Author Contributions
Conceptualization, V.D.Z. and I.B.S.; methodology, E.Y.-T., J.F.S., W.W., C.L.C., L.H.K.-V., A.S. and T.A.; software, T.A.; formal analysis, T.A.; resources, V.D.Z., I.B.S., E.Y.-T., J.F.S., W.W., C.L.C. and L.H.K.-V.; A.S.; writing—original draft preparation, V.D.Z. and I.B.S.; writing—review and editing, V.D.Z., I.B.S., E.Y.-T., J.F.S., W.W., C.L.C., L.H.K.-V., A.S. and T.A.; supervision, V.D.Z. All authors have read and agreed to the published version of the manuscript.
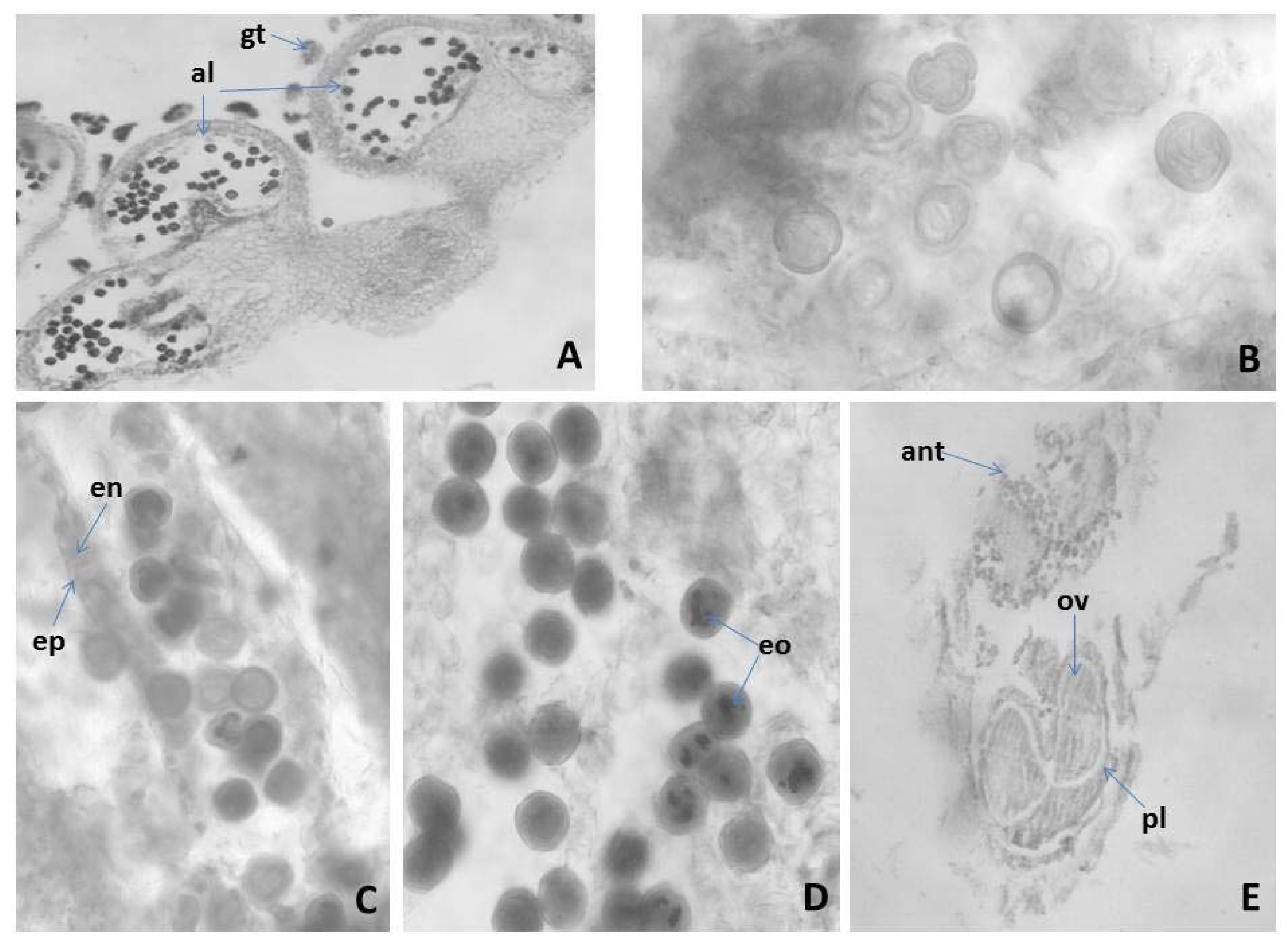
Figure 1.
