
Overexpression of CsMIXTA, a Transcription Factor from Cannabis sativa, Increases Glandular Trichome Density in Tobacco Leaves


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. CsMIXTA Is Upregulated during Female Flower Development
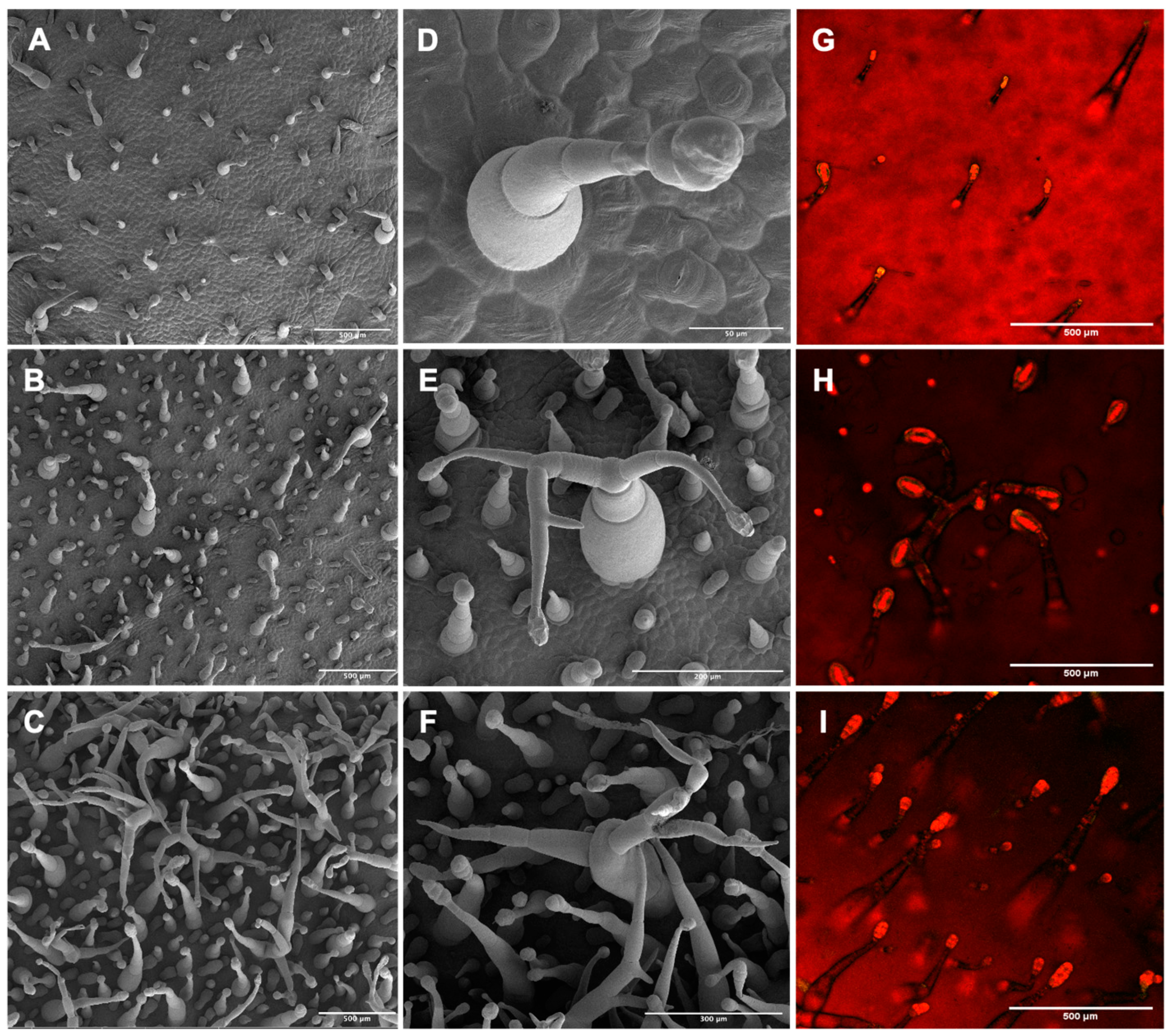
2.2. CsMIXTA Promotes Glandular Trichome Formation in Tobacco Leaves
3. Discussion
4. Materials and Methods
4.1. Cannabis Plant Growth
4.2. Isolation of RNA and cDNA Synthesis
4.3. Isolation of Cannabis Trichomes
4.4. Analysis of CsMIXTA Expression Using qPCR
4.5. Molecular Cloning of CsMIXTA
4.6. Agrobacterium Mediated Transformation of Nicotiana tabacum
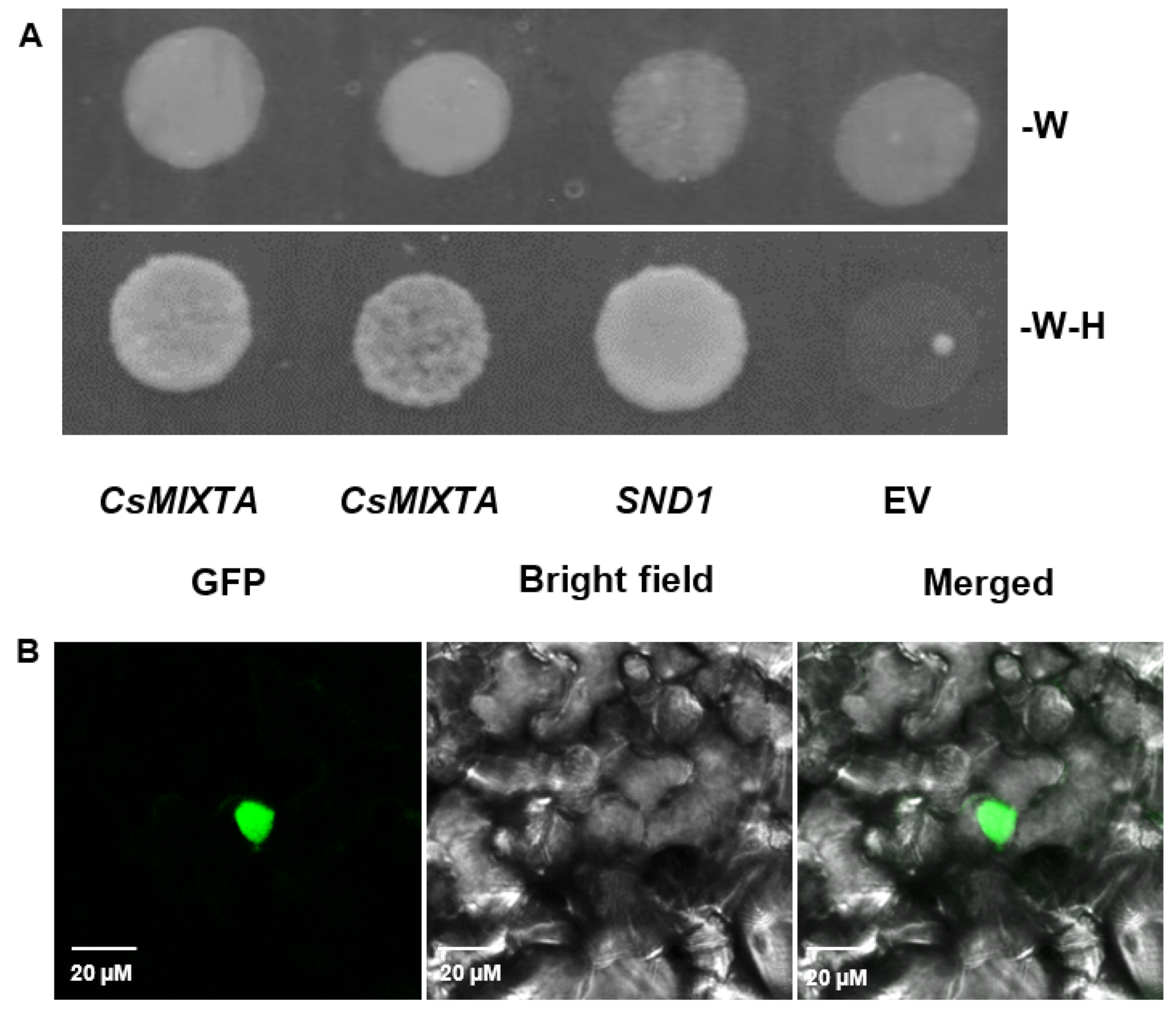
4.7. Yeast Transcriptional Activation Assay
4.8. Transient Expression of CsMIXTA in N. benthamiana Leaves
4.9. Scanning Electron Microscopy
4.10. Fluorescence and Confocal Microscopy
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Accession Numbers
References
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant Glandular Trichomes: Natural Cell Factories of High Biotechnological Interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [Green Version]
- Schuurink, R.; Tissier, A. Glandular Trichomes: Micro-organs with Model Status? New Phytol. 2020, 225, 2251–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, V.; Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Morpho-Anatomy of Marijuana (Cannabis sativa L.). In Cannabis sativa L.—Botany and Biotechnology; Chandra, S., Lata, H., ElSohly, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 123–136. ISBN 978-3-319-54563-9. [Google Scholar]
- Dayanandan, P.; Kaufman, P.B. Trichomes of Cannabis sativa L. (Cannabaceae). Am. J. Bot. 1976, 63, 578–591. [Google Scholar] [CrossRef] [Green Version]
- Hammond, C.T.; Mahlberg, P.G. Morphology of glandular hairs of Cannabis sativa from scanning electron microscopy. Am. J. Bot. 1973, 60, 524–528. [Google Scholar] [CrossRef]
