
Late Blight Resistance Conferred by Rpi-Smira2/R8 in Potato Genotypes In Vitro Depends on the Genetic Background

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Marker-Assisted Selection and Avr4 efector Agroinfiltration of Progeny Potato Genotypes Carrying Rpi-Smira2/R8 Gene
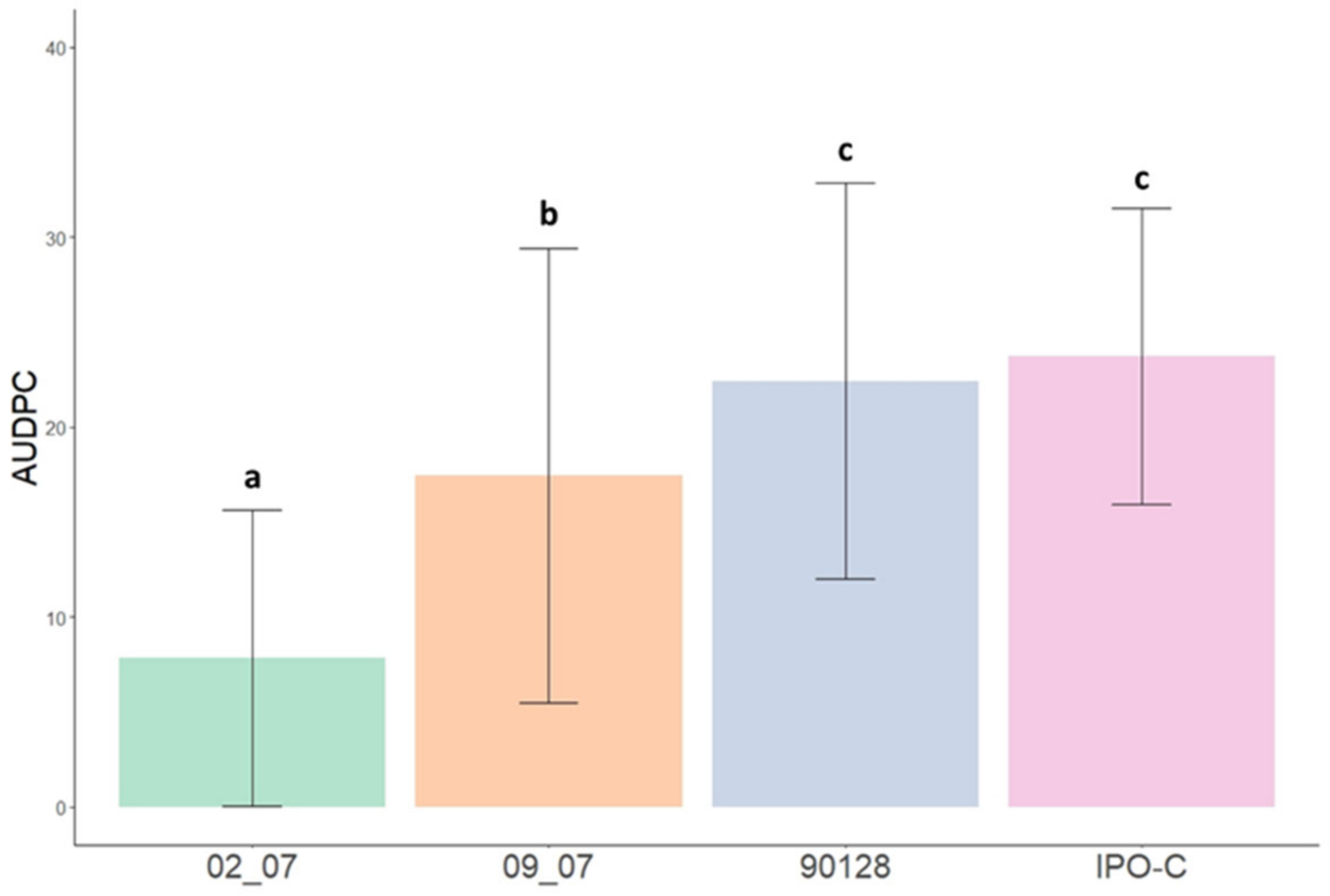
2.2. Disease Progression of Phytophthora infestans Isolates
2.3. Late Blight Resistance of Progeny R8 Genotypes
3. Discussion
3.1. Combining Available PCR-Based Markers for Late Blight Resistance and Avr4 Effector Gene Agroinfiltration Enabled the Selection of Potato Plantlets Carrying Solely Rpi-Smira2/R8 Gene
3.2. Slovenian Isolate 02_07 Was the Least Aggressive Isolate despite Belonging to EU_13_A2 Genotype
3.3. Rpi-Smira2/R8 Gene Is Capable of Conferring Resistance to Late Blight Comparable to Sárpo Mira, Although This Trait Depends on the Genetic Background
4. Materials and Methods
4.1. Plant Material
4.2. Marker-Assisted Selection (MAS) of Potato Genotypes
4.3. Selection of R4-Negative Plants Using Avr4 Effector Gene Agroinfiltration
4.4. Maintenance of Phytophthora infestans Isolates and Inoculum Preparation
4.5. In Vitro Inoculation of R8 Plants with Phytophthora infestans Zoospores
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Production. Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 6 April 2022).
