; Çiğ, F.; Meena, R.S.; Erman, M. Consequences of Salinity Stress on the Quality of Crops and Its Mitigation Strategies for Sustainable Crop Production: An Outlook of Arid and Semi-Arid Regions. In Environment, Climate, Plant and Vegetation Growth; Springer: Berlin/Heidelberg, Germany, 2020; pp. 503–533. [Google Scholar]
- Liang, Y.; Tabien, R.E.; Tarpley, L.; Mohammed, A.R.; Septiningsih, E.M. Transcriptome Profiling of Two Rice Genotypes under Mild Field Drought Stress during Grain-Filling Stage. AoB Plants 2021, 13, plab043. [Google Scholar] [CrossRef]
- Damalas, C.A. Improving Drought Tolerance in Sweet Basil (Ocimum basilicum) with Salicylic Acid. Sci. Hortic. (Amsterdam) 2019, 246, 360–365. [Google Scholar] [CrossRef]
- Ahmad, S.; Kamran, M.; Ding, R.; Meng, X.; Wang, H.; Ahmad, I.; Fahad, S.; Han, Q. Exogenous Melatonin Confers Drought Stress by Promoting Plant Growth, Photosynthetic Capacity and Antioxidant Defense System of Maize Seedlings. PeerJ 2019, 2019, e7793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, C.; Bagavathiannan, M.; Wang, H.; Sharpe, S.M.; Meng, W.; Yu, J. Osmopriming with Polyethylene Glycol (Peg) for Abiotic Stress Tolerance in Germinating Crop Seeds: A Review. Agronomy 2021, 11, 2194. [Google Scholar] [CrossRef]
- Liang, B.; Gao, T.; Zhao, Q.; Ma, C.; Chen, Q.; Wei, Z.; Li, C.; Li, C.; Ma, F. Effects of Exogenous Dopamine on the Uptake, Transport, and Resorption of Apple Ionome under Moderate Drought. Front. Plant Sci. 2018, 9, 755. [Google Scholar] [CrossRef]
- Elnaggar, A.; El-Keblawy, A.; Mosa, K.A.; Soliman, S. Drought Tolerance during Germination Depends on Light and Temperature of Incubation in Salsola Imbricata, a Desert Shrub of Arabian Deserts. Flora Morphol. Distrib. Funct. Ecol. Plants 2018, 249, 156–163. [Google Scholar] [CrossRef]
- Paponov, M.; Kechasov, D.; Lacek, J.; Verheul, M.J.; Paponov, I.A. Supplemental Light-Emitting Diode Inter-Lighting Increases Tomato Fruit Growth Through Enhanced Photosynthetic Light Use Efficiency and Modulated Root Activity. Front. Plant Sci. 2020, 10, 1656. [Google Scholar] [CrossRef]
- Kumar, S.; Islam, A.R.M.T.; Islam, H.M.T.; Hasanuzzaman, M.; Ongoma, V.; Khan, R.; Mallick, J. Water Resources Pollution Associated with Risks of Heavy Metals from Vatukoula Goldmine Region, Fiji. J. Environ. Manag. 2021, 293, 112868. [Google Scholar] [CrossRef]
- Naz, S.; Perveen, S. Response of Wheat (Triticum aestivum L. Var. Galaxy-2013) to Pre-Sowing Seed Treatment with Thiourea under Drought Stress. Pakistan J. Bot. 2021, 53, 1209–1217. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.; Mi, N.; Ming, H.; Zhang, S.; Zhang, H.; Zhao, X. Maize (Zea mays L.) Physiological Responses to Drought and Rewatering, and the Associations with Water Stress Degree. Agric. Water Manag. 2020, 241, 106379. [Google Scholar] [CrossRef]
- Widuri, L.I.; Lakitan, B.; Sodikin, E.; Hasmeda, M.; Meihana, M.; Kartika, K.; Siaga, E. Shoot and Root Growth in Common Bean (Phaseolus vulgaris L.) Exposed to Gradual Drought Stress. Agrivita 2018, 40, 442–452. [Google Scholar] [CrossRef]
- Tůmová, L.; Tarkowská, D.; Řřová, K.; Marková, H.; Kočová, M.; Rothová, O.; čečetka, P.; Holá, D. Drought-Tolerant and Drought-Sensitive Genotypes of Maize (Zea mays L.) Differ in Contents of Endogenous Brassinosteroids and Their Drought-Induced Changes. PLoS ONE 2018, 13, e0197870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocchini, M.; D’Amato, R.; Ciancaleoni, S.; Fontanella, M.C.; Palmerini, C.A.; Beone, G.M.; Onofri, A.; Negri, V.; Marconi, G.; Albertini, E.; et al. Soil Selenium (Se) Biofortification Changes the Physiological, Biochemical and Epigenetic Responses to Water Stress in Zea mays L. by Inducing a Higher Drought Tolerance. Front. Plant Sci. 2018, 9, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begum, N.; Ahanger, M.A.; Su, Y.; Lei, Y.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L. Improved Drought Tolerance by AMF Inoculation in Maize (Zea mays) Involves Physiological and Biochemical Implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A. Effect of Soil Water Deficit on Growth and Development of Plants: A Review. Soil Water Deficit Physiol. Issues Plants 2021, 2021, 393–488. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, B. Arbuscular Mycorrhiza Induced Putrescine Degradation into γ-Aminobutyric Acid, Malic Acid Accumulation, and Improvement of Nitrogen Assimilation in Roots of Water-Stressed Maize Plants. Mycorrhiza 2020, 30, 329–339. [Google Scholar] [CrossRef]
- Valliere, J.M.; Zhang, J.; Sharifi, M.R.; Rundel, P.W. Can We Condition Native Plants to Increase Drought Tolerance and Improve Restoration Success? Ecol. Appl. 2019, 29, e01863. [Google Scholar] [CrossRef]
- Sharma, M.; Delta, A.K.; Kaushik, P. Glomus Mosseae and Pseudomonas Fluorescens Application Sustains Yield and Promote Tolerance to Water Stress in Helianthus annuus L. Stresses 2021, 1, 305–316. [Google Scholar] [CrossRef]
- Malinowska, M.; Donnison, I.; Robson, P. Morphological and Physiological Traits That Explain Yield Response to Drought Stress in Miscanthus. Agronomy 2020, 10, 1194. [Google Scholar] [CrossRef]
- Istanbuli, T.; Baum, M.; Touchan, H.; Hamwieh, A. Evaluation of Morpho-Physiological Traits under Drought Stress Conditions in Barley (Hordeum vulgare L.). Photosynthetica 2020, 58, 1059–1067. [Google Scholar] [CrossRef]
- Nofouzi, F. Evaluation of Seed Yield of Durum Wheat (Triticum durum) under Drought Stress and Determining Correlation among Some Yield Components Using Path Coefficient Analysis. UNED Res. J. 2018, 10, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Kalagare, V.S.; Ganesan, N.M.; Iyanar, K.; Chitdeshwari, T.; Chandrasekhar, C.N. Strategy of Multiple Selection Indices for Discrimination of Potential Genotypes and Associated Traits for Yield Improvement in Pearl Millet [Pennisetum glaucum (L.) R.Br.]. Electron. J. Plant Breed. 2021, 12, 895–906. [Google Scholar] [CrossRef]
- Ullah, A.; Farooq, M. The Challenge of Drought Stress for Grain Legumes and Options for Improvement. Arch. Agron. Soil Sci. 2021, 2021, 1–18. [Google Scholar] [CrossRef]
- Wellstein, C.; Poschlod, P.; Gohlke, A.; Chelli, S.; Campetella, G.; Rosbakh, S.; Canullo, R.; Kreyling, J.; Jentsch, A.; Beierkuhnlein, C. Effects of Extreme Drought on Specific Leaf Area of Grassland Species: A Meta-Analysis of Experimental Studies in Temperate and Sub-Mediterranean Systems. Glob. Chang. Biol. 2017, 23, 2473–2481. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.; Waraich, E.A.; Akhtar, S.; Anjum, S.; Ahmad, T.; Mahboob, W.; Hafeez, O.B.A.; Tapera, T.; Labuschagne, M.; Rizwan, M. Physiological Responses of Wheat to Drought Stress and Its Mitigation Approaches. Acta Physiol. Plant. 2018, 40, 80. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant Responses of Wheat Plants under Stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate Change Regulated Abiotic Stress Mechanisms in Plants: A Comprehensive Review. Plant Cell Rep. 2022, 41, 1–31. [Google Scholar] [CrossRef]