- Hammond, C.T.; Mahlberg, P.G. Morphogenesis of capitate glandular hairs of Cannabis sativa (Cannabaceae). Am. J. Bot. 1977, 64, 1023–1031. [Google Scholar] [CrossRef]
- Gülck, T.; Møller, B.L. Phytocannabinoids: Origins and Biosynthesis. Trends Plant Sci. 2020, 25, 985–1004. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Mahlberg, P.G. Secretory Vesicle Formation in the Secretory Cavity of Glandular Trichomes of Cannabis sativa L. (Cannabaceae). Mol. Cells 2003, 15, 387–395. [Google Scholar] [PubMed]
- Matías-Hernández, L.; Aguilar-Jaramillo, A.E.; Cigliano, R.A.; Sanseverino, W.; Pelaz, S. Flowering and Trichome Development Share Hormonal and Transcription Factor Regulation. J. Exp. Bot. 2016, 67, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis Glandular Trichomes Alter Morphology and Metabolite Content during Flower Maturation. Plant J. 2020, 101, 37–56. [Google Scholar] [CrossRef]
- Chalvin, C.; Drevensek, S.; Dron, M.; Bendahmane, A.; Boualem, A. Genetic Control of Glandular Trichome Development. Trends Plant Sci. 2020, 25, 477–487. [Google Scholar] [CrossRef]
- Shi, P.; Fu, X.; Shen, Q.; Liu, M.; Pan, Q.; Tang, Y.; Jiang, W.; Lv, Z.; Yan, T.; Ma, Y.; et al. The Roles of AaMIXTA1 in Regulating the Initiation of Glandular Trichomes and Cuticle Biosynthesis in Artemisia annua. New Phytol. 2018, 217, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Li, L.; Xie, L.; Chen, M.; Shen, Q.; Pan, Q.; Fu, X.; Shi, P.; Tang, Y.; Huang, H.; et al. A Novel HD-ZIP IV/MIXTA Complex Promotes Glandular Trichome Initiation and Cuticle Development in Artemisia annua. New Phytol. 2018, 218, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Glover, B.J.; Perez-Rodriguez, M.; Martin, C. Development of Several Epidermal Cell Types Can Be Specified by the Same MYB-Related Plant Transcription Factor. Development 1998, 125, 3497–3508. [Google Scholar] [CrossRef] [PubMed]