Anther and development of the male gametophyte revealed after light microscopy observations: (A) tetrasporangiate anther; (B) two celled mature pollen grains; (C) mature pollen grains and anther wall with epidermis and fibrous endothecium; (D) mature pollen grains; (E) flower with anther and ovary; al—anther locule, gt—glandular trihomes, ep—epidermis, en—endothecium, eo—essential oil, pl—pistil, ant—anther, ov—ovule Magnification: (A,E) (×100); (B–D) (×400).
Figure 1.
Anther and development of the male gametophyte revealed after light microscopy observations: (A) tetrasporangiate anther; (B) two celled mature pollen grains; (C) mature pollen grains and anther wall with epidermis and fibrous endothecium; (D) mature pollen grains; (E) flower with anther and ovary; al—anther locule, gt—glandular trihomes, ep—epidermis, en—endothecium, eo—essential oil, pl—pistil, ant—anther, ov—ovule Magnification: (A,E) (×100); (B–D) (×400).
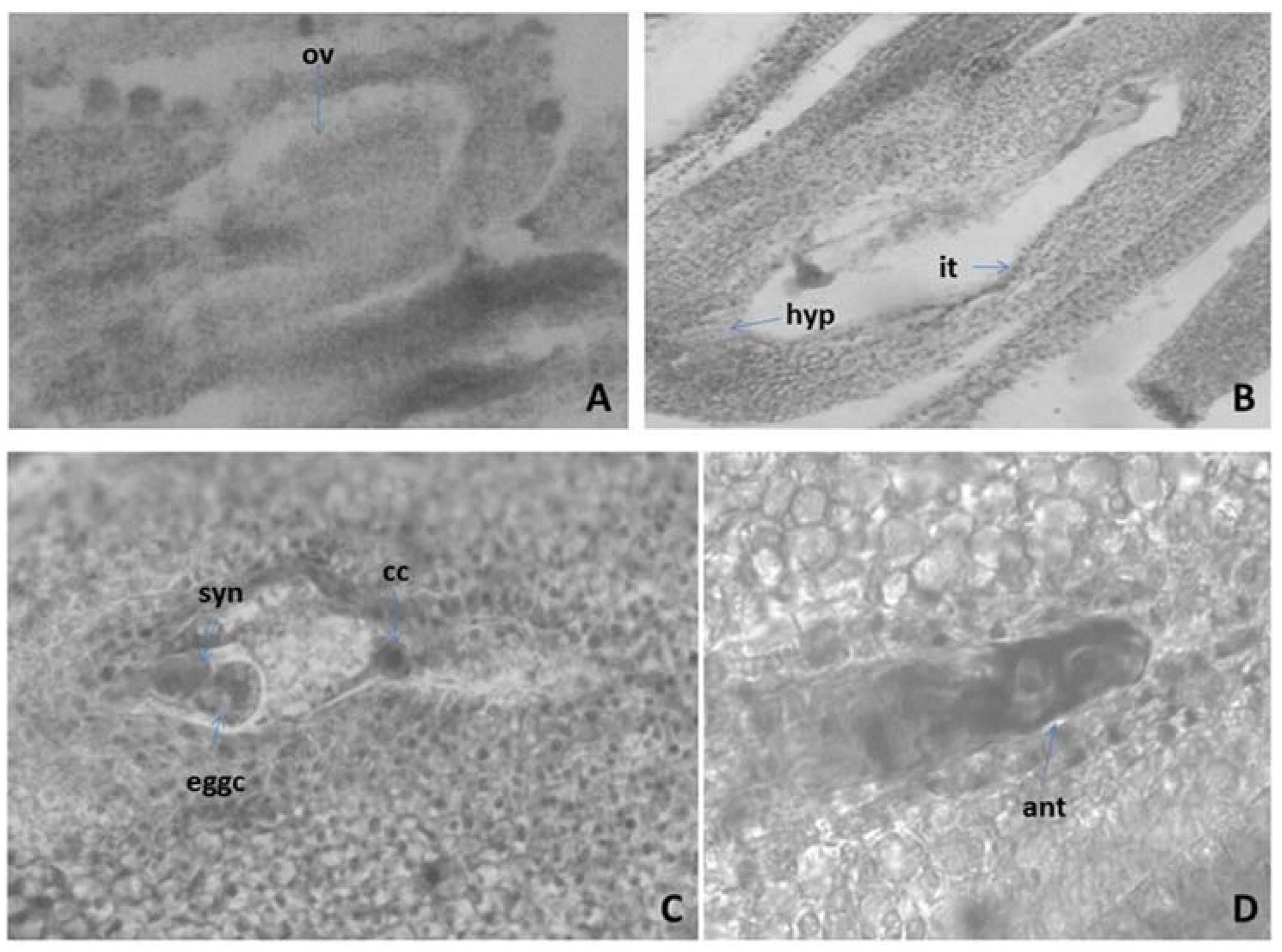
Figure 2.
