
Ascorbic Acid Preconditioning Effect on Broccoli Seedling Growth and Photosynthesis under Drought Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
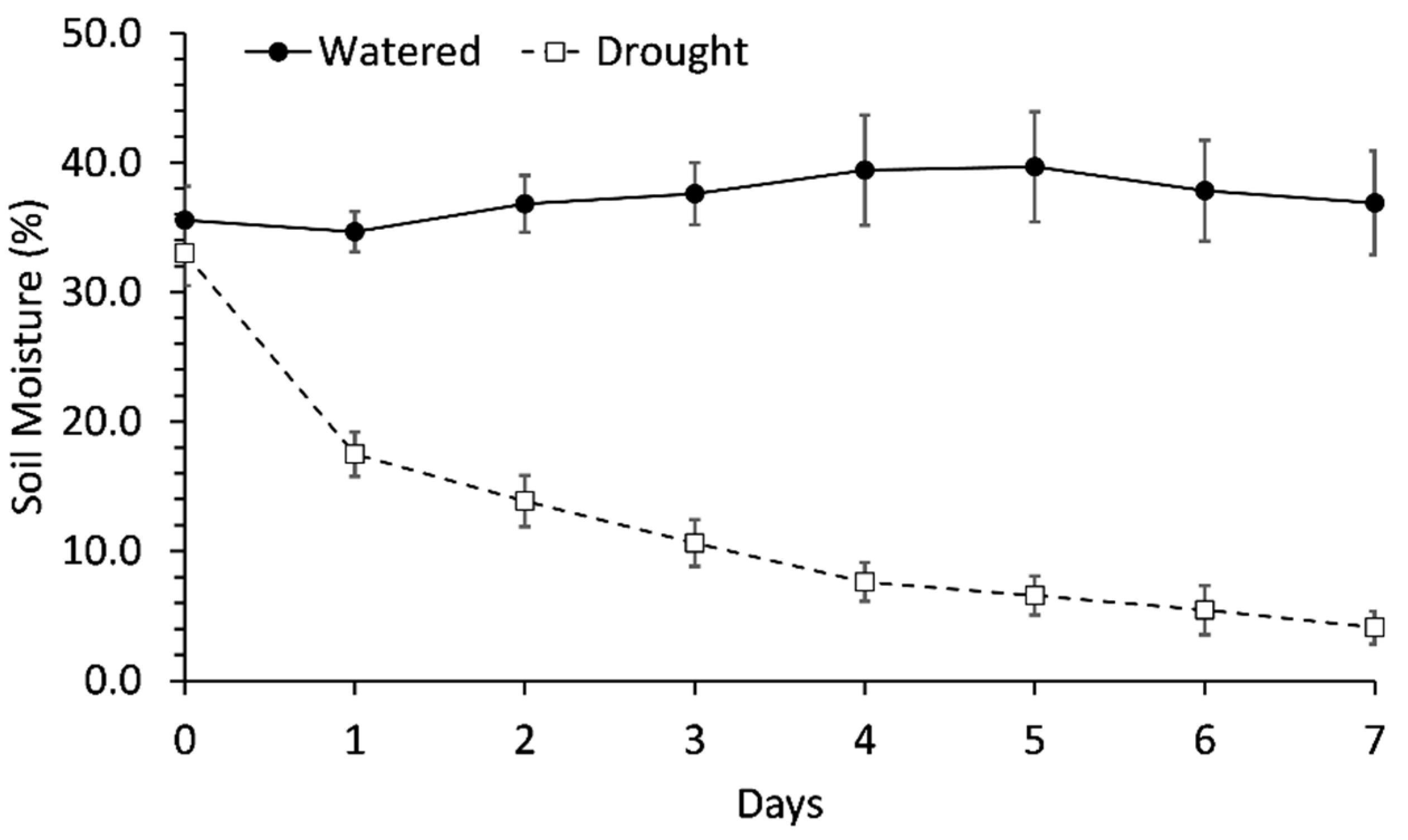
2.1. Soil Moisture
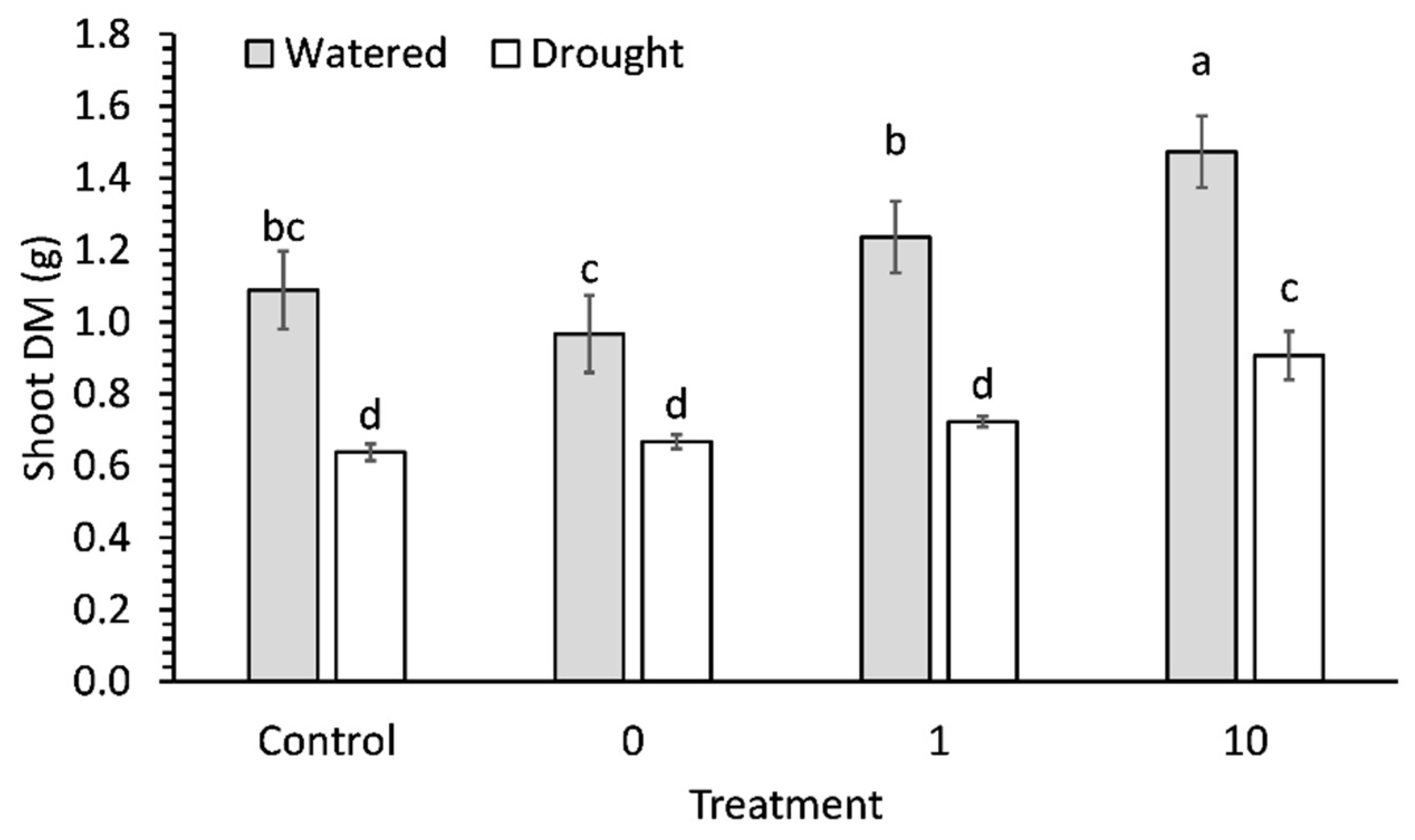
2.2. Biomass
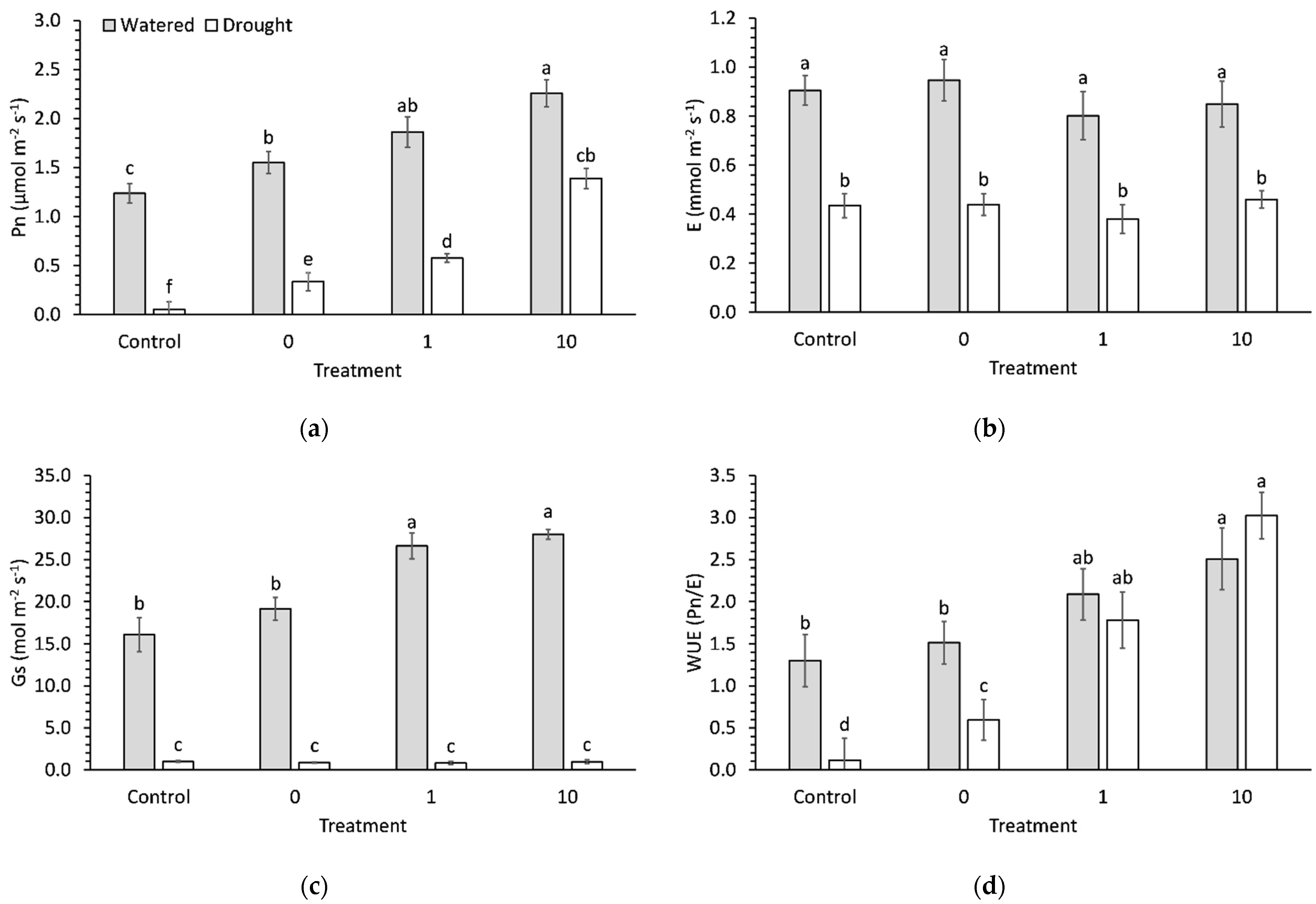
2.3. Photosynthetic Measurements
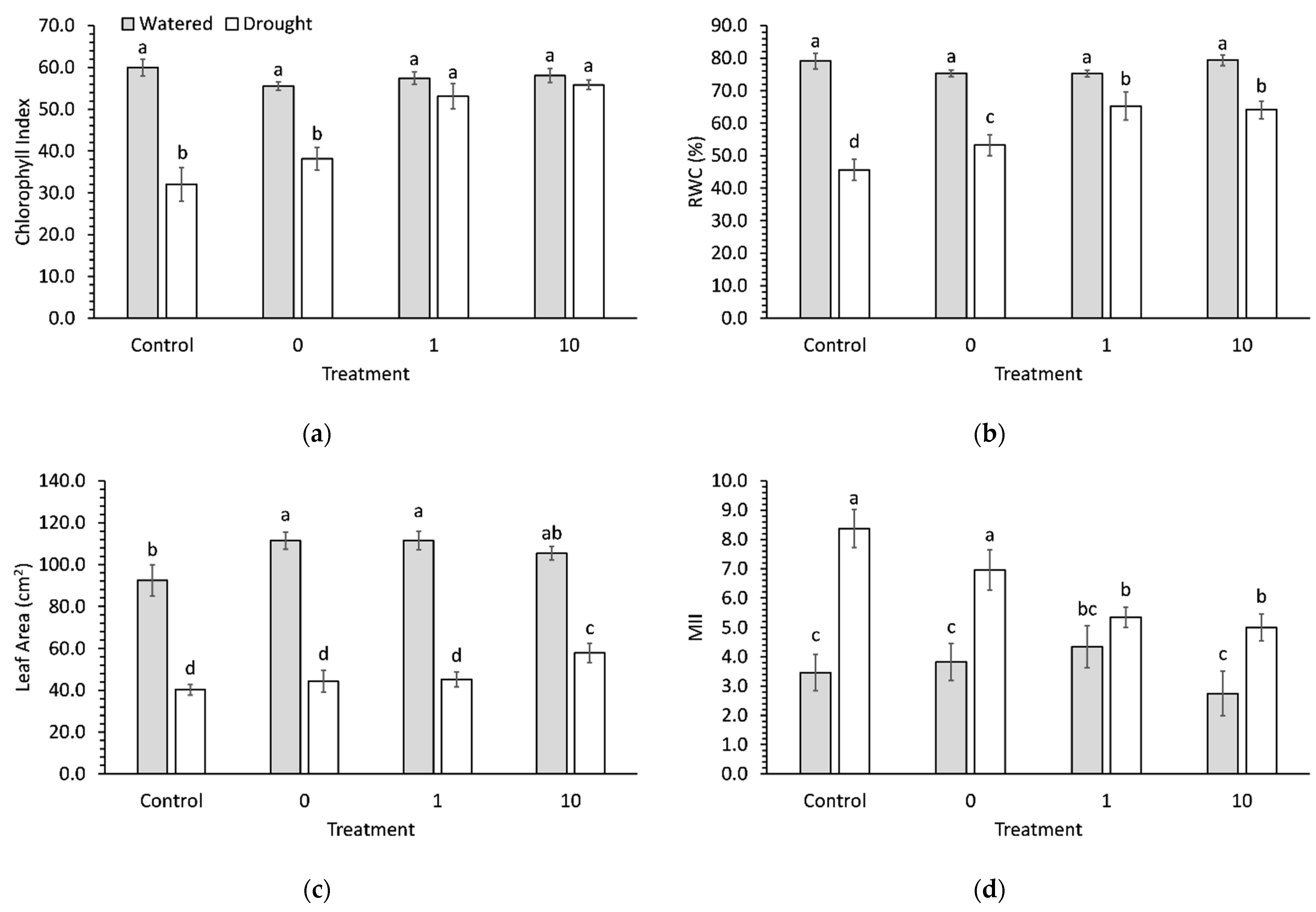
2.4. Leaf Measurements
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Seed Preconditioning
4.3. Growing Conditions
4.4. Photosynthetic Measurements
4.5. Intact Leaf Measurements
4.6. Detached Leaf Measurements
4.7. Biomass Measurements
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diaz-Perez, J.C. Root Zone Temperature, Plant Growth and Yield of Broccoli [Brassica oleracea (Plenck) var. italica as Affected by Film Mulches. Sci. Hortic. 2009, 123, 156–163. [Google Scholar] [CrossRef]
- Ilahy, R.; Tlili, I.; Pék, Z.; Montefusco, A.; Siddiqui, M.W.; Homa, F.; Hdider, C.; R’Him, T.; Lajos, H.; Lenucci, M.S. Pre- and Post-Harvest Factors Affecting Glucosinolate Content in Broccoli. Front. Nutr. 2020, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W. Brassica: Characteristics and Properties. In Encyclopedia of Food and Health; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 469–477. [Google Scholar]
- Osman, A.S.; Wahed, M.H.A.; Rady, M.M. Ascorbic Acid Improves Productivity, Physio Biochemical Attributes and Antioxidant Activity of Deficit Irrigated Broccoli Plants. Biomed. J. Sci. Tech. Res. 2018, 11, 8196–8205. [Google Scholar] [CrossRef]