- Apicella, P.V.; Sands, L.B.; Ma, Y.; Berkowitz, G.A. Delineating the Genetic Regulation of Cannabinoid Biosynthesis during Female Flower Development in Cannabis sativa. Plant Direct, 2022; accepted. [Google Scholar]
- Deguchi, M.; Bogush, D.; Weeden, H.; Spuhler, Z.; Potlakayala, S.; Kondo, T.; Zhang, Z.J.; Rudrabhatla, S. Establishment and Optimization of a Hemp (Cannabis sativa L.) Agroinfiltration System for Gene Expression and Silencing Studies. Sci. Rep. 2020, 10, 3504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galán-Ávila, A.; Gramazio, P.; Ron, M.; Prohens, J.; Herraiz, F.J. A Novel and Rapid Method for Agrobacterium-Mediated Production of Stably Transformed Cannabis sativa L. Plants. Ind. Crops Prod. 2021, 170, 113691. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, G.; Cheng, C.; Lei, L.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Establishment of an Agrobacterium -mediated Genetic Transformation and CRISPR/Cas9-mediated Targeted Mutagenesis in Hemp (Cannabis Sativa L.). Plant Biotechnol. J 2021, 19, 1979–1987. [Google Scholar] [CrossRef]
- Uzelac, B.; Stojičić, D.; Budimir, S. Glandular Trichomes on the Leaves of Nicotiana tabacum: Morphology, Developmental Ultrastructure, and Secondary Metabolites. In Plant Cell and Tissue Differentiation and Secondary Metabolites; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Reference Series in Phytochemistry; Springer International Publishing: Cham, Switzerland, 2021; pp. 25–61. ISBN 978-3-030-30184-2. [Google Scholar]
- Lashbrooke, J.G.; Adato, A.; Lotan, O.; Alkan, N.; Tsimbalist, T.; Rechav, K.; Fernandez Moreno, J.-P.; Widemann, E.; Grausem, B.; Pinot, F.; et al. The Tomato MIXTA-like Transcription Factor Coordinates Fruit Epidermis Conical Cell Development and Cuticular Lipid Biosynthesis and Assembly. Plant Physiol. 2015, 169, 2553–2571. [Google Scholar] [CrossRef]