Ovule and development of the female gametophyte revealed after light microscopy observations: (A) Anatropous unitegmic ovule; (B) Mature Polygonum-type embryo sac (ES); (C) Mature ES with egg apparatus and central cell; (D) Antipodals in the ES—cavity ov—ovule, hyp—hypostase it—integumental tapetum (endothelium), eggc—egg cell, syn—synergids, cc—central cell, ant—antipodal. Magnification: (A,B) (×100); (C,D) (×400).
Figure 2.
Ovule and development of the female gametophyte revealed after light microscopy observations: (A) Anatropous unitegmic ovule; (B) Mature Polygonum-type embryo sac (ES); (C) Mature ES with egg apparatus and central cell; (D) Antipodals in the ES—cavity ov—ovule, hyp—hypostase it—integumental tapetum (endothelium), eggc—egg cell, syn—synergids, cc—central cell, ant—antipodal. Magnification: (A,B) (×100); (C,D) (×400).
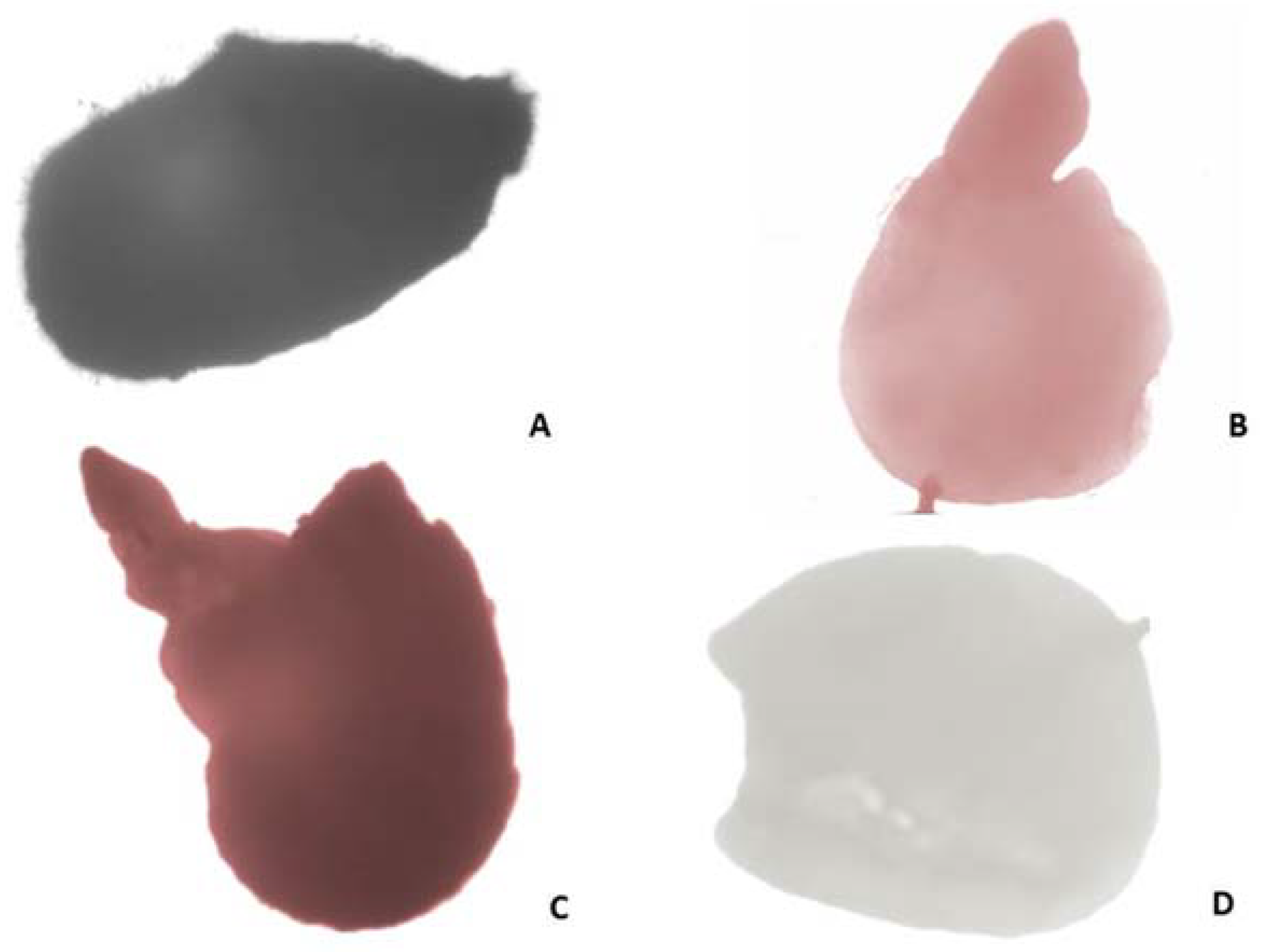
Figure 3.
Estimation of seed viability according to Tetrazolium test: (A) Entire seed; (B,C) viable embryos: pink colored/B/, stained in dark red/C/; (D) unstained non-viable embryo.