- Beacham, A.M.; Hand, P.; Pink, D.A.; Monaghan, J.M. Analysis of Brassica Oleracea Early Stage Abiotic Stress Responses Reveals Tolerance in Multiple Crop Types and for Multiple Sources of Stress. J. Sci. Food Agric. 2017, 97, 5271–5277. [Google Scholar] [CrossRef]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.M.; Nayyar, H. Individual and Combined Effects of Transient Drought and Heat Stress on Carbon Assimilation and Seed Filling in Chickpea. Funct. Plant Biol. FPB 2014, 41, 1148–1167. [Google Scholar] [CrossRef] [Green Version]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory Metabolic Networks in Drought Stress Responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Vadez, V.; Berger, J.D.; Warkentin, T.; Asseng, S.; Ratnakumar, P.; Rao, K.P.C.; Gaur, P.M.; Munier-Jolain, N.; Larmure, A.; Voisin, A.-S.; et al. Adaptation of Grain Legumes to Climate Change: A Review. Agron. Sustain. Dev. 2012, 32, 31–44. [Google Scholar] [CrossRef]
- Galmés, J.; Medrano, H.; Flexas, J. Photosynthetic Limitations in Response to Water Stress and Recovery in Mediterranean Plants with Different Growth Forms. New Phytol. 2007, 175, 81–93. [Google Scholar] [CrossRef]
- Seminario, A.; Song, L.; Zulet, A.; Nguyen, H.T.; González, E.M.; Larrainzar, E. Drought Stress Causes a Reduction in the Biosynthesis of Ascorbic Acid in Soybean Plants. Front. Plant Sci. 2017, 8, 1042. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Liu, H.; Sultan, M.A.R.F.; Liu, X.L.; Zhang, J.; Yu, F.; Zhao, H.X. Physiological and Comparative Proteomic Analysis Reveals Different Drought Responses in Roots and Leaves of Drought-Tolerant Wild Wheat (Triticum boeoticum). PLoS ONE 2015, 