- Oshima, Y.; Shikata, M.; Koyama, T.; Ohtsubo, N.; Mitsuda, N.; Ohme-Takagi, M. MIXTA-Like Transcription Factors and WAX INDUCER1/SHINE1 Coordinately Regulate Cuticle Development in Arabidopsis and Torenia Fournieri. Plant Cell 2013, 25, 1609–1624. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Taylor, L.; Pucker, B.; Feng, T.; Glover, B.J.; Brockington, S.F. The Land Plant-specific MIXTA-MYB Lineage Is Implicated in the Early Evolution of the Plant Cuticle and the Colonization of Land. New Phytol. 2021, 229, 2324–2338. [Google Scholar] [CrossRef]
- Payne, T.; Clement, J.; Arnold, D.; Lloyd, A. Heterologous Myb Genes Distinct from GL1 Enhance Trichome Production When Overexpressed in Nicotiana Tabacum. Development 1999, 126, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Guo, H.; Zhang, Q.; Guo, M.; Xu, Y.; Zeng, M.; Lv, P.; Chen, X.; Yang, M. Evaluation of Reference Genes for RT-QPCR Analysis in Wild and Cultivated Cannabis. Biosci. Biotechnol. Biochem. 2018, 82, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Rivas, S.; Mestre, P.; Baulcombe, D. Retracted: An Enhanced Transient Expression System in Plants Based on Suppression of Gene Silencing by the P19 Protein of Tomato Bushy Stunt Virus: An Enhanced Transient Expression System in N. Benthamiana. Plant J. 2003, 33, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haiden, S.R.; Apicella, P.V.; Ma, Y.; Berkowitz, G.A. Overexpression of CsMIXTA, a Transcription Factor from Cannabis sativa, Increases Glandular Trichome Density in Tobacco Leaves. Plants 2022, 11, 1519. https://doi.org/10.3390/plants11111519
Haiden SR, Apicella PV, Ma Y, Berkowitz GA. Overexpression of CsMIXTA, a Transcription Factor from Cannabis sativa, Increases Glandular Trichome Density in Tobacco Leaves. Plants. 2022; 11(11):1519. https://doi.org/10.3390/plants11111519
Chicago/Turabian StyleHaiden, Samuel R., Peter V. Apicella, Yi Ma, and Gerald A. Berkowitz. 2022. "Overexpression of CsMIXTA, a Transcription Factor from Cannabis sativa, Increases Glandular Trichome Density in Tobacco Leaves" Plants 11, no. 11: 1519. https://doi.org/10.3390/plants11111519