
Physiological Characteristics of Cultivated Tepary Bean (Phaseolus acutifolius A. Gray) and Its Wild Relatives Grown at High Temperature and Acid Soil Stress Conditions in the Amazon Region of Colombia

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
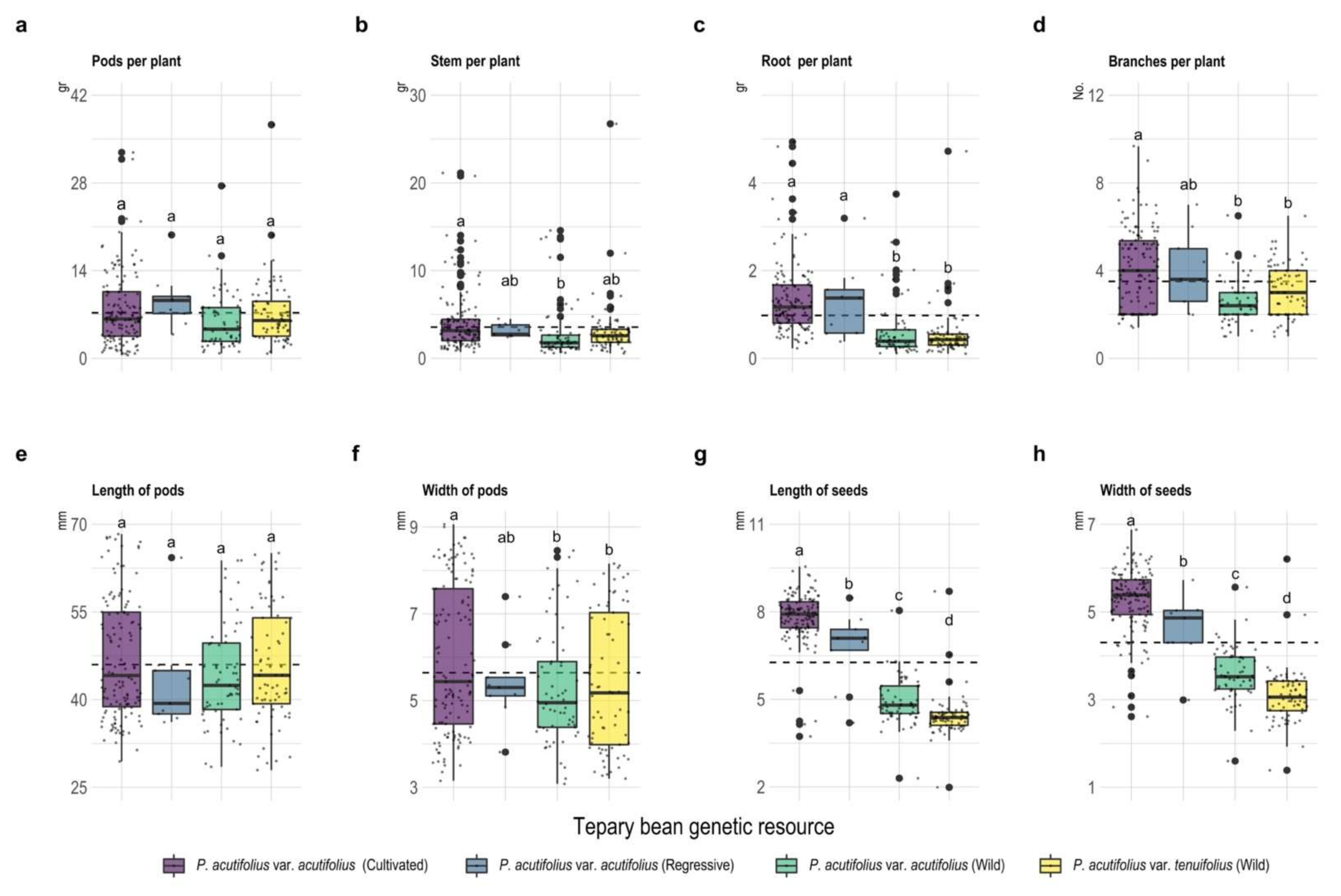
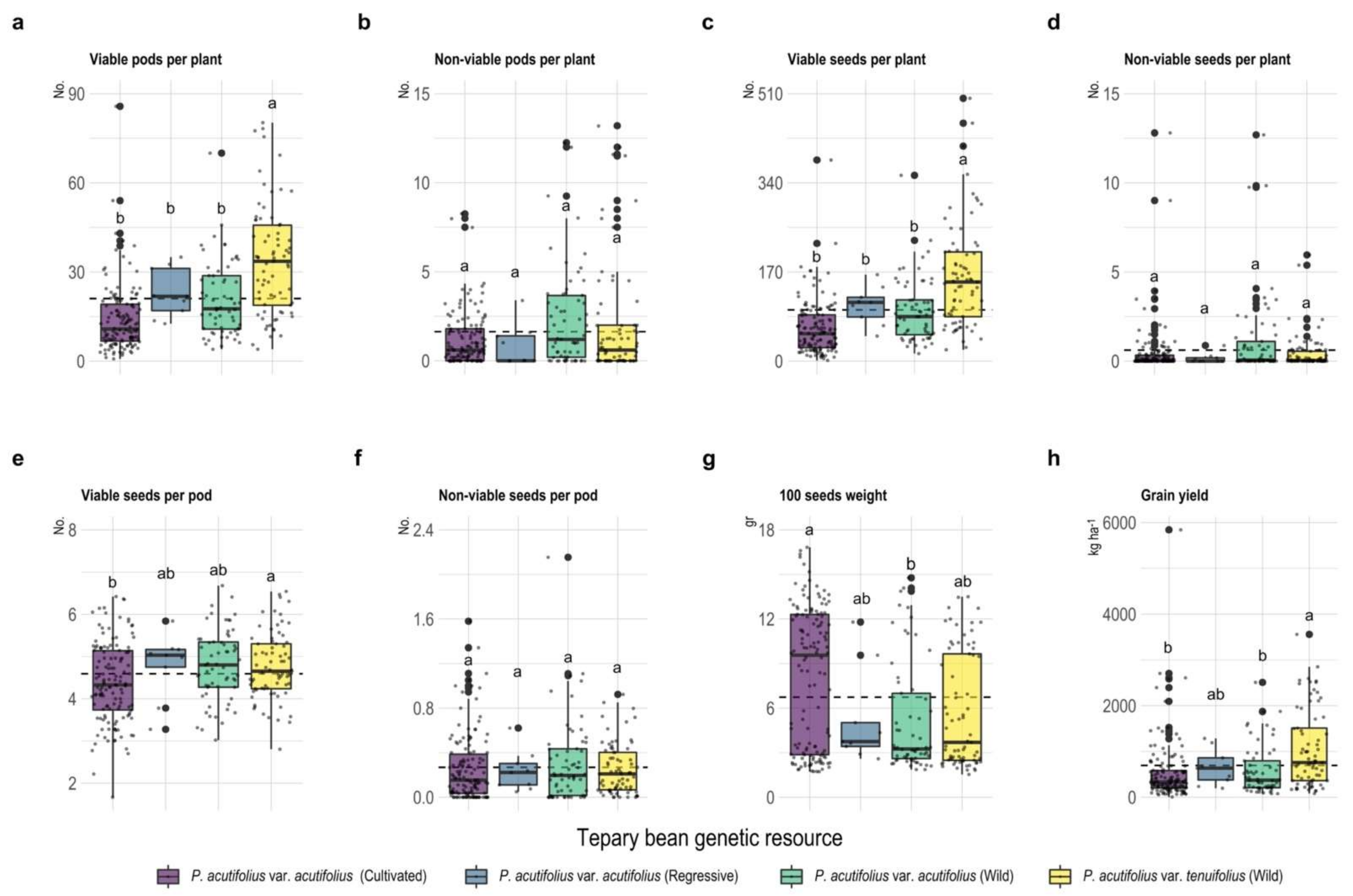
2.1. Development of Different Vegetative Organs under High Temperature and Acid Soil Stress Conditions
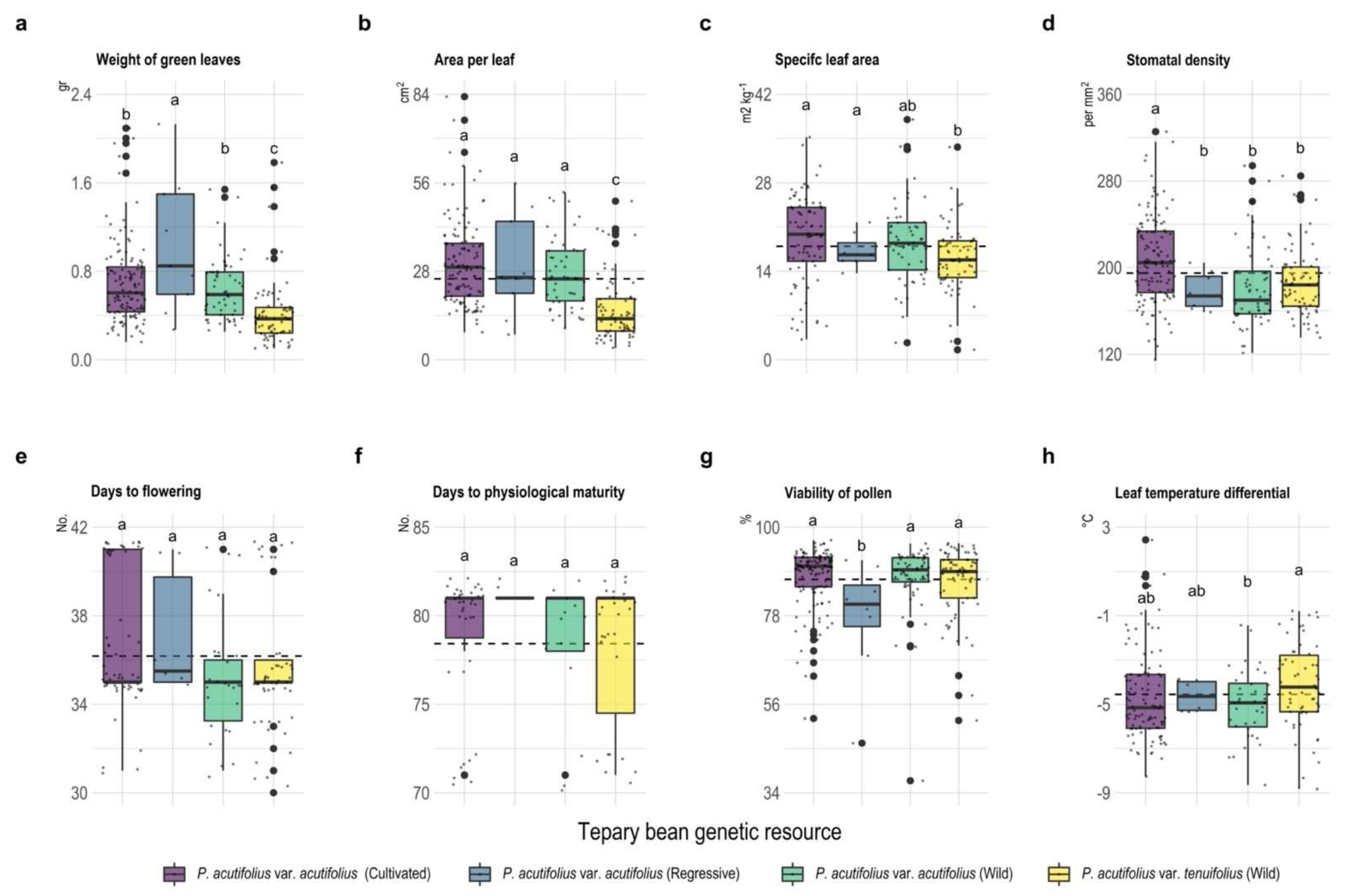
2.2. Morphological and Phenological Traits
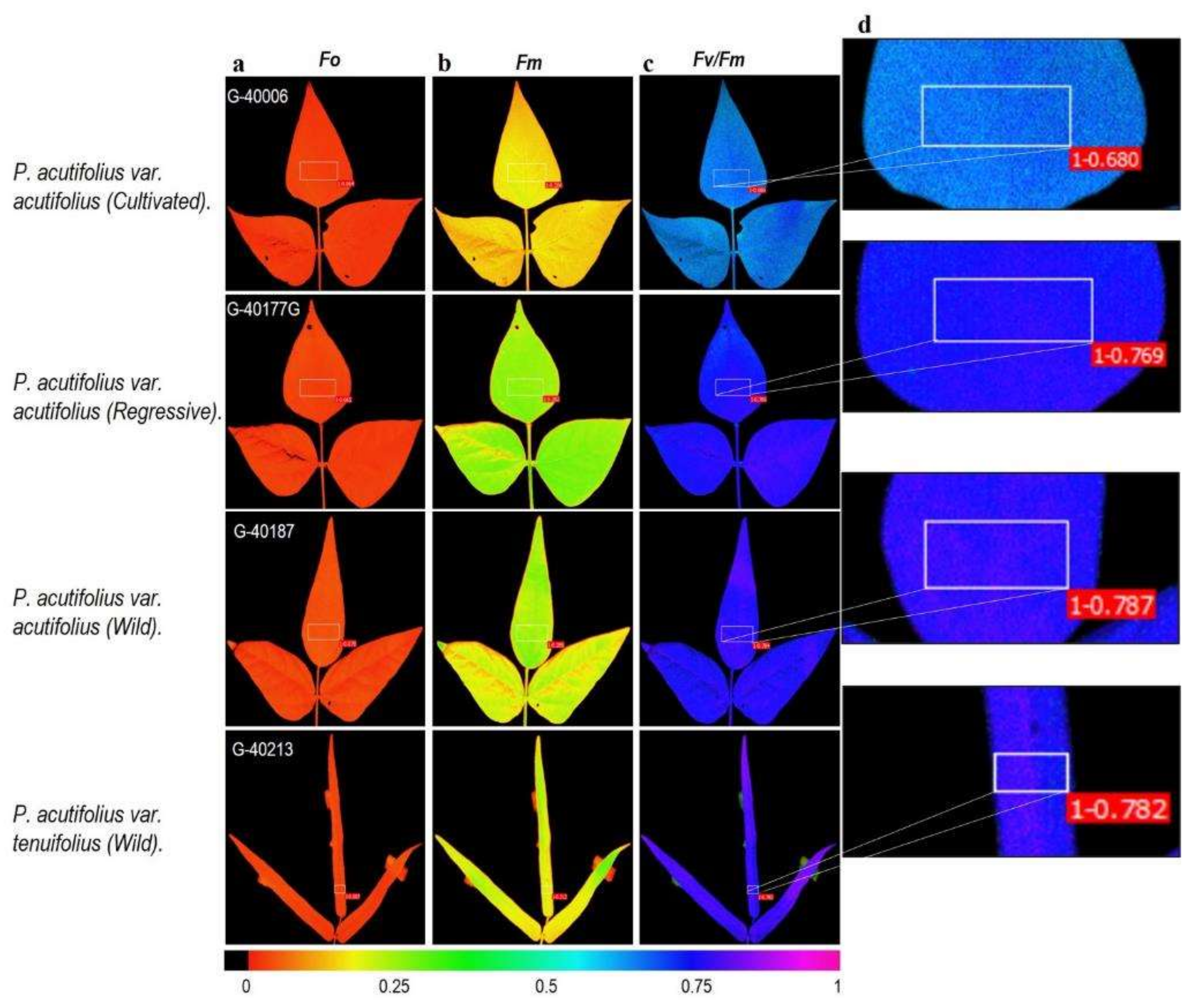
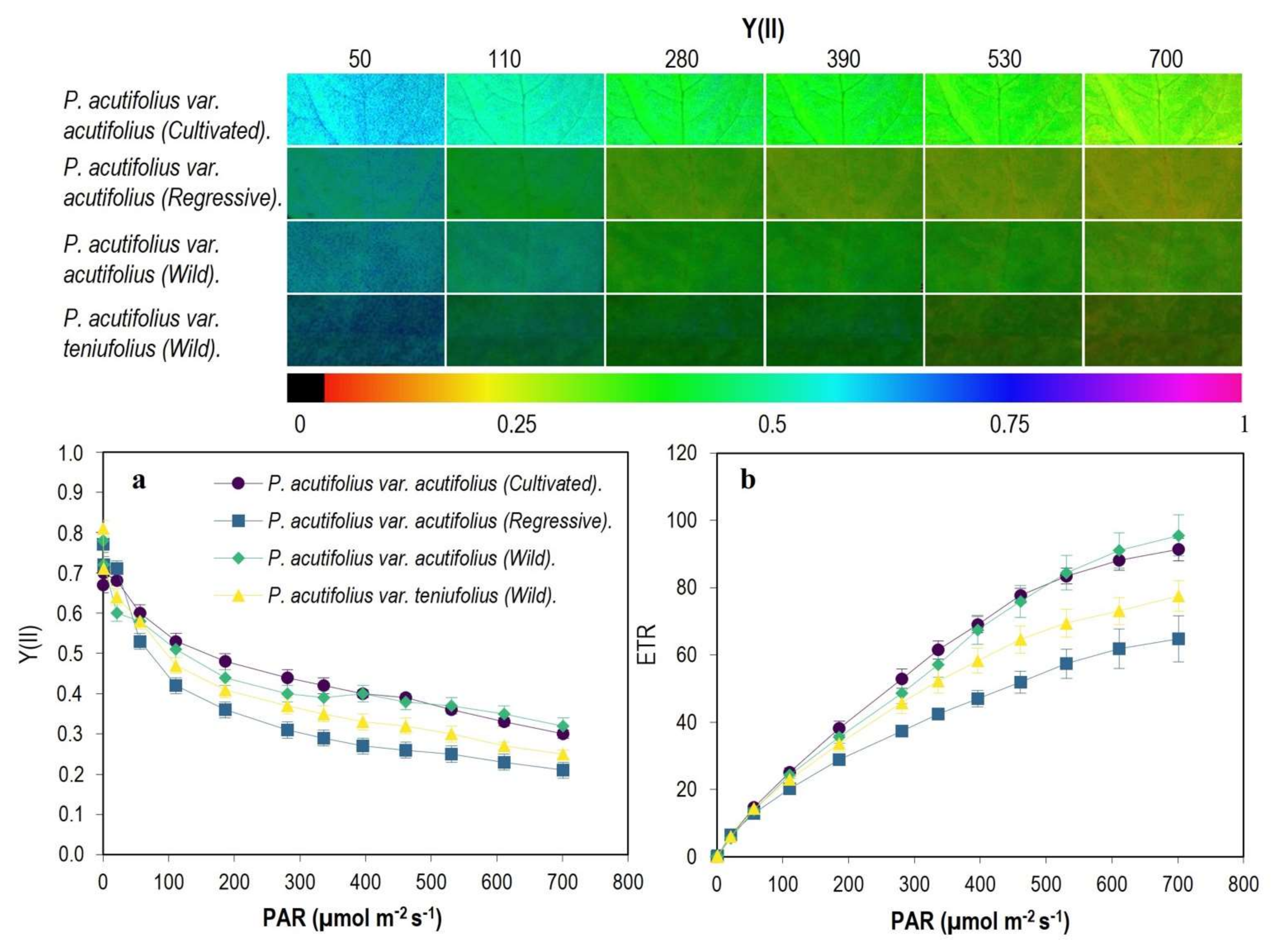
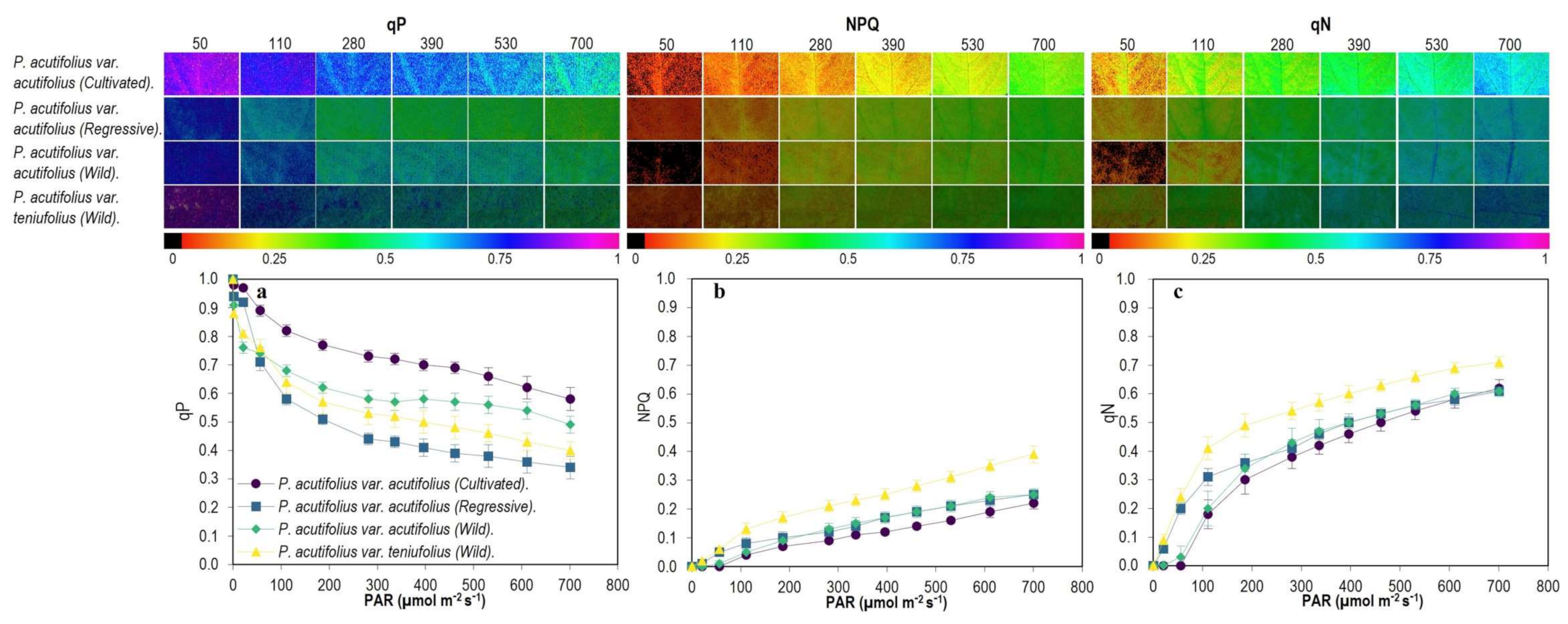
2.3. Fluorescence and Images of Chlorophyll a (Chla) Fluorescence Parameters
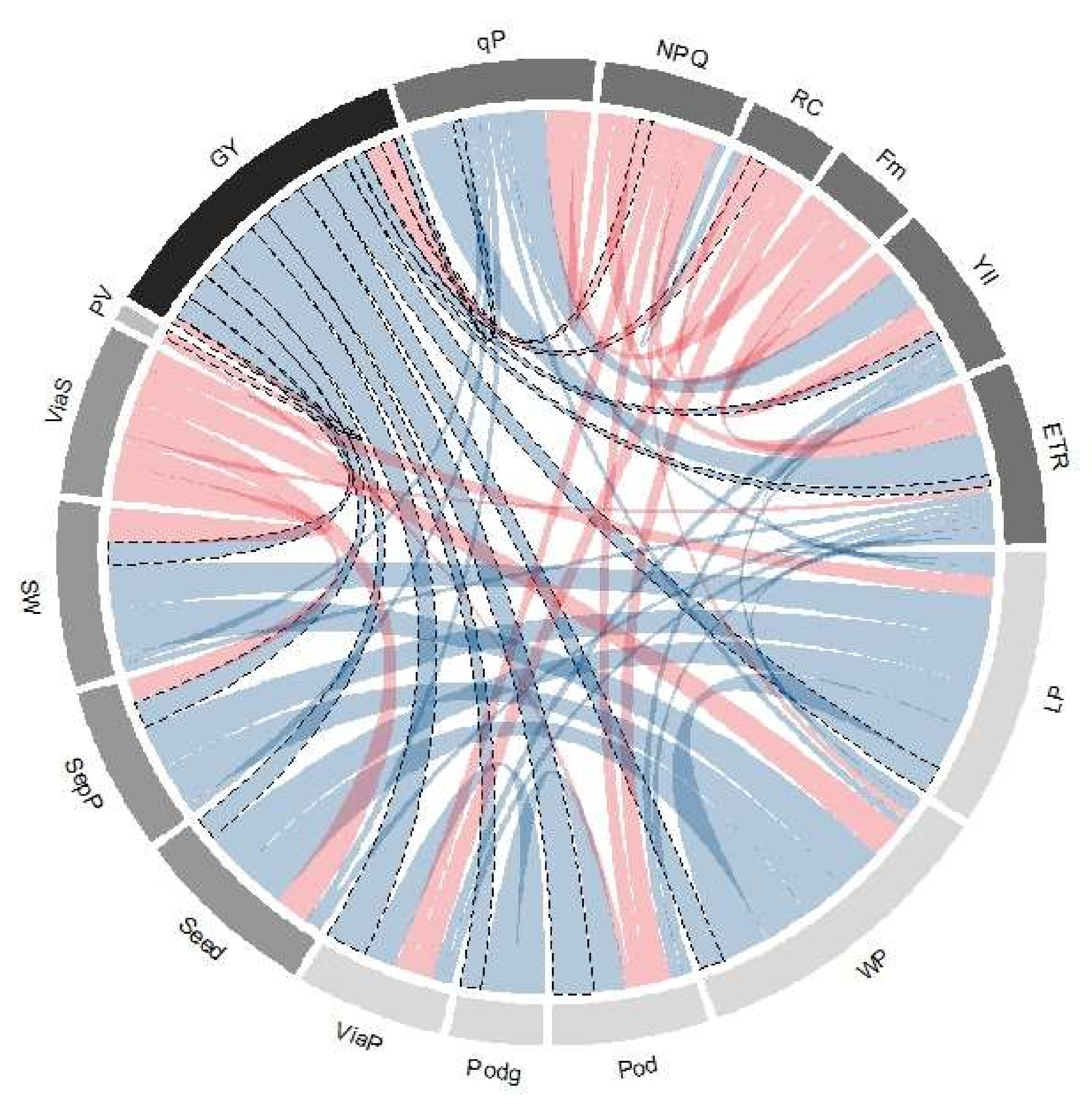
2.4. Correlations between Grain Yield and Agronomic and Physiological Variables under Temperature and Acid Soil Conditions
3. Discussion
3.1. Contribution of Morpho-Agronomic Traits of Tepary Bean for Adaptation to Combined Stress of High Temperature and Acid Soil
3.2. Leaf Characteristics and Phenological Development Contribute to Better Adaptation of Tepary Beans to Combined Stress
3.3. Energy Dissipation Mechanisms in Response to High Temperature and Acid Soil Stress
3.4. Adaptive Mechanisms for Increasing Grain Yield under Combined Stress of High Temperature and Acid Soil
4. Materials and Methods
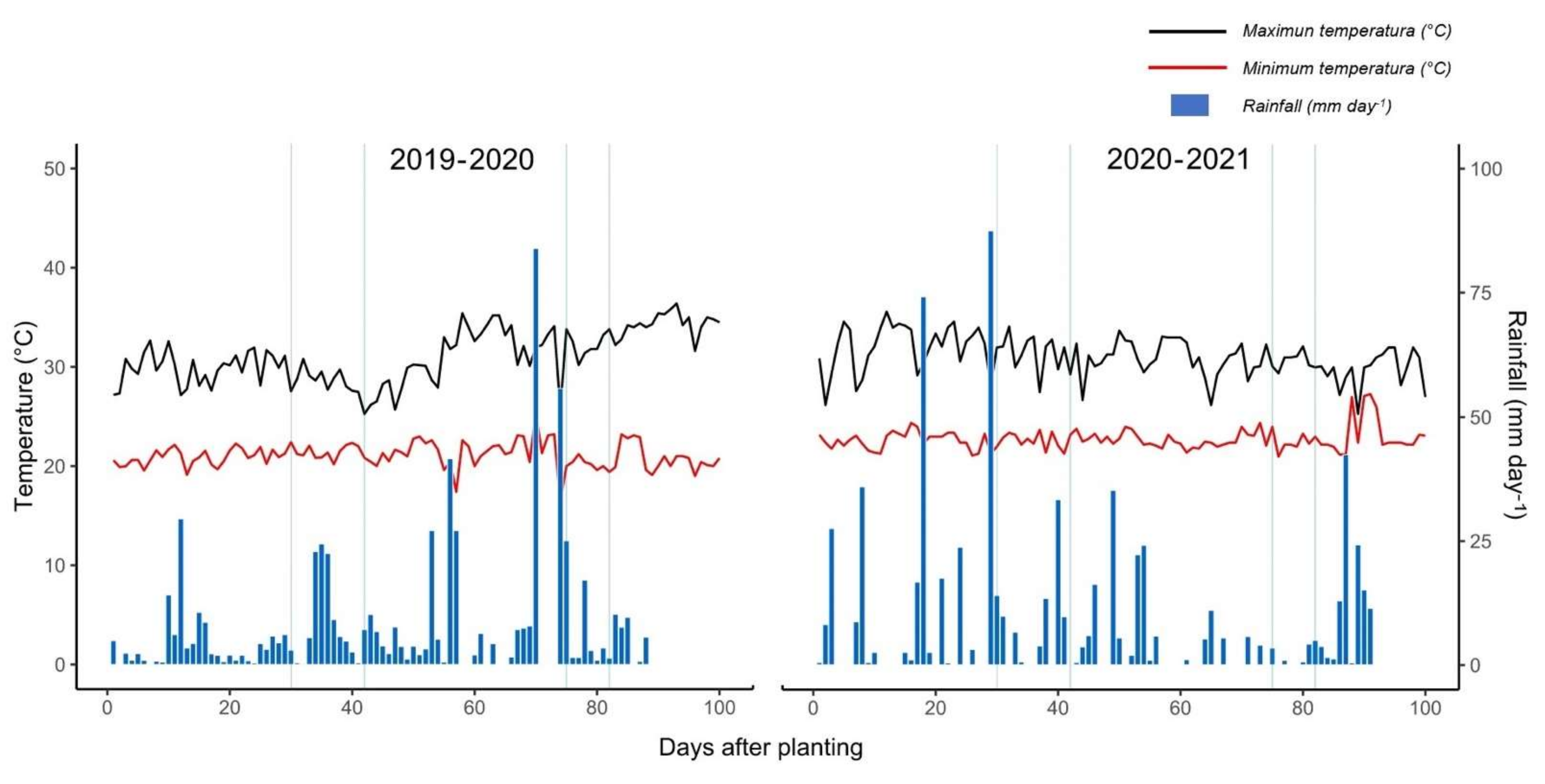
4.1. Experimental Site and Meteorological Conditions
4.2. Plant Material and Experimental Design
4.3. Development of Different Vegetative Organs under High Temperature and Acid Soil Stress Conditions
4.4. Morphological and Phenological Characteristics
4.5. Fluorescence and Imaging for Chlorophyll (Chla) Parameters
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chmura, H.E.; Glass, T.W.; Williams, C.T. Biologging physiological and ecological responses to climatic variation: New tools for the climate change era. Front. Ecol. Evol. 2018, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Masson-Delmotte, V.; Pörtner, H.; Skea, J.; Zhai, P.; Roberts, D.; Shukla, P.; Pirani, A.; Pidcock, R.; Chen, Y.; Moufouma-Okia, W.; et al. Calentamiento global de 1.5 °C Informe especial del IPCC sobre los impactos del calentamiento global de 1.5 °C con respecto a los niveles preindustriales y las trayectorias correspondientes que deberían seguir las e. In Resumen Para Responsables de Políticas; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- Pourkheirandish, M.; Golicz, A.A.; Bhalla, P.L.; Singh, M.B. Global role of crop genomics in the face of climate change. Front. Plant Sci. 2020, 11, 922. [Google Scholar] [CrossRef] [PubMed]
- Raggi, L.; Caproni, L.; Carboni, A.; Negri, V. Genome-wide association study reveals candidate genes for flowering time variation in common bean (Phaseolus vulgaris L.). Front. Plant Sci. 2019, 10, 962. [Google Scholar] [CrossRef] [Green Version]
- Porch, T.G.; Hall, A.E. Heat tolerance. In Genomics and Breeding for Climate-Resilient Crops; Springer: Berlin/Heidelberg, Germany, 2013; Volume 2, pp. 167–202. [Google Scholar] [CrossRef]