- Kijowska-Oberc, J.; Staszak, A.M.; Kamiński, J.; Ratajczak, E. Adaptation of Forest Trees to Rapidly Changing Climate. Forests 2020, 11, 123. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Abdel-Hamid, A.M.E.; Yessoufou, K.; Al-Mana, F.A.; El-Ansary, D.O.; Mahmoud, E.A.; Al-Yafrasi, M.A. Physiological and Molecular Characterization of Water-Stressed Chrysanthemum under Robinin and Chitosan Treatment. Acta Physiol. Plant. 2020, 42, 31. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of Plant Responses to Water Stress and Related Genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Correia, M.J.; Rodrigues, M.L.; Ferreira, M.I.; Pereira, J.S. Diurnal Change in the Relationship between Stomatal Conductance and Abscisic Acid in the Xylem Sap of Field-Grown Peach Trees. J. Exp. Bot. 1997, 48, 1727–1736. [Google Scholar] [CrossRef]
- Kim, K.H.; Kabir, E.; Jahan, S.A. Exposure to Pesticides and the Associated Human Health Effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Badr, A.; Brüggemann, W. Comparative Analysis of Drought Stress Response of Maize Genotypes Using Chlorophyll Fluorescence Measurements and Leaf Relative Water Content. Photosynthetica 2020, 58, 638–645. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Du, Y.; Wang, J.; Wu, A.; Qiao, S.; Xu, B.; Zhang, S.; Siddique, K.H.M.; Chen, Y. Moderate Drought Stress Affected Root Growth and Grain Yield in Old, Modern and Newly Released Cultivars of Winter Wheat. Front. Plant Sci. 2017, 8, 672. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Khalid, M.F.; Saqib, M.; Ahmad, S.; Zafar, W.; Rao, M.J.; Morillon, R.; Anjum, M.A. Drought Tolerance in Citrus Rootstocks Is Associated with Better Antioxidant Defense Mechanism. Acta Physiol. Plant. 2018, 40, 135. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Alghory, A.; Yazar, A. Evaluation of Crop Water Stress Index and Leaf Water Potential for Deficit Irrigation Management of Sprinkler-Irrigated Wheat. Irrig. Sci. 2019, 37, 61–77. [Google Scholar] [CrossRef]
- Yan, F.; Sun, Y.; Xu, H.; Yin, Y.; Wang, H.; Wang, C.; Guo, C.; Yang, Z.; Sun, Y.; Ma, J. Effects of Wheat Straw Mulch Application and Nitrogen Management on Rice Root Growth, Dry Matter Accumulation and Rice Quality in Soils of Different Fertility. Paddy Water Environ. 2018, 16, 507–518. [Google Scholar] [CrossRef]
- Ashrafi, M.; Azimi-Moqadam, M.R.; Mohsenifard, E.; Shekari, F.; Jafary, H.; Moradi, P.; Pucci, M.; Abate, G.; Mastinu, A. Physiological and Molecular Aspects of Two Thymus Species Differently Sensitive to Drought Stress. BioTech 2022, 11, 8. [Google Scholar] [CrossRef]
- Duursma, R.A.; Blackman, C.J.; Lopéz, R.; Martin-StPaul, N.K.; Cochard, H.; Medlyn, B.E. On the Minimum Leaf Conductance: Its Role in Models of Plant Water Use, and Ecological and Environmental Controls. New Phytol. 2019, 221, 693–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misson, L.; Limousin, J.M.; Rodriguez, R.; Letts, M.G. Leaf Physiological Responses to Extreme Droughts in Mediterranean Quercus Ilex Forest. Plant Cell Environ. 2010, 33, 1898–1910. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Tian, S.B.; Di, Q.; Duan, S.H.; Dai, K. Effects of Exogenous Calcium on Mesophyll Cell Ultrastructure, Gas Exchange, and Photosystem II in Tobacco (Nicotiana tabacum Linn.) under Drought Stress. Photosynthetica 2018, 56, 1204–1211. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I. Optimising Photosynthesis for Environmental Fitness. Funct. Plant Biol. 2020, 47. [Google Scholar] [CrossRef]
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P.; Pintado, M.M.; Vasconcelos, M.W. Preserving the Nutritional Quality of Crop Plants under a Changing Climate: Importance and Strategies. Plant Soil 2019, 443, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Devi, M.J.; Bhatnagar-Mathur, P.; Sharma, K.K.; Serraj, R.; Anwar, S.Y.; Vadez, V. Relationships Between Transpiration Efficiency and Its Surrogate Traits in the Rd29A:DREB1A Transgenic Lines of Groundnut. J. Agron. Crop Sci. 2011, 197, 272–283. [Google Scholar] [CrossRef]
- Ferrara, A.; Lovelli, S.; Di Tommaso, T.; Perniola, M. Flowering, Growth and Fruit Setting in Greenhouse Bell Pepper under Water Stress. J. Agron. 2011, 10, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Tátrai, Z.A.; Sanoubar, R.; Pluhár, Z.; Mancarella, S.; Orsini, F.; Gianquinto, G. Morphological and Physiological Plant Responses to Drought Stress in Thymus Citriodorus. Int. J. Agron. 2016, 2016, 4165750. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Liu, J.; Liu, H.; Qiu, R.; Gao, Y.; Duan, A. Corrigendum: Role of Hydraulic Signal and ABA in Decrease of Leaf Stomatal and Mesophyll Conductance in Soil Drought-Stressed Tomato (Frontiers in Plant Science, (2021), 12, (653186), 10.3389/Fpls.2021.653186). Front. Plant Sci. 2021, 12, 711. [Google Scholar] [CrossRef]
- Ings, J.; Mur, L.A.J.; Robson, P.R.H.; Bosch, M. Physiological and Growth Responses to Water Deficit in the Bioenergy Crop Miscanthus x Giganteus. Front. Plant Sci. 2013, 4, 468. [Google Scholar] [CrossRef] [Green Version]
- Pourghasemian, N.; Moradi, R.; Naghizadeh, M.; Landberg, T. Mitigating Drought Stress in Sesame by Foliar Application of Salicylic Acid, Beeswax Waste and Licorice Extract. Agric. Water Manag. 2020, 231, 105997. [Google Scholar] [CrossRef]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. Int. J. Mol. Sci. 2020, 21, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alamri, S.A.; Alam, P.; Ashraf, M.; Ahmad, P. Potential of Exogenously Sourced Kinetin in Protecting Solanum lycopersicum from NaCl-Induced Oxidative Stress through up-Regulation of the Antioxidant System, Ascorbate-Glutathione Cycle and Glyoxalase System. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef]
- Uarrota, V.G.; Stefen, D.L.V.; Leolato, L.S.; Gindri, D.M.; Nerling, D. Revisiting Carotenoids and Their Role in Plant Stress Responses: From Biosynthesis to Plant Signaling Mechanisms during Stress. In Antioxidants and Antioxidant Enzymes in Higher Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 207–232. ISBN 9783319750880. [Google Scholar]
- Riaz, M.; Zia-Ul-Haq, M.; Dou, D. Chemistry of Carotenoids. In Carotenoids: Structure and Function in the Human Body; Springer: Berlin/Heidelberg, Germany, 2021; pp. 43–76. [Google Scholar]
- Islam, M.J.; Kim, J.W.; Begum, M.K.; Sohel, M.A.T.; Lim, Y.S. Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants 2020, 9, 1511. [Google Scholar] [CrossRef] [PubMed]