- EUROSTAT Statistics Explained. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=The_EU_potato_sector_-_statistics_on_production,_prices_and_trade#Production:_area.2C_harvest_and_farms (accessed on 6 April 2022).
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five Reasons to Consider Phytophthora infestans a Reemerging Pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haverkort, A.J.; Struik, P.C.; Visser, R.G.F.; Jacobsen, E. Applied Biotechnology to Combat Late Blight in Potato Caused by Phytophthora infestans. Potato Res. 2009, 52, 249–264. [Google Scholar] [CrossRef]
- Ivanov, A.A.; Ukladov, E.O.; Golubeva, T.S. Phytophthora infestans: An Overview of Methods and Attempts to Combat Late Blight. J. Fungi 2021, 7, 1071. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, A.P.; Vega-Arreguín, J.C.; Fei, Z.; Ponnala, L.; Lee, S.J.; Matas, A.J.; Patev, S.; Fry, W.E.; Rose, J.K.C. Transcriptional Dynamics of Phytophthora infestans during Sequential Stages of Hemibiotrophic Infection of Tomato. Mol. Plant Pathol. 2016, 17, 29–41. [Google Scholar] [CrossRef]
- Galindo, A.J.; Gallegly, M.E. The Nature of Sexuality in Phytophthora infestans. Phytopathology 1960, 50, 123–128. [Google Scholar]
- Fawke, S.; Doumane, M.; Schornack, S. Oomycete Interactions with Plants: Infection Strategies and Resistance Principles. Microbiol. Mol. Biol. Rev. 2015, 79, 263–280. [Google Scholar] [CrossRef] [Green Version]
- Mazáková, J.; Táborský, V.; Zouhar, M.; Ryšánek, P.; Hausvater, E.; Doležal, P. Occurrence and Distribution of Mating Types A1 and A2 of Phytophthora infestans (Mont.) de Bary in the Czech Republic. Plant Prot. Sci. 2018, 42, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Drenth, A.; Turkensteen, L.J.; Govers, F. The Occurrence of the A2 Mating Type of Phytophthora infestans in the Netherlands; Significance and Consequences. Neth. J. Plant Pathol. 1993, 99, 57–67. [Google Scholar] [CrossRef]
- Fry, W.E. Phytophthora infestans: New Tools (and Old Ones) Lead to New Understanding and Precision Management. Annu. Rev. Phytopathol. 2016, 54, 529–547. [Google Scholar] [CrossRef]
- EuroBlight Genotype Frequency Chart. Available online: https://agro.au.dk/forskning/internationale-platforme/euroblight/pathogen-monitoring/genotype-frequency-chart (accessed on 3 May 2022).
- Cooke, D.E.L.; Cano, L.M.; Raffaele, S.; Bain, R.A.; Cooke, L.R.; Etherington, G.J.; Deahl, K.L.; Farrer, R.A.; Gilroy, E.M.; Goss, E.M.; et al. Genome Analyses of an Aggressive and Invasive Lineage of the Irish Potato Famine Pathogen. PLoS Pathog. 2012, 8, e1002940. [Google Scholar] [CrossRef]
- EuroBlight Results of the EuroBlight Potato Late Blight Monitoring in 2020. Available online: https://agro.au.dk/forskning/internationale-platforme/euroblight/currently/news/nyhed/artikel/results-of-the-euroblight-potato-late-blight-monitoring-in-2020 (accessed on 6 April 2022).