- Buitrago-Bitar, M.A.; Cortés, A.J.; López-Hernández, F.; Londoño-Caicedo, J.M.; Muñoz-Florez, J.E.; Muñoz, L.C.; Blair, M.W. Allelic diversity at abiotic stress responsive genes in relationship to ecological drought indices for cultivated tepary bean, Phaseolus acutifolius A. Gray, and its wild relatives. Genes 2021, 12, 556. [Google Scholar] [CrossRef] [PubMed]
- Leal-Delgado, R.; Peña-Valdivia, C.B.; García-Nava, R.; García-Esteva, A.; Martínez-Barajas, E.; Padilla-Chacón, D. Phenotypical, physiological and biochemical traits of the vegetative growth of wild tepary bean (Phaseolus acutifolius) under restricted water conditions. S. Afr. J. Plant Soil 2019, 36, 261–270. [Google Scholar] [CrossRef]
- Beebe, S.; Ramírez, J.; Jarvis, A.; Rao, I.; Mosquera, J.; Bueno, J.; Blair, M. Genetic improvement of common beans and the challenges of climate change. In Crop Adaptation to Climate Change; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011; pp. 356–369. [Google Scholar] [CrossRef]
- Polania, J.A.; Poschenrieder, C.; Beebe, S.; Rao, I.M. Effective use of water and increased dry matter partitioned to grain contribute to yield of common bean improved for drought resistance. Front. Plant Sci. 2016, 7, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, J.C.; Polanía, J.A.; Contreras, A.; Suárez, L.; Beebe, S.; Rao, I. Agronomical, phenological and physiological performance of common bean lines in the Amazon region of Colombia. Theor. Exp. Plant Physiol. 2018, 30, 303–320. [Google Scholar]
- Traub, J.; Porch, T.; Naeem, M.; Urrea, C.; Austic, G.; Kelly, J.; Loescher, W. Screening for heat tolerance in Phaseolus spp. using multiple methods. Crop Sci. 2018, 58, 2459–2469. [Google Scholar] [CrossRef]
- Moghaddam, S.M.; Oladzad, A.; Koh, C.; Ramsay, L.; Hart, J.; Mamidi, S.; Hoopes, G.; Sreedasyam, A.; Wiersma, A.; Zhao, D.; et al. The tepary bean genome provides insight into evolution and domestication under heat stress. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Rivera, M.; Muñoz, J.E.; Sarsu, F.; Rao, I.M. Heat stress-induced changes in shoot and root characteristics of genotypes of tepary bean (Phaseolus acutifolius A. Gray), common bean (Phaseolus vulgaris L.) and their interspecific lines. Aust. J. Crop Sci. 2021, 15, 51–59. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Debouck, D.; Rivera, M.; Muñoz, J.; Alpala, D.; Sarsu, F.; Rao, I. Mutation breeding for heat and drought tolerance in tepary bean (Phaseolus acutifolius A. Gray). Aust. J. Crop Sci. 2021, 15, 60–68. [Google Scholar] [CrossRef]
- Parker, T.A.; Gepts, P. Population Genomics of Phaseolus spp.: A Domestication Hotspot; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–83. [Google Scholar] [CrossRef]
- Miklas, P.N.; Schwartz, H.F.; Salgado, M.O.; Nina, R.; Beaver, J.S. Reaction of select tepary bean to ashy stem blight and fusarium wilt. HortScience 1998, 33, 136–139. [Google Scholar]
- Porch, T.G.; Beaver, J.; Debouck, D.; Jackson, S.; Kelly, J.; Dempewolf, H. Use of Wild Relatives and Closely Related Species to Adapt Common Bean to Climate Change. Agronomy 2013, 3, 433–461. [Google Scholar] [CrossRef] [Green Version]
- Bayuelo-Jiménez, J.S.; Debouck, D.G.; Lynch, J.P. Salinity tolerance in Phaseolus species during early vegetative growth. Crop Sci. 2002, 42, 2184–2192. [Google Scholar] [CrossRef]