- Hashmat, S.; Shahid, M.; Tanwir, K.; Abbas, S.; Ali, Q.; Niazi, N.K.; Akram, M.S.; Saleem, M.H.; Javed, M.T. Elucidating Distinct Oxidative Stress Management, Nutrient Acquisition and Yield Responses of Pisum sativum L. Fertigated with Diluted and Treated Wastewater. Agric. Water Manag. 2021, 247, 106720. [Google Scholar] [CrossRef]
- Yang, Y.J.; Bi, M.H.; Nie, Z.F.; Jiang, H.; Liu, X.D.; Fang, X.W.; Brodribb, T.J. Evolution of Stomatal Closure to Optimize Water-Use Efficiency in Response to Dehydration in Ferns and Seed Plants. New Phytol. 2021, 230, 2001–2010. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive Oxygen Species, Oxidative Signaling and the Regulation of Photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Naeem, M.; Shahzad, K.; Saqib, S.; Shahzad, A.; Nasrullah; Younas, M.; Afridi, M.I. The Solanum Melongena COP1LIKE Manipulates Fruit Ripening and Flowering Time in Tomato (Solanum lycopersicum). Plant Growth Regul. 2022, 96, 369–382. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of Drought Stress on Photosynthesis and Photosynthetic Electron Transport Chain in Young Apple Tree Leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.; Shahid, S.; Nazar, N.; Hussain, A.I.; Ali, S.; Chatha, S.A.S.; Perveen, R.; Naseem, J.; Haider, M.Z.; Hussain, B.; et al. Use of Phytohormones in Conferring Tolerance to Environmental Stress. In Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II: Mechanisms of Adaptation and Stress Amelioration; Springer: Berlin/Heidelberg, Germany, 2020; pp. 245–355. ISBN 9789811521720. [Google Scholar]
- Simova-Stoilova, L.; Pecheva, D.; Kirova, E. Drought Stress Response in Winter Wheat Varieties—Changes in Leaf Proteins and Proteolytic Activities. Acta Bot. Croat. 2020, 7, 121–130. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M. Strategy Shifts in Leaf Physiology, Structure and Nutrient Content between Species of High- and Low-Rainfall and High- and Low-Nutrient Habitats. Funct. Ecol. 2001, 15, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Basit, A.; Hafeez, M.B.; Irshad, S.; Bashir, S.; Bashir, S.; Maqbool, M.M.; Saddiq, M.S.; Hasnain, Z.; Aljuaid, B.S.; et al. Moringa Leaf Extract Improves Biochemical Attributes, Yield and Grain Quality of Rice (Oryza sativa L.) under Drought Stress. PLoS ONE 2021, 16, e0254452. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Boyle, D.L.; Welti, R.; Jagadish, S.V.K.; Prasad, P.V.V. Decreased Photosynthetic Rate under High Temperature in Wheat Is Due to Lipid Desaturation, Oxidation, Acylation, and Damage of Organelles. BMC Plant Biol. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Ommen, O.E.; Donnelly, A.; Vanhoutvin, S.; Van Oijen, M.; Manderscheid, R. Chlorophyll Content of Spring Wheat Flag Leaves Grown under Elevated CO2 Concentrations and Other Environmental Stresses within the ‘ESPACE-Wheat’Project. Eur. J. Agron. 1999, 10, 197–203. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, Salinity and Drought Stresses: An Overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Talaat, N.B. Role of Reactive Oxygen Species Signaling in Plant Growth and Development. React. Oxyg. Nitrogen Sulfur Species Plants Prod. Metab. Signal. Def. Mech. 2019, 2019, 225–266. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the Physiological and Biochemical Impacts of Salt Stress on Plant Growth and Development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone Priming: Regulator for Heavy Metal Stress in Plants. J. Plant Growth Regul. 2019, 38, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, K.A.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.S.E.; Abu-Elsaoud, A.M.; Hafez, Y.M. Exogenous Application of Proline and Salicylic Acid Can Mitigate the Injurious Impacts of Drought Stress on Barley Plants Associated with Physiological and Histological Characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Mehmood, S.S.; Tabassum, J.; Batool, R. Targeting Plant Hormones to Develop Abiotic Stress Resistance in Wheat. In Wheat Production in Changing Environments; Springer: Berlin/Heidelberg, Germany, 2019; pp. 557–577. [Google Scholar]
- Mahmood, T.; Rana, R.M.; Ahmar, S.; Saeed, S.; Gulzar, A.; Khan, M.A.; Wattoo, F.M.; Wang, X.; Branca, F.; Mora-Poblete, F.; et al. Effect of Drought Stress on Capsaicin and Antioxidant Contents in Pepper Genotypes at Reproductive Stage. Plants 2021, 10, 1286. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Xiao, C.; Cui, J.; Hao, B.; Zhang, W.; Yang, Z.; Ahammed, G.J.; Liu, H.; Cui, H. Nitric Oxide and Its Interaction with Hydrogen Peroxide Enhance Plant Tolerance to Low Temperatures by Improving the Efficiency of the Calvin Cycle and the Ascorbate–Glutathione Cycle in Cucumber Seedlings. J. Plant Growth Regul. 2021, 40, 2390–2408. [Google Scholar] [CrossRef]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.M.N.; Jamshed, M.; Samuel, M.A. Abiotic Stress Signaling in Wheat—An Inclusive Overview of Hormonal Interactions during Abiotic Stress Responses in Wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.J. Role of Osmolytes and Antioxidant Enzymes for Drought Tolerance in Wheat. Glob. Wheat Prod. 2018, 51. [Google Scholar] [CrossRef] [Green Version]
- Rauf, M.; Munir, M.; Ul Hassan, M.; Ahmad, M.; Afzal, M. Performance of Wheat Genotypes under Osmotic Stress at Germination and Early Seedling Growth Stage. African J. Biotechnol. 2007, 6, 971–975. [Google Scholar] [CrossRef]
- Ghatak, A.; Chaturvedi, P.; Weckwerth, W. Cereal Crop Proteomics: Systemic Analysis of Crop Drought Stress Responses towards Marker-Assisted Selection Breeding. Front. Plant Sci. 2017, 8, 757. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Chowrasia, S.; Gaur, V.S.; Mondal, T.K. Allantoin: Emerging Role in Plant Abiotic Stress Tolerance. Plant Mol. Biol. Rep. 2016, 39, 648–661. [Google Scholar] [CrossRef]
- Dhanda, S.S.; Sethi, G.S.; Behl, R.K. Indices of Drought Tolerance in Wheat Genotypes at Early Stages of Plant Growth. J. Agron. Crop. Sci. 2004, 190, 6–12. [Google Scholar] [CrossRef]
- Talbi, S.; Rojas, J.A.; Sahrawy, M.; Rodríguez-Serrano, M.; Cárdenas, K.E.; Debouba, M.; Sandalio, L.M. Effect of Drought on Growth, Photosynthesis and Total Antioxidant Capacity of the Saharan Plant Oudeneya Africana. Environ. Exp. Bot. 2020, 176, 104099. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive Oxygen Species Homeostasis and Signalling during Drought and Salinity Stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the Regulation of Reactive Oxygen Species Metabolism in Plants under Abiotic Stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Franchina, D.G.; Dostert, C.; Brenner, D. Reactive Oxygen Species: Involvement in T Cell Signaling and Metabolism. Trends Immunol. 2018, 39, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Verma, G.; Srivastava, D.; Tiwari, P.; Chakrabarty, D. ROS Modulation in Crop Plants under Drought Stress. React. Oxyg. Nitrogen Sulfur Species Plants Prod. Metab. Signal. Def. Mech. 2019, 2019, 311–336. [Google Scholar]