- Jo, K.R.; Kim, C.J.; Kim, S.J.; Kim, T.Y.; Bergervoet, M.; Jongsma, M.A.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Development of Late Blight Resistant Potatoes by Cisgene Stacking. BMC Biotechnol. 2014, 14, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beketova, M.P.; Chalaya, N.A.; Zoteyeva, N.M.; Gurina, A.A.; Kuznetsova, M.A.; Armstrong, M.; Hein, I.; Drobyazina, P.E.; Khavkin, E.E.; Rogozina, E.V. Combination Breeding and Marker-Assisted Selection to Develop Late Blight Resistant Potato Cultivars. Agronomy 2021, 11, 2192. [Google Scholar] [CrossRef]
- Zimnoch-Guzowska, E.; Flis, B. Over 50 Years of Potato Parental Line Breeding Programme at the Plant Breeding and Acclimatization Institute in Poland. Potato Res. 2021, 64, 743–760. [Google Scholar] [CrossRef]
- Keijzer, P.; van Bueren, E.T.L.; Engelen, C.J.M.; Hutten, R.C.B. Breeding Late Blight Resistant Potatoes for Organic Farming—A Collaborative Model of Participatory Plant Breeding: The Bioimpuls Project. Potato Res. 2021, 65, 349–377. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Thieme, R.; König, J.; Nachtigall, M.; Hammann, T.; Denes, T.E.; Kruppa, K.; Molnár-Láng, M. Introgression of Two Broad-Spectrum Late Blight Resistance Genes, Rpi-Blb1 and Rpi-Blb3, From Solanum Bulbocastanum Dun Plus Race-Specific R Genes into Potato Pre-Breeding Lines. Front. Plant Sci. 2020, 11, 699. [Google Scholar] [CrossRef]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and Tomato Late Blight Caused by Phytophthora infestans: An over View of Pathology and Resistance Breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Flor, H.H. Current Status of the Gene-Fob-Gene Concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Du, J.; Rietman, H.; Vleeshouwers, V.G.A.A. Agroinfiltration and PVX Agroinfection in Potato and Nicotiana Benthamiana. J. Vis. Exp. 2014, 83, e50971. [Google Scholar] [CrossRef] [Green Version]
- Mba’u John, Y.; Faizal, A. Transient Transformation of Potato Plant (Solanum Tuberosum L.) Granola Cultivar Using Syringe Agroinfiltration. J. Agric. Sci. 2018, 40, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Vleeshouwers, V.G.A.A.; Van Dooijeweert, W.; Govers, F.; Kamoun, S.; Colon, L.T. The Hypersensitive Response is Associated with Host and Nonhost Resistance to Phytophthora infestans. Planta 2000, 210, 853–864. [Google Scholar] [CrossRef]
- Bhaskar, P.B.; Venkateshwaran, M.; Wu, L.; Ané, J.M.; Jiang, J. Agrobacterium-Mediated Transient Gene Expression and Silencing: A Rapid Tool for Functional Gene Assay in Potato. PLoS ONE 2009, 4, e5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.K.; Prat, S.; Hannapel, D.J. Efficient Production of Transgenic Potato (S. Tuberosum L. Ssp. Andigena) Plants via Agrobacterium Tumefaciens-Mediated Transformation. Plant Sci. 2006, 170, 732–738. [Google Scholar] [CrossRef]
- Heeres, P.; Schippers-Rozenboom, M.; Jacobsen, E.; Visser, R.G.F. Transformation of a Large Number of Potato Varieties: Genotype-Dependent Variation in Efficiency and Somaclonal Variability. Euphytica 2002, 124, 13–22. [Google Scholar] [CrossRef]