10, e0121852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and Biochemical Changes during Drought and Recovery Periods at Tillering and Jointing Stages in Wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of Drought Stress on Photosynthetic Response, Leaf Water Potential, and Stem Sap Flow in Two Cultivars of Bi-Leader Apple Trees (Malus domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of Morphological, Physiological, and Biochemical Traits for Assessing Drought Resistance in Eleven Tree Species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, J.S.; Tyagi, M.K.; Kumar, A.; Nashaat, N.I.; Singh, M.; Singh, N.B.; Jakhar, M.L.; Welham, S.J. Drought Effects on Yield and Its Components in Indian Mustard (Brassica juncea L.). Plant Breed. 2007, 126, 399–402. [Google Scholar] [CrossRef]
- Sabagh, A.E.; Hossain, A.; Barutcular, C.; Islam, M.S.; Ratnasekera, D.; Kumar, N.; Meena, R.S.; Gharib, H.S.; Saneoka, H.; da Silva, J.A.T. Drought and Salinity Stress Management for Higher and Sustainable Canola (“Brassica Napus” L.) Production: A Critical Review. Aust. J. Crop Sci. 2019, 13, 88–96. [Google Scholar] [CrossRef]
- IBU Repository. Effects of Water Stress on Yield and Some Quality Parameters of Broccoli. Available online: https://eprints.ibu.edu.ba/items/show/3026 (accessed on 3 May 2022).
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of Extreme Weather Disasters on Global Crop Production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Kim, W.; Iizumi, T.; Nishimori, M. Global Patterns of Crop Production Losses Associated with Droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar] [CrossRef]