Figure 3.
Estimation of seed viability according to Tetrazolium test: (A) Entire seed; (B,C) viable embryos: pink colored/B/, stained in dark red/C/; (D) unstained non-viable embryo.
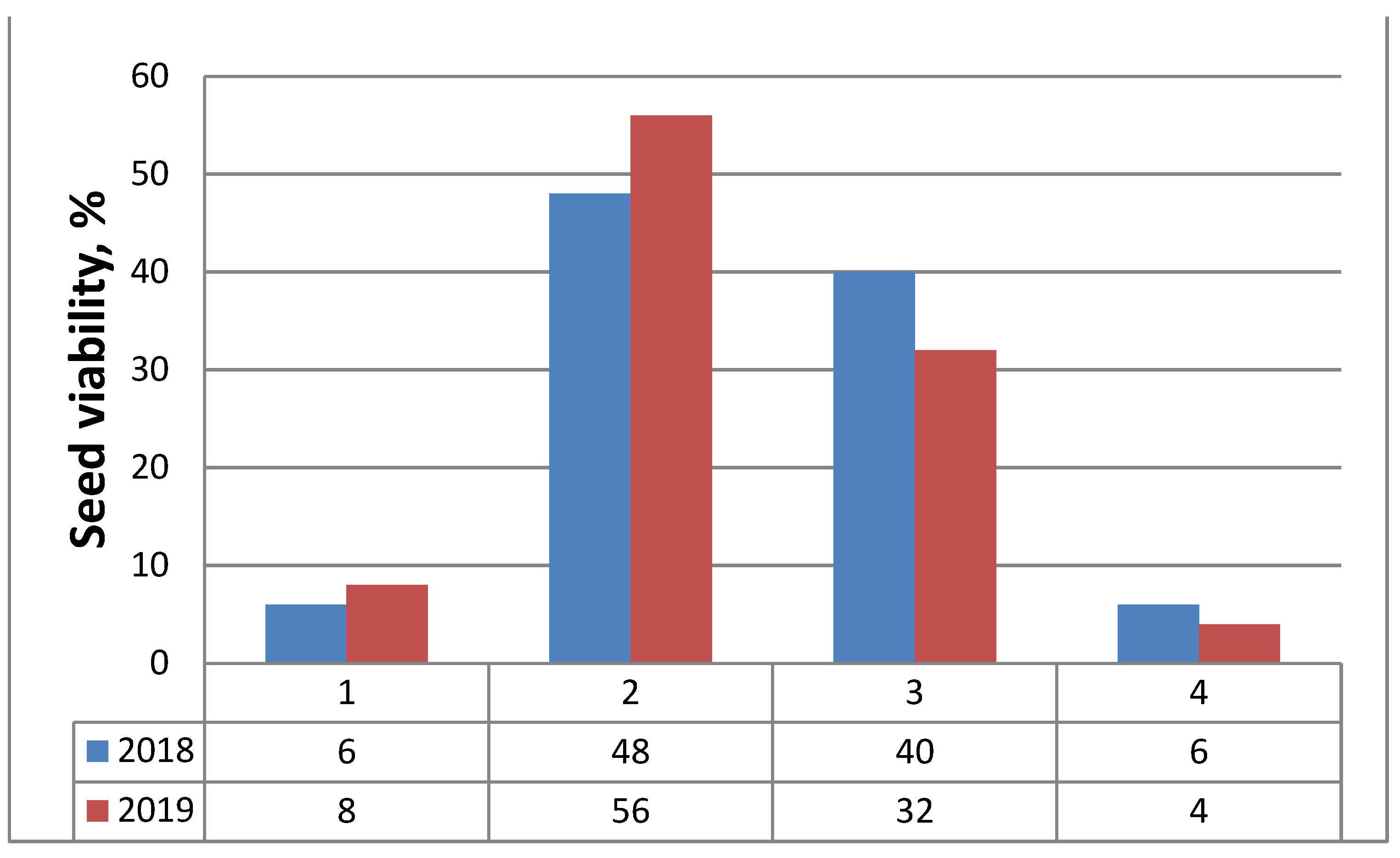
Figure 4.
Frequency of seeds (embryos) viability (%) assessed by tetrazolium test in classes according to the color patterns of tested embryos for two consecutive years (2018, 2019): 1—Class I, viable embryos (stained in red); 2—Class II, viable embryos (pink colored embryos); 3—Class III, non-viable embryos (not stained); 4—Class IV, non-viable (empty) seeds.
Figure 4.
Frequency of seeds (embryos) viability (%) assessed by tetrazolium test in classes according to the color patterns of tested embryos for two consecutive years (2018, 2019): 1—Class I, viable embryos (stained in red); 2—Class II, viable embryos (pink colored embryos); 3—Class III, non-viable embryos (not stained); 4—Class IV, non-viable (empty) seeds.
Figure 5.
The leaves and calyx surfaces revealed after scanning electron microscopy (SEM) observations; (A) calyx inner surface with non-glandular trichomes; (B) calyx outer surface with non-glandular trichomes; (C) leaves surfaces with non-glandular and glandular trichomes; (D) close up of calix outer surfaces with non-glandular and glandular trichomes; (E) surfaces of non-glandular trichomes; (F) glandular peltate trichome, calyx; (G) glandular capitate trichome with some non-glandular trichomes; (H) leaves surfaces with non-glandular and glandular capitate trichomes.
Figure 5.
The leaves and calyx surfaces revealed after scanning electron microscopy (SEM) observations; (A) calyx inner surface with non-glandular trichomes; (B) calyx outer surface with non-glandular trichomes; (C) leaves surfaces with non-glandular and glandular trichomes; (D) close up of calix outer surfaces with non-glandular and glandular trichomes; (E) surfaces of non-glandular trichomes; (F) glandular peltate trichome, calyx; (G) glandular capitate trichome with some non-glandular trichomes; (H) leaves surfaces with non-glandular and glandular capitate trichomes.