- Serra, O.; Soler, M.; Hohn, C.; Sauveplane, V.; Pinot, F.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. CYP86A33-Targeted Gene Silencing in Potato Tuber Alters Suberin Composition, Distorts Suberin Lamellae, and Impairs the Periderm’s Water Barrier Function. Plant Physiol. 2009, 149, 1050–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, T.H.; Vleeshouwers, V.G.A.A.; Jacobsen, E.; Van Der Vossen, E.; Visser, R.G.F. Molecular Breeding for Resistance to Phytophthora infestans (Mont.) de Bary in Potato (Solanum tuberosum L.): A Perspective of Cisgenesis. Plant Breed. 2009, 128, 109–117. [Google Scholar] [CrossRef]
- Veale, M.A.; Slabbert, M.M.; Van Emmenes, L. Agrobacterium-Mediated Transformation of Potato Cv. Mnandi for Resistance to the Potato Tuber Moth (Phthorimaea Operculella). S. Afr. J. Bot. 2012, 80, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Vleeshouwers, V.G.A.A.; Van Dooijeweert, W.; Keizer, L.C.P.; Sijpkes, L.; Govers, F.; Colon, L.T. A Laboratory Assay for Phytophthora infestans Resistance in Various Solanum Species Reflects the Field Situation. Eur. J. Plant Pathol. 1999, 105, 241–250. [Google Scholar] [CrossRef]
- Turkensteen, L.J. Durable Resistance of Potatoes against Phytophthora infestans. In Durability of Disease Resistance; Jacobs, T., Parlevliet, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993; pp. 115–124. [Google Scholar]
- Rodewald, J.; Trognitz, B. Solanum Resistance Genes against Phytophthora infestans and Their Corresponding Avirulence Genes. Mol. Plant Pathol. 2013, 14, 740–757. [Google Scholar] [CrossRef]
- Witek, K.; Lin, X.; Karki, H.S.; Jupe, F.; Witek, A.I.; Steuernagel, B.; Stam, R.; van Oosterhout, C.; Fairhead, S.; Heal, R.; et al. A Complex Resistance Locus in Solanum Americanum Recognizes a Conserved Phytophthora Effector. Nat. Plants 2021, 7, 198–208. [Google Scholar] [CrossRef]
- White, S.; Shaw, D. The Usefulness of Late-Blight Resistant Sárpo Cultivars-a Case Study. Acta Hort. 2009, 834, 161–166. [Google Scholar] [CrossRef]
- Tomczyńska, I.; Stefańczyk, E.; Chmielarz, M.; Karasiewicz, B.; Kamiński, P.; Jones, J.D.G.; Lees, A.K.; Śliwka, J. A Locus Conferring Effective Late Blight Resistance in Potato Cultivar Sárpo Mira Maps to Chromosome XI. Theor. Appl. Genet. 2014, 127, 647–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.; Shaw, D. Breeding for Host Resistance: The Key to Sustainable Potato Production. PPO-Spec. Rep. 2010, 14, 125–132. [Google Scholar]
- Lees, A.K.; Stewart, J.A.; Lynott, J.S.; Carnegie, S.F.; Campbell, H.; Roberts, A.M.I. The Effect of a Dominant Phytophthora infestans Genotype (13_A2) in Great Britain on Host Resistance to Foliar Late Blight in Commercial Potato Cultivars. Potato Res. 2012, 55, 125–134. [Google Scholar] [CrossRef]
- Rietman, H.; Bijsterbosch, G.; Cano, L.M.; Lee, H.-R.; Vossen, J.H.; Jacobsen, E.; Visser, R.G.F.; Kamoun, S.; Vleeshouwers, V.G.A.A. Qualitative and Quantitative Late Blight Resistance in the Potato Cultivar Sarpo Mira Is Determined by the Perception of Five Distinct RXLR Effectors. Mol. Plant-Microbe Interact. 2012, 25, 910–919. [Google Scholar] [CrossRef] [Green Version]
- Rietman, H. Putting the Phytophthora infestans Genome Sequence at Work; Multiple Novel Avirulence and Potato Resistance Gene Candidates Revealed. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 20 June 2011. [Google Scholar]
- Orłowska, E.; Fiil, A.; Kirk, H.G.; Llorente, B.; Cvitanich, C. Differential Gene Induction in Resistant and Susceptible Potato Cultivars at Early Stages of Infection by Phytophthora infestans. Plant Cell Rep. 2012, 31, 187–203. [Google Scholar] [CrossRef]