- Madzikane-Mlungwana, O.; Moyo, M.; Aremu, A.O.; Plíhalová, L.; Doležal, K.; Van Staden, J.; Finnie, J.F. Differential Responses to Isoprenoid, N 6-Substituted Aromatic Cytokinins and Indole-3-Butyric Acid in Direct Plant Regeneration of Eriocephalus Africanus. Plant Growth Regul. 2017, 82, 103–110. [Google Scholar] [CrossRef]
- Blum, A. Plant Water Relations, Plant Stress and Plant Production. In Plant Breeding for Water-Limited Environments; Springer: Berlin/Heidelberg, Germany, 2011; pp. 11–52. [Google Scholar]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wȩdzony, M. Physiological and Biochemical Tools Useful in Drought-Tolerance Detection in Genotypes of Winter Triticale: Accumulation of Ferulic Acid Correlates with Drought Tolerance. Ann. Bot. 2007, 100, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Sahay, S.; Khan, E.; Gupta, M. Nitric Oxide and Abscisic Acid Protects against PEG-Induced Drought Stress Differentially in Brassica Genotypes by Combining the Role of Stress Modulators, Markers and Antioxidants. Nitric Oxide—Biol. Chem. 2019, 89, 81–92. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of Ascorbic Acid, Glutathione and Proline Applied as Singly or in Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under Different Levels of Irrigation Intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.Y.; Sina, A.A.I.; Khandker, S.S.; Neesa, L.; Tanvir, E.M.; Kabir, A.; Khalil, M.I.; Gan, S.H. Nutritional Composition and Bioactive Compounds in Tomatoes and Their Impact on Human Health and Disease: A Review. Foods 2021, 10, 45. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Sharma, D.; Kapoor, D. Salicylic Acid Signaling and ROS Balance in Plants. Salicylic Acid Contrib. Plant Biol. Chang. Environ. 2021, 2021, 87–114. [Google Scholar]
- Lin, K.H.; Chao, P.Y.; Yang, C.M.; Cheng, W.C.; Lo, H.F.; Chang, T.R. The Effects of Flooding and Drought Stresses on the Antioxidant Constituents in Sweet Potato Leaves. Bot. Stud. 2006, 47, 417–426. [Google Scholar]
- Ahanger, M.A.; Qi, M.; Huang, Z.; Xu, X.; Begum, N.; Qin, C.; Zhang, C.; Ahmad, N.; Mustafa, N.S.; Ashraf, M.; et al. Improving Growth and Photosynthetic Performance of Drought Stressed Tomato by Application of Nano-Organic Fertilizer Involves up-Regulation of Nitrogen, Antioxidant and Osmolyte Metabolism. Ecotoxicol. Environ. Saf. 2021, 216, 112195. [Google Scholar] [CrossRef] [PubMed]
- Guddimalli, R.; Somanaboina, A.K.; Palle, S.R.; Edupuganti, S.; Kummari, D.; Palakolanu, S.R.; Naravula, J.; Gandra, J.; Qureshi, I.A.; Marka, N.; et al. Overexpression of RNA-Binding Bacterial Chaperones in Rice Leads to Stay-Green Phenotype, Improved Yield and Tolerance to Salt and Drought Stresses. Physiol. Plant. 2021, 173, 1351–1368. [Google Scholar] [CrossRef] [PubMed]
- Doneva, D.; Pál, M.; Brankova, L.; Szalai, G.; Tajti, J.; Khalil, R.; Ivanovska, B.; Velikova, V.; Misheva, S.; Janda, T.; et al. The Effects of Putrescine Pre-Treatment on Osmotic Stress Responses in Drought-Tolerant and Drought-Sensitive Wheat Seedlings. Physiol. Plant. 2021, 171, 200–216. [Google Scholar] [CrossRef]
- Al-Ghzawi, A.L.A.; Khalaf, Y.B.; Al-Ajlouni, Z.I.; Al-Quraan, N.A.; Musallam, I.; Hani, N.B. The Effect of Supplemental Irrigation on Canopy Temperature Depression, Chlorophyll Content, and Water Use Efficiency in Three Wheat (Triticum aestivum L. and T. durum Desf.) Varieties Grown in Dry Regions of Jordan. Agriculture 2018, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Anwar, S.; Yu, S.; Sun, M.; Yang, Z.; Gao, Z.Q. Development of Drought-Tolerant Transgenic Wheat: Achievements and Limitations. Int. J. Mol. Sci. 2019, 20, 3350. [Google Scholar] [CrossRef] [Green Version]
- Hassanein, R.A.; Amin, A.B.A.E.S.; Rashad, E.S.M.; Ali, H. Effect of Thiourea and Salicylic Acid on Antioxidant Defense of Wheat Plants under Drought Stress. Int. J. Chem. Tech. Res. 2015, 7, 346–354. [Google Scholar]
- Li, J.; Luan, Y.; Liu, Z. SpWRKY1 Mediates Resistance to Phytophthora Infestans and Tolerance to Salt and Drought Stress by Modulating Reactive Oxygen Species Homeostasis and Expression of Defense-Related Genes in Tomato. Plant Cell. Tissue Organ. Cult. 2015, 123, 67–81. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology (Basel) 2021, 10, 267. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Xiong, H.; Hua, L.; Reyna-Llorens, I.; Shi, Y.; Chen, K.M.; Smirnoff, N.; Kromdijk, J.; Hibberd, J.M. Photosynthesis-Independent Production of Reactive Oxygen Species in the Rice Bundle Sheath during High Light Is Mediated by NADPH Oxidase. Proc. Natl. Acad. Sci. USA 2021, 118, e2022702118. [Google Scholar] [CrossRef]
- Siddiqui, M.N.; León, J.; Naz, A.A.; Ballvora, A. Genetics and Genomics of Root System Variation in Adaptation to Drought Stress in Cereal Crops. J. Exp. Bot. 2021, 72, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.K.; Dubeaux, G.; Takahashi, Y.; Schroeder, J.I. Signaling Mechanisms in Abscisic Acid-Mediated Stomatal Closure. Plant J. 2021, 105, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Lüking, I.; Yang, O. Plant Tolerance to Drought and Salinity: Stress Regulating Transcription Factors and Their Functional Significance in the Cellular Transcriptional Network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Varoquaux, N.; Cole, B.; Gao, C.; Pierroz, G.; Baker, C.R.; Patel, D.; Madera, M.; Jeffers, T.; Hollingsworth, J.; Sievert, J.; et al. Transcriptomic Analysis of Field-Droughted Sorghum from Seedling to Maturity Reveals Biotic and Metabolic Responses. Proc. Natl. Acad. Sci. USA 2019, 116, 27124–27132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Yin, J.; Lu, Y.; Song, J.; Wang, S.; Wei, S.; Liu, Z.; Zhang, Y.; Fang, Z. Transcriptomic Analysis Reveals the Temporal and Spatial Changes in Physiological Process and Gene Expression in Common Buckwheat (Fagopyrum Esculentum Moench) Grown under Drought Stress. Agronomy 2019, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Brozynska, M.; Furtado, A.; Henry, R.J. Genomics of Crop Wild Relatives: Expanding the Gene Pool for Crop Improvement. Plant Biotechnol. J. 2016, 14, 1070–1085. [Google Scholar] [CrossRef]
- Zhu, M.; Monroe, J.G.; Suhail, Y.; Villiers, F.; Mullen, J.; Pater, D.; Hauser, F.; Jeon, B.W.; Bader, J.S.; Kwak, J.M.; et al. Molecular and Systems Approaches towards Drought-Tolerant Canola Crops. New Phytol. 2016, 210, 1169–1189. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.L.; Grondin, A.; Courtois, B.; Gantet, P. Next-Generation Sequencing Accelerates Crop Gene Discovery. Trends Plant Sci. 2019, 24, 263–274. [Google Scholar] [CrossRef]