- Suárez, J.C.; Polania, J.; Contreras, A.; Rodriguez, L.; Machado, L.; Ordoñez, C.; Rao, I. Adaptation of common bean lines to high temperature conditions: Genotypic differences in phenological and agronomic performance. Euphytica 2020, 216, 1–20. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Noga, G.J. Comparative drought postponing and tolerance ootentials of two Tepary bean lines in relation to seed yield. African Crop Sci. J. 2005, 13, 49–60. [Google Scholar]
- Beebe, S.; Rao, I.; Blair, M.; Acosta, J. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Amarteifio, J.O.; Moholo, D. The chemical composition of four legumes consumed in Botswana. J. Food Compos. Anal. 1998, 11, 329–332. [Google Scholar] [CrossRef]
- Bhardwaj, H.L.; Hamama, A.A. Oil and fatty acid composition of tepary bean seed. HortScience 2005, 40, 1436–1438. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, H.L.; Hamama, A.A. Protein and mineral composition of tepary bean seed. HortScience 2004, 39, 1363–1365. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Duque, M.C.; Debouck, D.G.; Blair, M.W. Taxonomy of tepary bean and wild relatives as determined by amplified fragment length polymorphism (AFLP) markers. Crop Sci. 2006, 46, 1744–1754. [Google Scholar] [CrossRef]
- Blair, M.W.; Pantoja, W.; Carmenza Muñoz, L. First use of microsatellite markers in a large collection of cultivated and wild accessions of tepary bean (Phaseolus acutifolius A. Gray). Theor. Appl. Genet. 2012, 125, 1137–1147. [Google Scholar] [CrossRef]
- CIAT. Unidad de Recursos Fitogenéticos, Frijol. 2021. Available online: https://genebank.ciat.cgiar.org/genebank/beancollection.do (accessed on 25 May 2021).
- Debouck, D.G.; Santaella, M.; Santos, L.G. History and impact of a bean (Phaseolus spp., Leguminosae, Phaseoleae) collection. Genet. Resour. 2021, 2, 21–43. [Google Scholar] [CrossRef]
- Chacón, S.M.I.; Pickersgill, B.; Debouck, D.G. Domestication patterns in common bean (Phaseolus vulgaris L.) and the origin of the Mesoamerican and Andean cultivated races. Theor. Appl. Genet. 2005, 110, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Freytag, G.F.; Debouck, D.G. Taxonomy, Distribution, and Ecology of the Genus Phaseolus (Leguminosae–Papilionoideae) in North America, Mexico, and Central America. Botanical Research Institute of Texas (BRIT). Econ. Bot. 2003, 57, 421. [Google Scholar] [CrossRef]
- Debouck, D.G. Systematics and morphology. In Common Beans: Research for Crop Improvement; van Schoonhoven, A., Voysest, O., Eds.; CIAT: Rome, Italy, 1991; pp. 55–118. [Google Scholar]
- Rahman, R.; Upadhyaya, H. Aluminium toxicity and its tolerance in plants: A Review. J. Plant Biol. 2020, 64, 101–121. [Google Scholar] [CrossRef]
- Mendoza-Soto, A.B.; Naya, L.; Leija, A.; Hernández, G. Responses of symbiotic nitrogen-fixing common bean to aluminum toxicity and delineation of nodule responsive microRNAs. Front. Plant Sci. 2015, 6, 587. [Google Scholar] [CrossRef] [PubMed]
- Suárez, J.C.; Urban, M.; Contreras, A.; Noriega, J.; Deva, C.; Beebe, S.; Polania, J.; Casanoves, F.; Rao, I. Water use, leaf cooling and carbon assimilation efficiency of heat resistant common beans evaluated in Western Amazonia. Front. Plant Sci. 2021, 12, 644010. [Google Scholar] [CrossRef]