- Orłowska, E.; Basile, A.; Kandzia, I.; Llorente, B.; Kirk, H.G.; Cvitanich, C. Revealing the Importance of Meristems and Roots for the Development of Hypersensitive Responses and Full Foliar Resistance to Phytophthora infestans in the Resistant Potato Cultivar Sarpo Mira. J. Exp. Bot. 2012, 63, 4765–4779. [Google Scholar] [CrossRef]
- Stefańczyk, E.; Sobkowiak, S.; Brylińska, M.; Śliwka, J. Expression of the Potato Late Blight Resistance Gene Rpi-Phu1 and Phytophthora infestans Effectors in the Compatible and Incompatible Interactions in Potato. Phytopathology 2017, 107, 740–748. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.; Cao, X.; Zhou, X.; Chen, Z.; Wang, Q.; Liu, W.; Chen, Q.; Zhao, H. The Histological, Effectoromic, and Transcriptomic Analyses of Solanum Pinnatisectum Reveal an Upregulation of Multiple NBS-LRR Genes Suppressing Phytophthora infestans Infection. Int. J. Mol. Sci. 2020, 21, 3211. [Google Scholar] [CrossRef]
- Jiang, R.; Li, J.; Tian, Z.; Du, J.; Armstrong, M.; Baker, K.; Tze-Yin Lim, J.; Vossen, J.H.; He, H.; Portal, L.; et al. Potato Late Blight Field Resistance from QTL DPI09c Is Conferred by the NB-LRR Gene R8. J. Exp. Bot. 2018, 69, 1545–1555. [Google Scholar] [CrossRef]
- Jo, K.R.; Arens, M.; Kim, T.Y.; Jongsma, M.A.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Mapping of the S. Demissum Late Blight Resistance Gene R8 to a New Locus on Chromosome IX. Theor. Appl. Genet. 2011, 123, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Mahfouze, H.A.; El-Sayed, O.E. Comparative Analysis of Late Blight Resistance R Genes and Their Coding Proteins in Some Potato Genotypes. Czech J. Genet. Plant Breed. 2022, 58, 10–20. [Google Scholar] [CrossRef]
- Jo, K.-R. Unveiling and Deploying Durability of Late Blight Resistance in Potato; From Natural Stacking to Cisgenic Stacking. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 17 September 2013. [Google Scholar]
- Huang, S. Discovery and Characterization of the Major Late Blight Resistance Complex in Potato. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 31 January 2005. [Google Scholar]
- Vossen, J.H.; van Arkel, G.; Bergervoet, M.; Jo, K.R.; Jacobsen, E.; Visser, R.G.F. The Solanum Demissum R8 Late Blight Resistance Gene is an Sw-5 Homologue That Has Been Deployed Worldwide in Late Blight Resistant Varieties. Theor. Appl. Genet. 2016, 129, 1785–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Lee, H.R.; Jo, K.R.; Mortazavian, S.M.M.; Huigen, D.J.; Evenhuis, B.; Kessel, G.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Broad Spectrum Late Blight Resistance in Potato Differential Set Plants MaR8 and MaR9 Is Conferred by Multiple Stacked R Genes. Theor. Appl. Genet. 2012, 124, 923–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Poppel, P.M.J.A. The Phytophthora infestans Avirulence Gene PiAvr4 and Its Potato Counterpart R4. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 4 February 2009. [Google Scholar]
- Huang, S.; Vleeshouwers, V.G.A.A.; Visser, R.G.F.; Jacobsen, E. An Accurate in Vitro Assay for High-Throughput Disease Testing of Phytophthora infestans in Potato. Plant Dis. 2005, 89, 1263–1267. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Van Der Vossen, E.A.G.; Kuang, H.; Vleeshouwers, V.G.A.A.; Zhang, N.; Borm, T.J.A.; Van Eck, H.J.; Baker, B.; Jacobsen, E.; Visser, R.G.F. Comparative Genomics Enabled the Isolation of the R3a Late Blight Resistance Gene in Potato. Plant J. 2005, 42, 251–261. [Google Scholar] [CrossRef]