- Martineau, C.; Li, X.; Lalancette, C.; Perreault, T.; Fournier, E.; Tremblay, J.; Gonzales, M.; Yergeau, É.; Quach, C. Serratia Marcescens Outbreak in a Neonatal Intensive Care Unit: New Insights from next-Generation Sequencing Applications. J. Clin. Microbiol. 2018, 56, e00235-18. [Google Scholar] [CrossRef] [Green Version]
- Zenda, T.; Liu, S.; Duan, H. Adapting Cereal Grain Crops to Drought Stress: 2020 and Beyond. Abiotic Stress Plants 2020, 2020, 1–30. [Google Scholar]
- Kumar, A.; Saripalli, G.; Jan, I.; Kumar, K.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Meta-QTL Analysis and Identification of Candidate Genes for Drought Tolerance in Bread Wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2020, 26, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol 2019, 70, 667–697. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells 2020, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alagoz, Y.; Gurkok, T.; Zhang, B.; Unver, T. Manipulating the Biosynthesis of Bioactive Compound Alkaloids for Next-Generation Metabolic Engineering in Opium Poppy Using CRISPR-Cas 9 Genome Editing Technology. Sci. Rep. 2016, 6, 30910. [Google Scholar] [CrossRef] [Green Version]
- Martignago, D.; Rico-Medina, A.; Blasco-Escámez, D.; Fontanet-Manzaneque, J.B.; Caño-Delgado, A.I. Drought Resistance by Engineering Plant Tissue-Specific Responses. Front. Plant Sci. 2020, 10, 1676. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, M.; Kim, J.-H.; Kim, J.A.; Lee, S.-I. Plant RNA Binding Proteins as Critical Modulators in Drought, High Salinity, Heat, and Cold Stress Responses: An Updated Overview. Int. J. Mol. Sci. 2021, 22, 6731. [Google Scholar] [CrossRef]
- Caddell, D.F.; Deng, S.; Coleman-Derr, D. Role of the Plant Root Microbiome in Abiotic Stress Tolerance. In Seed Endophytes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 273–311. [Google Scholar]
- Sattiraju, K.S.; Kotiyal, S.; Arora, A.; Maheshwari, M. Plant Growth-Promoting Microbes: Contribution to Stress Management in Plant Hosts. Environ. Biotechnol. Sustain. Futur. 2019, 2019, 199–236. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones Enhanced Drought Tolerance in Plants: A Coping Strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef]
- Chumikina, L.V.; Arabova, L.I.; Kolpakova, V.V.; Topunov, A.F. The Role of Phytohormones in the Regulation of the Tolerance of Wheat, Rye, and Triticale Seeds to the Effect of Elevated Temperatures during Germination. Appl. Biochem. Microbiol. 2019, 55, 59–66. [Google Scholar] [CrossRef]
- Jogawat, A.; Yadav, B.; Lakra, N.; Singh, A.K.; Narayan, O.P. Crosstalk between Phytohormones and Secondary Metabolites in the Drought Stress Tolerance of Crop Plants: A Review. Physiol. Plant. 2021, 172, 1106–1132. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical Regulators of Plant Hormones and Their Applications in Basic Research and Agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How Plant Hormones Mediate Salt Stress Responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Dutta, P.; Chakrabarty, D. MiRNAs Play Critical Roles in Response to Abiotic Stress by Modulating Cross-Talk of Phytohormone Signaling. Plant Cell Rep. 2021, 40, 1617–1630. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Charagh, S.; Zahid, Z.; Mubarik, M.S.; Javed, R.; Siddiqui, M.H.; Hasanuzzaman, M. Jasmonic Acid: A Key Frontier in Conferring Abiotic Stress Tolerance in Plants. Plant Cell Rep. 2021, 40, 1513–1541. [Google Scholar] [CrossRef]
- Yadav, A.N.; Yadav, N. Stress-Adaptive Microbes for Plant Growth Promotion and Alleviation of Drought Stress in Plants. Acta Sci. Agric. 2018, 2, 85–88. [Google Scholar]
- Campos-Rivero, G.; Osorio-Montalvo, P.; Sánchez-Borges, R.; Us-Camas, R.; Duarte-Aké, F.; De-la-Peña, C. Plant Hormone Signaling in Flowering: An Epigenetic Point of View. J. Plant Physiol. 2017, 214, 16–27. [Google Scholar] [CrossRef]
- Sheng, J.; Li, X.; Zhang, D. Gibberellins, Brassinolide, and Ethylene Signaling Were Involved in Flower Differentiation and Development in Nelumbo Nucifera. Hortic. Plant J. 2022, 8, 243–250. [Google Scholar] [CrossRef]
- Waterman, P.G.; Mole, S. Extrinsic Factors Influencing Production of Secondary Metabolites in Plants. In Insect-Plant Interactions; CRC Press: Boca Raton, FL, USA, 2019; pp. 107–134. ISBN 0429290918. [Google Scholar]
- Mir, R.A.; Bhat, B.A.; Yousuf, H.; Islam, S.T.; Raza, A.; Rizvi, M.A.; Charagh, S.; Albaqami, M.; Sofi, P.A.; Zargar, S.M. Multidimensional Role of Silicon to Activate Resilient Plant Growth and to Mitigate Abiotic Stress. Front. Plant Sci. 2022, 13, 819658. [Google Scholar] [CrossRef]
- Iqbal, N.; Fatma, M.; Gautam, H.; Sehar, Z.; Rasheed, F.; Khan, M.I.R.; Sofo, A.; Khan, N.A. Salicylic Acid Increases Photosynthesis of Drought Grown Mustard Plants Effectively with Sufficient-N via Regulation of Ethylene, Abscisic Acid, and Nitrogen-Use Efficiency. J. Plant Growth Regul. 2022, 2022, 1–12. [Google Scholar] [CrossRef]
- Yadav, B.; Jogawat, A.; Gnanasekaran, P.; Kumari, P.; Lakra, N.; Lal, S.K.; Pawar, J.; Narayan, O.P. An Overview of Recent Advancement in Phytohormones-Mediated Stress Management and Drought Tolerance in Crop Plants. Plant Gene 2021, 25, 100264. [Google Scholar]
- Signorelli, S.; Tarkowski, Ł.P.; Van den Ende, W.; Bassham, D.C. Linking Autophagy to Abiotic and Biotic Stress Responses. Trends Plant Sci. 2019, 24, 413–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llanes, A.; Andrade, A.; Alemano, S.; Luna, V. Alterations of Endogenous Hormonal Levels in Plants under Drought and Salinity. Am. J. Plant Sci. 2016, 7, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, S.; Gómez-Bellot, M.J.; Acosta-Motos, J.R.; Sánchez-Blanco, M.J. Application of Deficit Irrigation in Phillyrea Angustifolia for Landscaping Purposes. Agric. Water Manag. 2019, 218, 193–202. [Google Scholar] [CrossRef]
- Shen, Q.; Liu, Y.; Naqvi, N.I. Fungal Effectors at the Crossroads of Phytohormone Signaling. Curr. Opin. Microbiol. 2018, 46, 1–6. [Google Scholar] [CrossRef]
- La, V.H.; Lee, B.R.; Zhang, Q.; Park, S.H.; Islam, M.T.; Kim, T.H. Salicylic Acid Improves Drought-Stress Tolerance by Regulating the Redox Status and Proline Metabolism in Brassica Rapa. Hortic. Environ. Biotechnol. 2019, 60, 31–40. [Google Scholar] [CrossRef]