- Bhattarai, G.P.; Meyerson, L.A.; Anderson, J.; Cummings, D.; Allen, W.J.; Cronin, J.T. Biogeography of a plant invasion: Genetic variation and plasticity in latitudinal clines for traits related to herbivory. Ecol. Monogr. 2017, 87, 57–75. [Google Scholar] [CrossRef]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Brix, H. Cosmopolitan species as models for ecophysiological responses to global change: The common reed phragmites australis. Front. Plant Sci. 2017, 8, 1833. [Google Scholar] [CrossRef] [Green Version]
- Salgado, M.O.; Schwartz, H.F.; Brick, M.A.; Corrales, M.A.P. Resistance to Fusarium oxysporum f. sp. phaseoli in tepary beans (Phaseolus acutifolius). Plant Dis. 1994, 78, 357–360. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Keutgen, N.; Tawfika, A.A.; Noga, G. Dehydration-avoidance responses of tepary bean lines differing in drought resistance. J. Plant Physiol. 2002, 159, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S. Common bean breeding in the tropics. Plant Breed. Rev. 2012, 36, 357–426. [Google Scholar]
- Crossa, J.; Gauch, H.G.; Zobel, R.W. Additive main effects and multiplicative interaction analysis of two international maize cultivar trials. Crop Sci. 1990, 30, 493–500. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Amelework, B.; Shimelis, H.; Modi, A.T.; Mashilo, J. Genetic differentiation among selected tepary bean collections revealed by morphological traits and simple sequence repeat markers. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 608–618. [Google Scholar] [CrossRef]
- Orduño, A.; Troyo, E. Morfología y Desarrollo de Frijol Tepari Phaseolus acutifolius A. Gray; Centro de Investigaciones Biologicas del Noreste: La Paz, Mexico, 2003. [Google Scholar]
- Baath, G.S.; Rocateli, A.C.; Kakani, V.G.; Singh, H.; Northup, B.K.; Gowda, P.H.; Katta, J.R. Growth and physiological responses of three warm-season legumes to water stress. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Arunakumara, K.K.I.U.; Walpola, B.C.; Yoon, M.H. Aluminum toxicity and tolerance mechanism in cereals and legumes—A review. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 1–9. [Google Scholar] [CrossRef]
- Florez, C.; Debouck, D.; Gutierrez, A. Patrones de diversidad genética y domesticación en frijol Tepari (Phaseolus acutifolius A Gray). Acta Agron. 1997, 47, 19–24. [Google Scholar]
- Gepts, P. Genetic diversity of seed storage proteins in plants. In Plant Population Genetics, Breeding, and Genetic Resources; Brown, A.H.D., Clegg, M.T., Kahler, A.L., Weir, B.S., Eds.; Sinauer Associates Inc.: Sunderland, MA, USA, 1990; pp. 64–82. [Google Scholar]
- Lersten, N.R.; Carlson, J.B. Vegetative morphology. In Soybeans: Improvement, Production, and Uses; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 15–57. [Google Scholar] [CrossRef]
- Rosas, J.C.; Erazo, J.D.; Moncada, J.R. Tolerancia a la Sequía en Germoplasma de Frijol Común y Frijol Tepari. CEIBA 1991, 32, 91–106. [Google Scholar]