- Rajasekaran, B. New Plant Growth Regulators Protect Photosynthesis and Enhance Growth Under Drought of Jack Pine Seedlings. J. Plant Growth Regul. 1999, 18, 175–181. [Google Scholar] [CrossRef]
- Lada, R.R.; Stiles, A.; Blake, T.J. The Effects of Natural and Synthetic Seed Preconditioning Agents (SPAs) in Hastening Seedling Emergence and Enhancing Yield and Quality of Processing Carrots. Sci. Hortic. 2005, 106, 25–37. [Google Scholar] [CrossRef]
- MacDonald, M.T.; Lada, R.R.; Robinson, A.R.; Hoyle, J. The Benefits of Ambiol® in Promoting Germination, Growth, and Drought Tolerance Can Be Passed on to Next-Generation Tomato Seedlings. J. Plant Growth Regul. 2010, 29, 357–365. [Google Scholar] [CrossRef]
- Simma, B.; Polthanee, A.; Goggi, A.S.; Siri, B.; Promkhambut, A.; Caragea, P.C. Wood Vinegar Seed Priming Improves Yield and Suppresses Weeds in Dryland Direct-Seeding Rice under Rainfed Production. Agron. Sustain. Dev. 2017, 37, 56. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, M.T.; Lada, R.R.; Robinson, A.R.; Hoyle, J. Seed Preconditioning with Natural and Synthetic Antioxidants Induces Drought Tolerance in Tomato Seedlings. HortScience 2009, 44, 1323–1329. [Google Scholar] [CrossRef] [Green Version]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic Acid Priming Enhances Seed Germination and Seedling Growth of Winter Wheat under Low Temperature Due to Late Sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Anwar, S.; Iqbal, M.; Raza, S.H.; Iqbal, N. Efficacy of Seed Preconditioning with Salicylic and Ascorbic Acid in Increasing Vigor of Ruce (Oryza sativa L.) Seedling. Pak. J. Bot. 2013, 45, 157–162. [Google Scholar]