- Castro, P.H.; Couto, D.; Freitas, S.; Verde, N.; Macho, A.P.; Huguet, S.; Botella, M.A.; Ruiz-Albert, J.; Tavares, R.M.; Bejarano, E.R.; et al. SUMO Proteases ULP1c and ULP1d Are Required for Development and Osmotic Stress Responses in Arabidopsis Thaliana. Plant Mol. Biol. 2016, 92, 143–159. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Chourasia, K.N.; Naga, K.C.; Kumar, D.; Das, S.K.; Zinta, G. Mechanistic Insights on Melatonin-mediated Drought Stress Mitigation in Plants. Physiol. Plant. 2021, 172, 1212–1226. [Google Scholar] [CrossRef]
- Khalvandi, M.; Siosemardeh, A.; Roohi, E.; Keramati, S. Salicylic Acid Alleviated the Effect of Drought Stress on Photosynthetic Characteristics and Leaf Protein Pattern in Winter Wheat. Heliyon 2021, 7, e05908. [Google Scholar] [CrossRef]
- Garg, N.; Bharti, A. Salicylic Acid Improves Arbuscular Mycorrhizal Symbiosis, and Chickpea Growth and Yield by Modulating Carbohydrate Metabolism under Salt Stress. Mycorrhiza 2018, 28, 727–746. [Google Scholar] [CrossRef]
- Bandurska, H. Salicylic Acid: An Update on Biosynthesis and Action in Plant Response to Water Deficit and Performance Under Drought. In Salicylic Acid; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–14. [Google Scholar]
- Jahan, M.S.; Wang, Y.; Shu, S.; Zhong, M.; Chen, Z.; Wu, J.; Sun, J.; Guo, S. Exogenous Salicylic Acid Increases the Heat Tolerance in Tomato (Solanum lycopersicum L) by Enhancing Photosynthesis Efficiency and Improving Antioxidant Defense System through Scavenging of Reactive Oxygen Species. Sci. Hortic. (Amsterdam) 2019, 247, 421–429. [Google Scholar] [CrossRef]
- Otálora, G.; Piñero, M.C.; Collado-González, J.; López-Marín, J.; Del Amor, F.M. Exogenous Salicylic Acid Modulates the Response to Combined Salinity-Temperature Stress in Pepper Plants (Capsicum annuum L. Var. Tamarin). Plants 2020, 9, 1790. [Google Scholar] [CrossRef] [PubMed]
- Torun, H. Time-Course Analysis of Salicylic Acid Effects on ROS Regulation and Antioxidant Defense in Roots of Hulled and Hulless Barley under Combined Stress of Drought, Heat and Salinity. Physiol. Plant. 2019, 165, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Salem, K.F.M.; Saleh, M.M.; Abu-Ellail, F.F.B.; Aldahak, L.; Alkuddsi, Y.A. The Role of Salicylic Acid in Crops to Tolerate Abiotic Stresses. In Salicylic Acid-A Versatile Plant Growth Regulator; Springer: Berlin/Heidelberg, Germany, 2021; pp. 93–152. [Google Scholar]
- Maghsoudi, K.; Emam, Y.; Ashraf, M.; Arvin, M.J. Alleviation of Field Water Stress in Wheat Cultivars by Using Silicon and Salicylic Acid Applied Separately or in Combination. Crop Pasture Sci. 2019, 70, 36–43. [Google Scholar] [CrossRef]
- Munsif, F.; Shah, T.; Arif, M.; Jehangir, M.; Afridi, M.Z.; Ahmad, I.; Jan, B.L.; Alansi, S. Combined Effect of Salicylic Acid and Potassium Mitigates Drought Stress through the Modulation of Physio-Biochemical Attributes and Key Antioxidants in Wheat. Saudi J. Biol. Sci. 2022, 29, 103294. [Google Scholar] [CrossRef] [PubMed]
- Saheri, F.; Barzin, G.; Pishkar, L.; Boojar, M.M.A.; Babaeekhou, L. Correction to: Foliar Spray of Salicylic Acid Induces Physiological and Biochemical Changes in Purslane (Portulaca oleracea L.) under Drought Stress (Biologia, (2020), 10.2478/S11756-020-00571-2). Biologia (Bratisl) 2020, 75, 2201. [Google Scholar] [CrossRef]
- Seif El-Yazal, S.; Seif El-Yazal, M.; Dwidar, E.; Rady, M. Phytohormone Crosstalk Research: Cytokinin and Its Crosstalk with Other Phytohormones. Curr. Protein Pept. Sci. 2015, 16, 395–405. [Google Scholar] [CrossRef]
- Vrabka, J.; Niehaus, E.-M.; Münsterkötter, M.; Proctor, R.H.; Brown, D.W.; Novák, O.; Pěnčik, A.; Tarkowská, D.; Hromadová, K.; Hradilová, M. Production and Role of Hormones during Interaction of Fusarium Species with Maize (Zea mays L.) Seedlings. Front. Plant Sci. 2019, 9, 1936. [Google Scholar] [CrossRef]
- Tarkowská, D.; Strnad, M. Isoprenoid-Derived Plant Signaling Molecules: Biosynthesis and Biological Importance. Planta 2018, 247, 1051–1066. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Kojima, M.; Yamaya, T.; Sakakibara, H. Molecular Characterization of Cytokinin-Responsive Histidine Kinases in Maize. Differential Ligand Preferences and Response to Cis-Zeatin. Plant Physiol. 2004, 134, 1654–1661. [Google Scholar] [CrossRef] [Green Version]
- Bidon, B.; Kabbara, S.; Courdavault, V.; Glévarec, G.; Oudin, A.; Héricourt, F.; Carpin, S.; Spíchal, L.; Binder, B.M.; Cock, J.M. Cytokinin and Ethylene Cell Signaling Pathways from Prokaryotes to Eukaryotes. Cells 2020, 9, 2526. [Google Scholar] [CrossRef]
- Borghi, L.; Kang, J.; de Brito Francisco, R. Filling the Gap: Functional Clustering of ABC Proteins for the Investigation of Hormonal Transport in Planta. Front. Plant Sci. 2019, 10, 422. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Munné-Bosch, S. Hormonal Impact on Photosynthesis and Photoprotection in Plants. Plant Physiol. 2021, 185, 1500–1522. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Negi, N.P.; Pareek, S.; Mudgal, G.; Kumar, D. Auxin Response Factors in Plant Adaptation to Drought and Salinity Stress. Physiol. Plant. 2022, 2022, e13714. [Google Scholar] [CrossRef] [PubMed]
- Khosravi-nejad, F.; Khavari-nejad, R.A.; Moradi, F.; Najafi, F. Cytokinin and Abscisic Acid Alleviate Drought Stress through Changing Organic Acids Profile, Ion Immolation, and Fatty Acid Profile to Improve Yield of Wheat (Triticum aestivum L.) Cultivars. Physiol. Mol. Biol. Plants 2022, 28, 1119–1129. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Biochar Increased Plant Growth-Promoting Hormones and Helped to Alleviates Salt Stress in Common Bean Seedlings. J. Plant Growth Regul. 2018, 37, 591–601. [Google Scholar] [CrossRef]
- Deng, Y.; Zhou, Q.; Wu, Y.; Chen, X.; Zhong, F. Properties and Mechanisms of Flavin-Dependent Monooxygenases and Their Applications in Natural Product Synthesis. Int. J. Mol. Sci. 2022, 23, 2622. [Google Scholar] [CrossRef]
- Ronzan, M.; Piacentini, D.; Fattorini, L.; Caboni, E.; Eiche, E.; Ziegler, J.; Hause, B.; Riemann, M.; Betti, C.; Altamura, M.M. Auxin-Jasmonate Crosstalk in Oryza Sativa L. Root System Formation after Cadmium and/or Arsenic Exposure. Environ. Exp. Bot. 2019, 165, 59–69. [Google Scholar] [CrossRef]
- Kalra, G.; Bhatla, S.C. Gibberellins. In Plant Physiology, Development and Metabolism; Springer: Berlin/Heidelberg, Germany, 2018; pp. 617–628. [Google Scholar]
- Sedaghat, M.; Emam, Y.; Mokhtassi-Bidgoli, A.; Hazrati, S.; Lovisolo, C.; Visentin, I.; Cardinale, F.; Tahmasebi-Sarvestani, Z. The Potential of the Synthetic Strigolactone Analogue GR24 for the Maintenance of Photosynthesis and Yield in Winter Wheat under Drought: Investigations on the Mechanisms of Action and Delivery Modes. Plants 2021, 10, 1223. [Google Scholar] [CrossRef]
- Rasheed, A.; Hassan, M.U.; Aamer, M.; Batool, M.; Sheng, F.; Ziming, W.U.; Huijie, L.I. A Critical Review on the Improvement of Drought Stress Tolerance in Rice (Oryza sativa L.). Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1756–1788. [Google Scholar] [CrossRef]