- Assefa, T.; Wu, J.; Beebe, S.; Rao, I.; Marcomin, D.; Claude, R. Participatory plant breeding with traders and farmers for white pea bean in Ethiopia. J. Agric. Educ. Ext. 2014, 20, 497–512. [Google Scholar] [CrossRef]
- Rao, I.; Beebe, S.; Polania, J.; Ricaute, J.; Cajiao, C.; Garcia, R.; River, M. Can tepary bean be a model for improvement of drought resistance in common bean? African Crop Sci. J. 2013, 21, 265–281. [Google Scholar]
- Schinkel, C.; Gepts, P. Phaseolin diversity in the tepary bean, Phaseolus acutifolius A. Gray. Plant Breed. 1988, 101, 292–301. [Google Scholar] [CrossRef]
- Schinkel, C.; Gepts, P. Allozyme variability in the tepary bean, Phaseolus acutifolius A. Gray. Plant Breed. 1989, 102, 182–195. [Google Scholar] [CrossRef]
- Garvin, D.F.; Weeden, N.F. Isozyme evidence supporting a single geographic origin for domesticated tepary bean. Crop Sci. 1994, 34, 1390–1395. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Shimelis, H.A.; Amelework, B.; Modi, A.T.; Mashilo, J. Variance components and heritability of yield and yield-related traits in tepary bean (Phaseolus acutifolius). South African J. Plant Soil 2019, 36, 117–128. [Google Scholar] [CrossRef]
- Rosales, M.A.; Ocampo, E.; Rodríguez-Valentín, R.; Olvera-Carrillo, Y.; Acosta-Gallegos, J.; Covarrubias, A.A. Physiological analysis of common bean (Phaseolus vulgaris L.) cultivars uncovers characteristics related to terminal drought resistance. Plant Physiol. Biochem. 2012, 56, 24–34. [Google Scholar] [CrossRef]
- Blair, M.W.; González, L.F.; Kimani, P.M.; Butare, L. Genetic diversity, inter-gene pool introgression and nutritional quality of common beans (Phaseolus vulgaris L.) from Central Africa. Theor. Appl. Genet. 2010, 121, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Pratt, R.C.; Nabhan, G.P. Evolution and Diversity of Phaseolus Acutifolius Genetic Resources; Springer: Berlin/Heidelberg, Germany, 1988; pp. 409–440. [Google Scholar] [CrossRef]
- Ticha, I. Photosynthetic characteristics during ontogenesis of leaves. 7. Stomata density and sizes. Photosynthetica 1982, 16, 375–471. [Google Scholar]
- Garcia, M.; Madriz, P. Anatomía foliar comparada de cinco genotipos de frijol mungo, Vigna radiata (L.) Wilczek. Rev. Fac. Agron. Univ. Zulia 1999, 16, 610–620. [Google Scholar]
- Kim, L.; Balani, S.; Edelberg, M.; Macke, N. Effects of various environmental factors on stomatal density, area, and potential conductance index. J. Emerg. Investig. 2021, 4, 1–7. Available online: https://emerginginvestigators.org/articles/effects-of-various-environmental-factors-on-stomatal-density-area-and-potential-conductance-index/pdf (accessed on 17 June 2021).
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An ‘electronic fluorescent pictograph’ browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Biswal, A.K.; Dionora, J.; Perdigon, K.M.; Balahadia, C.P.; Mazumdar, S.; Chater, C.; Lin, H.; Coe, R.; Kretzschmar, T.; et al. CRISPR-Cas9 and CRISPR-Cpf1 mediated targeting of a stomatal developmental gene EPFL9 in rice. Plant Cell Rep. 2017, 36, 745–757. [Google Scholar] [CrossRef]