- Mirkarimi, H.R.; Abasi-moghadam, A.; Mozafari, J. In Vitro and Greenhouse Evaluation for Resistance to Early Blight of Potato Isolated from Alternaria Alternata. Agric. Sci. 2013, 4, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jung, C.S.; De Jong, W.S. Genetic Analysis of Pigmented Tuber Flesh in Potato. Theor. Appl. Genet. 2009, 119, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, C.; Valkonen, J.P.T. Organization of Genes Controlling Disease Resistance in The Potato Genome. Annu. Rev. Phytopathol. 2001, 39, 79–102. [Google Scholar] [CrossRef]
- Van Poppel, P.M.J.A.; Huigen, D.J.; Govers, F. Differential Recognition of Phytophthora infestans Races in Potato R4 Breeding Lines. Phytopathology 2009, 99, 1150–1155. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Vleeshouwers, V.G.A.A.; Werij, J.S.; Hutten, R.C.B.; Van Eck, H.J.; Visser, R.G.F.; Jacobsen, E. The R3 Resistance to Phytophthora infestans in Potato Is Conferred by Two Closely Linked R Genes with Distinct Specificities. Mol. Plant-Microbe Interact. 2004, 17, 428–435. [Google Scholar] [CrossRef] [Green Version]
- Jo, K.R.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Characterisation of the Late Blight Resistance in Potato Differential MaR9 Reveals a Qualitative Resistance Gene, R9a, Residing in a Cluster of Tm-2 2 Homologs on Chromosome IX. Theor. Appl. Genet. 2015, 128, 931–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žerjav, M. Značilnosti Populacije Krompirjeve Plesni (Phytophthora infestans (Mont.) de Bary) v Sloveniji v Obdobju Med 2002 in 2015. Master’s Thesis, Univerza v Ljubljani, Ljubljana, Slovenia, 4 May 2016. [Google Scholar]
- Najdabbasi, N.; Mirmajlessi, S.M.; Dewitte, K.; Ameye, M.; Mänd, M.; Audenaert, K.; Landschoot, S.; Haesaert, G. Green Leaf Volatile Confers Management of Late Blight Disease: A Green Vaccination in Potato. J. Fungi 2021, 7, 312. [Google Scholar] [CrossRef] [PubMed]
- Mariette, N.; Mabon, R.; Corbière, R.; Boulard, F.; Glais, I.; Marquer, B.; Pasco, C.; Montarry, J.; Andrivon, D. Phenotypic and Genotypic Changes in French Populations of Phytophthora infestans: Are Invasive Clones the Most Aggressive? Plant Pathol. 2016, 65, 577–586. [Google Scholar] [CrossRef]
- Mastenbroek, C. Investigations into the Inheritance of the Immunity from Phytophthora infestans de B. of Solanum Demissum Lindl. Euphytica 1952, 1, 187–198. [Google Scholar] [CrossRef]
- Janiszewska, M.; Sobkowiak, S.; Stefańczyk, E.; Śliwka, J. Population Structure of Phytophthora infestans from a Single Location in Poland Over a Long Period of Time in Context of Weather Conditions. Microb. Ecol. 2021, 81, 746–757. [Google Scholar] [CrossRef]
- Wang, X.; El Hadrami, A.; Adam, L.; Daayf, F. US-1 and US-8 Genotypes of Phytophthora infestans Differentially Affect Local, Proximal and Distal Gene Expression of Phenylalanine Ammonia-Lyase and 3-Hydroxy, 3-Methylglutaryl CoA Reductase in Potato Leaves. Physiol. Mol. Plant Pathol. 2004, 65, 157–167. [Google Scholar] [CrossRef]
- Brooks, F.E. Detached-Leaf Bioassay for Evaluating Taro Resistance to Phytophthora Colocasiae. Plant Dis. 2008, 92, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. Murashige. Physiol. Plant. 1962, 15, 474–497. [Google Scholar]