- Salvi, P.; Manna, M.; Kaur, H.; Thakur, T.; Gandass, N.; Bhatt, D.; Muthamilarasan, M. Phytohormone Signaling and Crosstalk in Regulating Drought Stress Response in Plants. Plant Cell Rep. 2021, 40, 1305–1329. [Google Scholar] [CrossRef]
- Choudhary, P.; Pramitha, L.; Rana, S.; Verma, S.; Aggarwal, P.R.; Muthamilarasan, M. Hormonal Crosstalk in Regulating Salinity Stress Tolerance in Graminaceous Crops. Physiol. Plant. 2021, 173, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F. The Role of Salicylic Acid and Gibberellin Signaling in Plant Responses to Abiotic Stress with an Emphasis on Heavy Metals. Plant Signal. Behav. 2020, 15, 1777372. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. The Current Status of Research on Gibberellin Biosynthesis. Plant Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef] [PubMed]
- Vishal, B.; Kumar, P.P. Regulation of Seed Germination and Abiotic Stresses by Gibberellins and Abscisic Acid. Front. Plant Sci. 2018, 9, 838. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cheng, Z.; Liu, L.; Wang, M.; You, X.; Wang, J.; Zhang, F.; Zhou, C.; Zhang, Z.; Zhang, H. Small Grain and Dwarf 2, Encoding an HD-Zip II Family Transcription Factor, Regulates Plant Development by Modulating Gibberellin Biosynthesis in Rice. Plant Sci. 2019, 288, 110208. [Google Scholar] [CrossRef]
- Goldschmidt, E.E.; Sadka, A. Yield Alternation: Horticulture, Physiology, Molecular Biology, and Evolution. Hortic. Rev. (Am. Soc. Hortic. Sci). 2021, 48, 363–418. [Google Scholar]
- Verbancic, J. Carbon Supply and the Regulation of Primary Cell Wall Synthesis in Arabidopsis Thaliana 2021. Doctoral Dissertation, Universität Potsdam, Potsdam, Germany, 2021. [Google Scholar]
- Omena-Garcia, R.P.; Martins, A.O.; Medeiros, D.B.; Vallarino, J.G.; Ribeiro, D.M.; Fernie, A.R.; Araújo, W.L.; Nunes-Nesi, A. Growth and Metabolic Adjustments in Response to Gibberellin Deficiency in Drought Stressed Tomato Plants. Environ. Exp. Bot. 2019, 159, 95–107. [Google Scholar] [CrossRef]
- Salazar-Cerezo, S.; Martínez-Montiel, N.; García-Sánchez, J.; Pérez-y-Terrón, R.; Martínez-Contreras, R.D. Gibberellin Biosynthesis and Metabolism: A Convergent Route for Plants, Fungi and Bacteria. Microbiol. Res. 2018, 208, 85–98. [Google Scholar] [CrossRef]
- Kumar, B. Plant Bio-Regulators for Enhancing Grain Yield and Quality of Legumes: A Review. Agric. Rev. 2021, 42, 175–182. [Google Scholar] [CrossRef]
- Draweel, M.M.; Soegianto, A.; Soetopo, L.; Kuswanto, K. Evaluation of Some Morphological Criteria to Drought Tolerance on Seedling of Bambara Groundnut [Vigna subterranea (L.) Verdc.] Using Polyethylene Glycol (Peg6000). Legum. Res. Int. J. 2021, 1, 10. [Google Scholar] [CrossRef]
- Rademacher, W. Chemical Regulators of Gibberellin Status and Their Application in Plant Production. Annu. Plant Rev. Online 2018, 2018, 359–403. [Google Scholar]
- Shohat, H.; Cheriker, H.; Kilambi, H.V.; Illouz Eliaz, N.; Blum, S.; Amsellem, Z.; Tarkowská, D.; Aharoni, A.; Eshed, Y.; Weiss, D. Inhibition of Gibberellin Accumulation by Water Deficiency Promotes Fast and Long-term ‘Drought Avoidance’Responses in Tomato. New Phytol. 2021, 232, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Litvin, A.G.; van Iersel, M.W.; Malladi, A. Drought Stress Reduces Stem Elongation and Alters Gibberellin-Related Gene Expression during Vegetative Growth of Tomato. J. Am. Soc. Hortic. Sci. 2016, 141, 591–597. [Google Scholar] [CrossRef]
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin Localization and Transport in Plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Hayat, K.; Iqbal, A.; Xie, L. Implications of Abscisic Acid in the Drought Stress Tolerance of Plants. Agronomy 2020, 10, 1323. [Google Scholar] [CrossRef]
- Waterland, N.L.; Campbell, C.A.; Finer, J.J.; Jones, M.L. Abscisic Acid Application Enhances Drought Stress Tolerance in Bedding Plants. HortScience 2010, 45, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, P.; Wang, G.; Augstein, F.; de Vries, J.; Carlsbecker, A. Continuous Root Xylem Formation and Vascular Acclimation to Water Deficit Involves Endodermal ABA Signalling via MiR165. Development 2018, 145, dev159202. [Google Scholar] [CrossRef] [Green Version]
- Fraudentali, I.; Ghuge, S.A.; Carucci, A.; Tavladoraki, P.; Angelini, R.; Rodrigues-Pousada, R.A.; Cona, A. Developmental, Hormone-and Stress-Modulated Expression Profiles of Four Members of the Arabidopsis Copper-Amine Oxidase Gene Family. Plant Physiol. Biochem. 2020, 147, 141–160. [Google Scholar] [CrossRef]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The Role of Stress-Responsive Transcription Factors in Modulating Abiotic Stress Tolerance in Plants. Agronomy 2020, 10, 788. [Google Scholar] [CrossRef]
- Chen, R.; Ma, J.; Luo, D.; Hou, X.; Ma, F.; Zhang, Y.; Meng, Y.; Zhang, H.; Guo, W. CaMADS, a MADS-Box Transcription Factor from Pepper, Plays an Important Role in the Response to Cold, Salt, and Osmotic Stress. Plant Sci. 2019, 280, 164–174. [Google Scholar] [CrossRef]
- Khan, A.; Pan, X.; Najeeb, U.; Tan, D.K.Y.; Fahad, S.; Zahoor, R.; Luo, H. Coping with Drought: Stress and Adaptive Mechanisms, and Management through Cultural and Molecular Alternatives in Cotton as Vital Constituents for Plant Stress Resilience and Fitness. Biol. Res. 2018, 51. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.E. Advances in Transgenic Technology for Crop Cultivation and Stomatal Regulation as Potent Role in Agriculture. Sch. Int. J. Biochem. 2021, 4, 86–90. [Google Scholar]
- Wang, N.-N.; Xu, S.-W.; Sun, Y.-L.; Liu, D.; Zhou, L.; Li, Y.; Li, X.-B. The Cotton WRKY Transcription Factor (GhWRKY33) Reduces Transgenic Arabidopsis Resistance to Drought Stress. Sci. Rep. 2019, 9, 724. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Azhar, M.T.; Hinze, L.; Qayyum, A.; Li, H.; Peng, Z.; Qin, G.; Jia, Y.; Pan, Z.; He, S. Insight into Abscisic Acid Perception and Signaling to Increase Plant Tolerance to Abiotic Stress. J. Plant Interact. 2021, 16, 222–237. [Google Scholar] [CrossRef]
- Ali, F.; Qanmber, G.; Li, F.; Wang, Z. Updated Role of ABA in Seed Maturation, Dormancy, and Germination. J. Adv. Res. 2022, 35, 199–214. [Google Scholar] [CrossRef]
- Yang, W.H.; Lu, C.Z.; Chen, W.; Xu, H.Y. Reduction of Early Fruit Abscission by Main-Branch-Girdling in Macadamia Is Related to the Favorable Status of Carbohydrates and Endogenous Hormones. HortScience 2022, 57, 40–47. [Google Scholar] [CrossRef]
- Razi, K.; Muneer, S. Drought Stress-Induced Physiological Mechanisms, Signaling Pathways and Molecular Response of Chloroplasts in Common Vegetable Crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef]