- Wang, C.; Liu, S.; Dong, Y.; Zhao, Y.; Geng, A.; Xia, X.; Yin, W. PdEPF1 regulates water-use efficiency and drought tolerance by modulating stomatal density in poplar. Plant Biotechnol. J. 2016, 14, 849–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anugoolprasert, O.; Kinoshita, S.; Naito, H.; Shimizu, M.; Ehara, H. Effect of low pH on the growth, physiological characteristics and nutrient absorption of sago palm in a hydroponic system. Plant Prod. Sci. 2012, 15, 125–131. [Google Scholar] [CrossRef]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of stomatal density and morphology on water-use efficiency in a changing world. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.; Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.; Gray, J. Reducing Stomatal Density in Barley Improves Drought Tolerance without Impacting on Yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef] [Green Version]
- Araus, J.L.; Slafer, G.A.; Reynolds, M.P.; Royo, C. Plant breeding and drought in C3 cereals: What should we breed for? Ann. Bot. 2002, 85, 925–940. [Google Scholar] [CrossRef]
- Rana, J.C.; Sharma, T.R.; Tyagi, R.K.; Chahota, R.K.; Gautam, N.K.; Singh, M.; Sharma, P.N.; Ojha, S.N. Characterisation of 4274 accessions of common bean (Phaseolus vulgaris L.) germplasm conserved in the Indian gene bank for phenological, morphological and agricultural traits. Euphytica 2015, 205, 441–457. [Google Scholar] [CrossRef]
- Suzuki, K.; Tsukaguchi, T.; Takeda, H.; Egawa, Y. Decrease of pollen stainability of green bean at high temperatures and relationship to heat tolerance. J. Am. Soc. Hortic. Sci. 2001, 126, 571–574. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.E. Comparative ecophysiology of cowpea, common bean, and peanut. In Physiology and Biotechnology Integration for Plant Breeding; CRC Press: Boca Raton, FL, USA, 2004; pp. 243–287. [Google Scholar] [CrossRef]
- Castonguay, Y.; Markhart, A.H. Leaf gas exchange in water—Stressed common bean and tepary bean. Crop Sci. 1992, 32, 980–986. [Google Scholar] [CrossRef]
- Yao, X.; Zhou, H.; Zhu, Q.; Li, C.; Zhang, H.; Wu, J.; Xie, F. Photosynthetic response of Soybean leaf to wide light-fluctuation in maize-soybean intercropping system. Front. Plant Sci. 2017, 8, 1695. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping plant responses to biotic stress by chlorophyll fluorescence Imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.P.; da Matta, F.M.; da Matta, J.C. Responses of the photosynthetic apparatus to aluminum stress in two sorghum cultivars. J. Plant Nutr. 2002, 25, 821–832. [Google Scholar] [CrossRef]
- Jiang, H.X.; Chen, L.; Zheng, J.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on Photosystem II photochemistry in citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef]
- Wujeska-Klause, A.; Bossinger, G.; Tausz, M. Responses to heatwaves of gas exchange, chlorophyll fluorescence and antioxidants ascorbic acid and glutathione in congeneric pairs of Acacia and Eucalyptus species from relatively cooler and warmer climates. Trees 2015, 29, 1929–1941. [Google Scholar] [CrossRef]
- Sterling, A.; Rodríguez, N.; Quiceno, E.; Trujillo, F.; Clavijo, A.; Suárez-Salazar, J.C. Dynamics of photosynthetic responses in 10 rubber tree (Hevea brasiliensis) clones in Colombian Amazon: Implications for breeding strategies. PLoS ONE 2019, 14, e0226254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.H.; Wang, E.M.; Zhao, T.F.; Wang, Q.Q.; Chen, L.J. Characteristics of chlorophyll fluorescence and antioxidant-oxidant balance in PEPC and PPDK transgenic rice under aluminum stress. Russ. J. Plant Physiol. 2018, 65, 49–56. [Google Scholar] [CrossRef]