- Caten, C.E.; Jinks, J.L. Spontaneous Variability of Single Isolates of Phytophthora infestans. I. Cultural Variation. Can. J. Bot. 1968, 46, 329–348. [Google Scholar] [CrossRef]
- Fry, W.E.; Patev, S.P.; Myers, K.L.; Bao, K.; Fei, Z. Phytophthora infestans Sporangia Produced in Culture and on Tomato Leaflet Lesions Show Marked Differences in Indirect Germination Rates, Aggressiveness, and Global Transcription Profiles. Mol. Plant-Microbe Interact. 2019, 32, 515–526. [Google Scholar] [CrossRef] [Green Version]
- Dolničar, P.; Rudolf Pilih, K. Genska Banka In Žlahtnjenje Krompirja V Sloveniji. Acta Agric. Slov. 2012, 99, 377–386. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Simko, I.; Piepho, H.P. The Area under the Disease Progress Stairs: Calculation, Advantage, and Application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, RStudio. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 3 May 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytophthora infestans Isolates | ||||
|---|---|---|---|---|
| 02_07 | 09_07 | 90128 | IPO-C | |
| Rioja | 3.28 a | 5.83 a | 6.00 ab | 5.94 ab |
| R7 | 2.56 ab | 6.00 a | 5.67 ab | 7.39 a |
| R15 | 2.13 ab | 5.75 a | 6.50 a | 6.28 ab |
| Sárpo Mira | 0.94b | 1.00 b | 4.56 b | 5.33 b |
| Lusa | 1.72 b | 4.89 a | 5.89 a | 6.82 a |
| L166 | 3.33 a | 5.22 a | 6.44 a | 6.13 ab |
| Sárpo Mira | 0.94 b | 1.00 b | 4.56 b | 5.33 b |
| Colomba | 2.06 a | 6.33 a | 7.44 a | 7.94 a |
| C419 | 1.33 a | 4.33 ab | 5.72 bc | 7.22 a |
| C557 | 0.78 a | 5.39 ab | 6.61 ab | 7.61 a |
| C571 | 2.33 a | 3.39 bc | 5.22 bc | 5.61 b |
| Sárpo Mira | 0.94 a | 1.00 c | 4.56 c | 5.33 b |
| Sylvana | 2.78 a | 7.17 a | 7.00 a | 7.40 a |
| S859 | 1.56 a | 5.67 ab | 5.00 ab | 6.72 ab |
| S989 | 1.83 a | 4.56 b | 5.89 ab | 7.06 ab |
| S999 | 2.27 a | 4.11 b | 6.28 ab | 7.06 ab |
| Sárpo Mira | 0.94 a | 1.00 c | 4.56 b | 5.33 b |
| Gene | Marker Name | Primer Sequence (5′-3′) | Ta (°C) | Amplicon Length (bp) | Reference | |
|---|---|---|---|---|---|---|
| R3a | Sha | F | ATCGTTGTCATGCTATGAGATTGTT | 60 | 982 | [54] |
| R | CTTCAAGGTAGTGGGCAGTATGCTT | |||||
| R3b | R3b | F | GTCGATGAATGCTATGTTTCTCGAGA | 55 | 378 | [51] |
| R | ACCAGTTTCTTGCAATTCCAGATTG | |||||
| Rpi-Smira1 | 45/XI | F | AGAGAGGTTGTTTCCGATAGACC | 58 | 1000 | [36] |
| R | TCGTTGTAGTTGTCATTCCACAC | |||||
| Rpi-Smira2/R8 | 184-81 | F | CCACCGTATGCTCCGCCGTC | 55 | 480 (RsaI) | [46] |
| R | GTTCCACTTAGCCTTGTCTTGCTCA | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blatnik, E.; Horvat, M.; Berne, S.; Humar, M.; Dolničar, P.; Meglič, V. Late Blight Resistance Conferred by Rpi-Smira2/R8 in Potato Genotypes In Vitro Depends on the Genetic Background. Plants 2022, 11, 1319. https://doi.org/10.3390/plants11101319
Blatnik E, Horvat M, Berne S, Humar M, Dolničar P, Meglič V. Late Blight Resistance Conferred by Rpi-Smira2/R8 in Potato Genotypes In Vitro Depends on the Genetic Background. Plants. 2022; 11(10):1319. https://doi.org/10.3390/plants11101319
Chicago/Turabian StyleBlatnik, Eva, Marinka Horvat, Sabina Berne, Miha Humar, Peter Dolničar, and Vladimir Meglič. 2022. "Late Blight Resistance Conferred by Rpi-Smira2/R8 in Potato Genotypes In Vitro Depends on the Genetic Background" Plants 11, no. 10: 1319. https://doi.org/10.3390/plants11101319