- Parveen, A.; Ahmar, S.; Kamran, M.; Malik, Z.; Ali, A.; Riaz, M.; Abbasi, G.H.; Khan, M.; Sohail, A.B.; Rizwan, M. Abscisic Acid Signaling Reduced Transpiration Flow, Regulated Na+ Ion Homeostasis and Antioxidant Enzyme Activities to Induce Salinity Tolerance in Wheat (Triticum aestivum L.) Seedlings. Environ. Technol. Innov. 2021, 24, 101808. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic Acid Dynamics, Signaling, and Functions in Plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of Polyamines, Abscisic Acid and Proline under Osmotic Stress in the Leaves of Wheat Plants. Sci. Rep. 2018, 8, 12839. [Google Scholar] [CrossRef]
- Ouledali, S.; Ennajeh, M.; Ferrandino, A.; Khemira, H.; Schubert, A.; Secchi, F. Influence of Arbuscular Mycorrhizal Fungi Inoculation on the Control of Stomata Functioning by Abscisic Acid (ABA) in Drought-Stressed Olive Plants. S. Afr. J. Bot. 2019, 121, 152–158. [Google Scholar] [CrossRef]
- El-Yazied, A.A.; Ibrahim, M.F.M.; Ibrahim, M.A.R.; Nasef, I.N.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Alzuaibr, F.M.; Alaklabi, A.; Dessoky, E.S.; Alabdallah, N.M. Melatonin Mitigates Drought Induced Oxidative Stress in Potato Plants through Modulation of Osmolytes, Sugar Metabolism, ABA Homeostasis and Antioxidant Enzymes. Plants 2022, 11, 1151. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhu, H.; Zhu, H.; Tao, Y.; Liu, C.; Liu, J.; Yang, F.; Li, M. Exogenous ABA Enhances the Antioxidant Defense System of Maize by Regulating the AsA-GSH Cycle under Drought Stress. Sustainability 2022, 14, 3071. [Google Scholar] [CrossRef]
- Xing, X.; Cao, C.; Xu, Z.; Qi, Y.; Fei, T.; Jiang, H.; Wang, X. Reduced Soybean Water Stress Tolerance by MiR393a-Mediated Repression of GmTIR1 and Abscisic Acid Accumulation. J. Plant Growth Regul. 2022, 2022, 1–17. [Google Scholar] [CrossRef]
- Yari Kamrani, Y.; Shomali, A.; Aliniaeifard, S.; Lastochkina, O.; Moosavi-Nezhad, M.; Hajinajaf, N.; Talar, U. Regulatory Role of Circadian Clocks on ABA Production and Signaling, Stomatal Responses, and Water-Use Efficiency under Water-Deficit Conditions. Cells 2022, 11, 1154. [Google Scholar] [CrossRef]
- Rehman, R.S.; Ali, M.; Zafar, S.A.; Hussain, M.; Pasha, A.; Naveed, M.S.; Ahmad, M.; Waseem, M. Abscisic Acid Mediated Abiotic Stress Tolerance in Plants. Asian J. Res. C Sci. 2022, 7, 1–17. [Google Scholar] [CrossRef]
- Huang, T.-H.; Hsu, W.-H.; Mao, W.-T.; Yang, C.-H. The Oncidium Ethylene Synthesis Gene Oncidium 1-Aminocyclopropane-1 Carboxylic Acid Synthase 12 and Ethylene Receptor Gene Oncidium ETR1 Affect GA-DELLA and Jasmonic Acid Signaling in Regulating Flowering Time, Anther Dehiscence, and Flower Senescence In. Front. Plant Sci. 2022, 13, 785441. [Google Scholar] [CrossRef]
- Patil, S.V.; Patil, C.D.; Mohite, B.V. Isolation and Screening of ACC Deaminase-Producing Microbes for Drought Stress Management in Crops. In Practical Handbook on Agricultural Microbiology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 361–367. [Google Scholar]
- Chandwani, S.; Amaresan, N. Role of ACC Deaminase Producing Bacteria for Abiotic Stress Management and Sustainable Agriculture Production. Environ. Sci. Pollut. Res. 2022, 29, 22843–22859. [Google Scholar] [CrossRef]
- Gautam, H.; Fatma, M.; Sehar, Z.; Iqbal, N.; Albaqami, M.; Khan, N.A. Exogenously-Sourced Ethylene Positively Modulates Photosynthesis, Carbohydrate Metabolism, and Antioxidant Defense to Enhance Heat Tolerance in Rice. Int. J. Mol. Sci. 2022, 23, 1031. [Google Scholar] [CrossRef]
- Costa, L.C.; Luz, L.M.; Nascimento, V.L.; Araujo, F.F.; Santos, M.N.S.; França, C.d.F.M.; Silva, T.P.; Fugate, K.K.; Finger, F.L. Selenium-Ethylene Interplay in Postharvest Life of Cut Flowers. Front. Plant Sci. 2020, 11, 2055. [Google Scholar] [CrossRef]
- Meena, S.; Taria, S.; Nagar, S.; Yadav, S. Phytohormone Engineering: A Potential Approach for Inducing Abiotic Stress Tolerance in Crop Plants. Multidisciplinary 2022, 2022, 35. [Google Scholar]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-Carboxylic Acid Oxidase (ACO): The Enzyme That Makes the Plant Hormone Ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Pérez, J.G.; Puertolas, J.; Albacete, A.; Dodd, I.C. Alternation of Wet and Dry Sides during Partial Rootzone Drying Irrigation Enhances Leaf Ethylene Evolution. Environ. Exp. Bot. 2020, 176, 104095. [Google Scholar] [CrossRef]
- Chung, K.M.; Demianski, A.J.; Harrison, G.A.; Laurie-Berry, N.; Mitsuda, N.; Kunkel, B.N. Jasmonate Hypersensitive 3 (JAH3) Negatively Regulates Both Jasmonate and Ethylene-Mediated Responses in Arabidopsis. J. Exp. Bot. 2022. [Google Scholar] [CrossRef]
- Kazan, K. Diverse Roles of Jasmonates and Ethylene in Abiotic Stress Tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, K.S.; Husen, A. Plant Response to Jasmonates: Current Developments and Their Role in Changing Environment. Bull. Natl. Res. Cent. 2019, 43, 153. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of Sodium Chloride Toxicity in Solanum lycopersicum L. By Supplementation of Jasmonic Acid and Nitric Oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of Jasmonic Acid in Plants: The Molecular Point of View. Plant Cell Rep. 2021, 40, 1471–1494. [Google Scholar] [CrossRef]
- Zaid, A.; Mohammad, F. Methyl Jasmonate and Nitrogen Interact to Alleviate Cadmium Stress in Mentha Arvensis by Regulating Physio-Biochemical Damages and ROS Detoxification. J. Plant Growth Regul. 2018, 37, 1331–1348. [Google Scholar] [CrossRef]
- Ali, M.S.; Baek, K.H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef] [Green Version]
- Zamani, H.; Arvin, M.J.; Jahromi, A.A.; Abdossi, V.; Torkashvand, A.M. The Effect of Sodium Silicate and Methyl Jasmonate on Pigments and Antioxidant Activity of Tomato (Solanum lycopersicum L.) under Salinity Stress. Tarim Bilim. Derg. 2020, 26, 479–487. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, M.I.R.; Anjum, N.A.; Masood, A.; Hussain, S.J.; Khan, N.A. Jasmonates in Plants under Abiotic Stresses: Crosstalk with Other Phytohormones Matters. Environ. Exp. Bot. 2018, 145, 104–120. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. https://doi.org/10.3390/plants11131620
Wahab A, Abdi G, Saleem MH, Ali B, Ullah S, Shah W, Mumtaz S, Yasin G, Muresan CC, Marc RA. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants. 2022; 11(13):1620. https://doi.org/10.3390/plants11131620
Chicago/Turabian StyleWahab, Abdul, Gholamreza Abdi, Muhammad Hamzah Saleem, Baber Ali, Saqib Ullah, Wadood Shah, Sahar Mumtaz, Ghulam Yasin, Crina Carmen Muresan, and Romina Alina Marc. 2022. "Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review" Plants 11, no. 13: 1620. https://doi.org/10.3390/plants11131620