- Kitao, M.; Lei, T.T. Circumvention of over-excitation of PSII by maintaining electron transport rate in leaves of four cotton genotypes developed under long-term drought. Plant Biol. 2007, 9, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tan, M.; Xu, Y.; Zhao, Y.; Cheng, F.; Ye, S.; Jiang, W. Effect of low pH and aluminum toxicity on the photosynthetic characteristics of different fast-growing Eucalyptus vegetatively propagated clones. PLoS ONE 2015, 10, e0130963. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, A.; Ostendorf, E.; Kohzuma, K.; Hoh, D.; Strand, D.D.; Sato-Cruz, M.; Savage, L.; Cruz, J.; Fisher, N.; Froehlich, J.; et al. Chloroplast ATP synthase modulation of the thylakoid proton motive force: Implications for photosystem I and photosystem II photoprotection. Front. Plant Sci. 2017, 8, 719. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, M.; Mekala, N.R.; Aro, E.M. Photosystem II photoinhibition-repair cycle protects Photosystem I from irreversible damage. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 210–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, Ü. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef] [PubMed]
- Hasni, I.; Yaakoubi, H.; Hamdani, S.; Tajmir-Riahi, H.A.; Carpentier, R. Mechanism of interaction of Al3+ with the proteins composition of Photosystem II. PLoS ONE 2015, 10, e0120876. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Ahammed, G.J.; Li, X.; Shi, K. Elevated CO2 Photosynthesis under high temperature by attenuating the functional limitations to energy fluxes, electron transport and redox homeostasis in tomato leaves. Front. Plant Sci. 2018, 871, 1739. [Google Scholar] [CrossRef] [Green Version]
- Micheletto, S.; Rodriguez-Uribe, L.; Hernandez, R.; Richins, R.D.; Curry, J.; O’Connell, M.A. Comparative transcript profiling in roots of Phaseolus acutifolius and P. vulgaris under water deficit stress. Plant Sci. 2007, 173, 510–520. [Google Scholar] [CrossRef]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A tool for large-scale plant phenotyping connected to the open photosynQ network. R. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Sonoike, K. Screening of mutants using chlorophyll fluorescence. J. Plant Res. 2021, 134, 653–664. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.D.; Niyogi, K.K. Use of a pulse-amplitude modulated chlorophyll fluorometer to study the efficiency of photosynthesis in Arabidopsis plants. Methods Mol. Biol. 2011, 775, 299–310. [Google Scholar] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Rios, V.S.; Rios, J.A.; Aucique-Pérez, C.E.; Silveira, P.R.; Barros, A.V.; Rodrigues, F.Á. Leaf gas exchange and chlorophyll a fluorescence in soybean leaves infected by Phakopsora pachyrhizi. J. Phytopathol. 2018, 166, 75–85. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Visualization of a correlation matrix. R Package Version 0.73 2017, 230, 11–15. [Google Scholar]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: The R project for statistical computing. In R Foundation for Statistical Computing; R Development Core Team: Vienna, Austria, 2021; ISBN 3-900051-07-0. [Google Scholar]