- Bhusal, N.; Lee, M.; Reum Han, A.; Han, A.; Kim, H.S. Responses to Drought Stress in Prunus Sargentii and Larix Kaempferi Seedlings Using Morphological and Physiological Parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Jo, H.; Chang, H.; An, J.; Cho, M.S.; Son, Y. Species Specific Physiological Responses of Pinus Densiflora and Larix Kaempferi Seedlings to Open-Field Experimental Warming and Precipitation Manipulation. For. Sci. Technol. 2019, 15, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Cao, X.; Niu, J. Effects of Exogenous Ascorbic Acid on Seed Germination and Seedling Salt-Tolerance of Alfalfa. PLoS ONE 2021, 16, e0250926. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.T.; Lada, R.R.; Veitch, R.S.; Thiagarajan, A.; Adams, A.D. Postharvest Needle Abscission Resistance of Balsam Fir (Abies balsamea) is Modified by Harvest Date. Can. J. For. Res. 2014, 44, 1394–1401. [Google Scholar] [CrossRef]
- Jamil, S.; Ali, Q.; Iqbal, M.; Javed, M.T.; Iftikhar, W.; Shahzad, F.; Perveen, R. Modulations in Plant Water Relations and Tissue-Specific Osmoregulation by Foliar-Applied Ascorbic Acid and the Induction of Salt Tolerance in Maize Plants. Braz. J. Bot. 2015, 38, 527–538. [Google Scholar] [CrossRef]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Zulqurnain Haider, M.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-Daim, M.M.; Elkelish, A.; et al. Use of Nitric Oxide and Hydrogen Peroxide for Better Yield of Wheat (Triticum aestivum L.) under Water Deficit Conditions: Growth, Osmoregulation, and Antioxidative Defense Mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Nephali, L.; Moodley, V.; Piater, L.; Steenkamp, P.; Buthelezi, N.; Dubery, I.; Burgess, K.; Huyser, J.; Tugizimana, F. A Metabolomic Landscape of Maize Plants Treated with a Microbial Biostimulant Under Well-Watered and Drought Conditions. Front. Plant Sci. 2021, 12, 676632. [Google Scholar] [CrossRef] [PubMed]
- Martínez, D.E.; Guiamet, J.J. Distortion of the SPAD 502 Chlorophyll Meter Readings by Changes in Irradiance and Leaf Water Status. Agronomie 2004, 24, 41–46. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, M.T.; Lada, R.R. Biophysical and Hormonal Changes Linked to Postharvest Needle Abscission in Balsam Fir. J. Plant Growth Regul. 2014, 33, 602–611. [Google Scholar] [CrossRef]
- Odlum, K.D.; Blake, T.J. A Comparison of Analytical Approaches for Assessing Freezing Damage in Black Spruce Using Electrolyte Leakage Methods. Can. J. Bot. 1996, 74, 952–958. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacDonald, M.T.; Kannan, R.; Jayaseelan, R. Ascorbic Acid Preconditioning Effect on Broccoli Seedling Growth and Photosynthesis under Drought Stress. Plants 2022, 11, 1324. https://doi.org/10.3390/plants11101324
MacDonald MT, Kannan R, Jayaseelan R. Ascorbic Acid Preconditioning Effect on Broccoli Seedling Growth and Photosynthesis under Drought Stress. Plants. 2022; 11(10):1324. https://doi.org/10.3390/plants11101324
Chicago/Turabian StyleMacDonald, Mason T., Rajeswari Kannan, and Renuga Jayaseelan. 2022. "Ascorbic Acid Preconditioning Effect on Broccoli Seedling Growth and Photosynthesis under Drought Stress" Plants 11, no. 10: 1324. https://doi.org/10.3390/plants11101324