- Di Rienzo, J.A.; Balzarini, M.; Gonzalez, L.; Casanoves, F.; Tablada, M.; Robledo, C.W. Infostat—Software Estadístico; Versión 2019; Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2019. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | P. Acutifolius var. Acutifolius (Cultivated) | P. Acutifolius var. Acutifolius (Regressive) | P. Acutifolius var. Acutifolius (Wild) | P. Acutifolius var. tenuifolius (Wild) |
|---|---|---|---|---|
| Plant height at 10 days | 0.20 * | 0.17 | 0.12 | 0.12 |
| Number of branches/plant | −0.15 | −0.31 | 0.10 | 0.45 *** |
| Stem weight/plant (g) | 0.31 *** | 0.06 | −0.03 | 0.07 |
| Fresh Weight (g) | −0.04 | −0.01 ** | 0.32 | −0.04 |
| Total number of pods/plant | 0.84 *** | 0.63 | 0.65 *** | 0.62 *** |
| Pod weight/plant (g) | 0.41 *** | 0.73 ** | 0.47 *** | 0.34 ** |
| Number of viable Pods/plant | 0.84 *** | 0.61 | 0.68 *** | 0.63 *** |
| Number of non-Viable pods per plant | 0.25 ** | 0.28 | 0.18 | 0.03 |
| Width of pods (mm) | 0.36 *** | 0.49 | 0.62 *** | 0.67 *** |
| Length of pods (mm) | 0.41 *** | 0.53 | 0.59 *** | 0.62 *** |
| Seed weight/plant (g) | 0.35 *** | 0.88 *** | 0.67 *** | 0.73 *** |
| Number of viable seeds (No) | −0.04 | −0.27 | −0.26 * | −0.37 ** |
| Number of seeds per pod (g) | 0.42 *** | 0.78 ** | 0.70 *** | 0.76 *** |
| Number of viable seeds per plant | 0.77 *** | 0.37 | 0.59 *** | 0.47 *** |
| Number of non-viable seeds per plant | 0.18 * | 0.10 | 0.11 | 0.06 |
| 100 seed weight (SW, g) | 0.35 *** | 0.88 *** | 0.67 *** | 0.73 *** |
| Relative chlorophyll (RC) | −0.24 ** | 0.03 | −0.03 | −0.27 * |
| Pollen Viability (PV, %) | 0.14 | −0.24 | −0.14 | −0.33 ** |
| Fm′ | −0.11 | 0.07 | 0.30 ** | 0.14 |
| Y(II) | 0.11 | 0.22 | 0.24 | 0.28 ** |
| NPQ | −0.18 * | −0.09 | −0.32 ** | −0.30 ** |
| qN | −0.10 | 0.03 | −0.35 ** | −0.24 ** |
| ETR | 0.12 *** | 0.50 | 0.15 | 0.26 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez, J.C.; Contreras, A.T.; Anzola, J.A.; Vanegas, J.I.; Rao, I.M. Physiological Characteristics of Cultivated Tepary Bean (Phaseolus acutifolius A. Gray) and Its Wild Relatives Grown at High Temperature and Acid Soil Stress Conditions in the Amazon Region of Colombia. Plants 2022, 11, 116. https://doi.org/10.3390/plants11010116
Suárez JC, Contreras AT, Anzola JA, Vanegas JI, Rao IM. Physiological Characteristics of Cultivated Tepary Bean (Phaseolus acutifolius A. Gray) and Its Wild Relatives Grown at High Temperature and Acid Soil Stress Conditions in the Amazon Region of Colombia. Plants. 2022; 11(1):116. https://doi.org/10.3390/plants11010116
Chicago/Turabian StyleSuárez, Juan Carlos, Amara Tatiana Contreras, José Alexander Anzola, José Iván Vanegas, and Idupulapati M. Rao. 2022. "Physiological Characteristics of Cultivated Tepary Bean (Phaseolus acutifolius A. Gray) and Its Wild Relatives Grown at High Temperature and Acid Soil Stress Conditions in the Amazon Region of Colombia" Plants 11, no. 1: 116. https://doi.org/10.3390/plants11010116