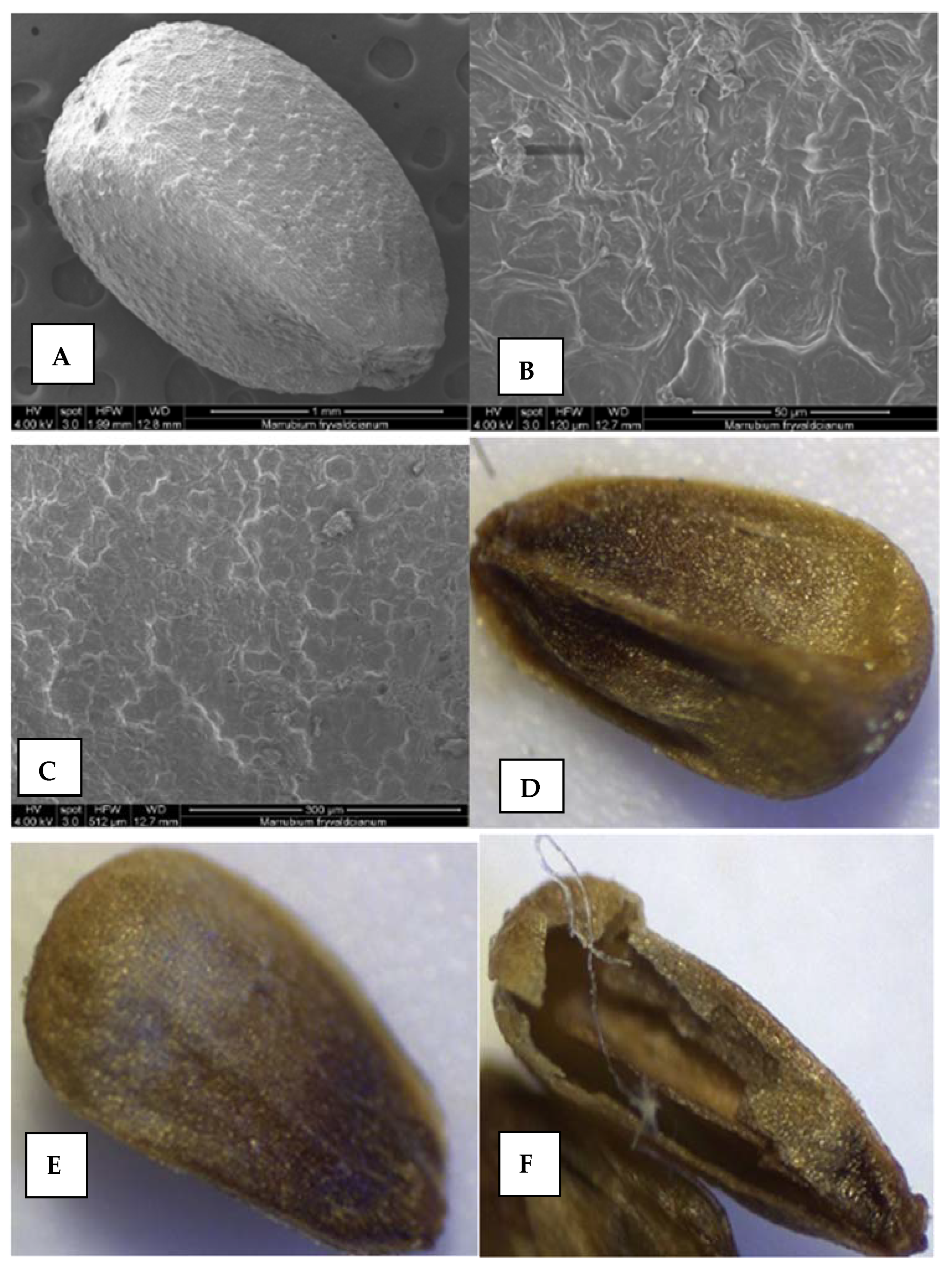
Figure 6.
The nutlets and pollen surfaces revealed after scanning electron microscopy (SEM) and light microscopy (LM) observations. (A–F) nutlet; (G–I) pollen surfaces; (I) polar view.
Figure 6.
The nutlets and pollen surfaces revealed after scanning electron microscopy (SEM) and light microscopy (LM) observations. (A–F) nutlet; (G–I) pollen surfaces; (I) polar view.
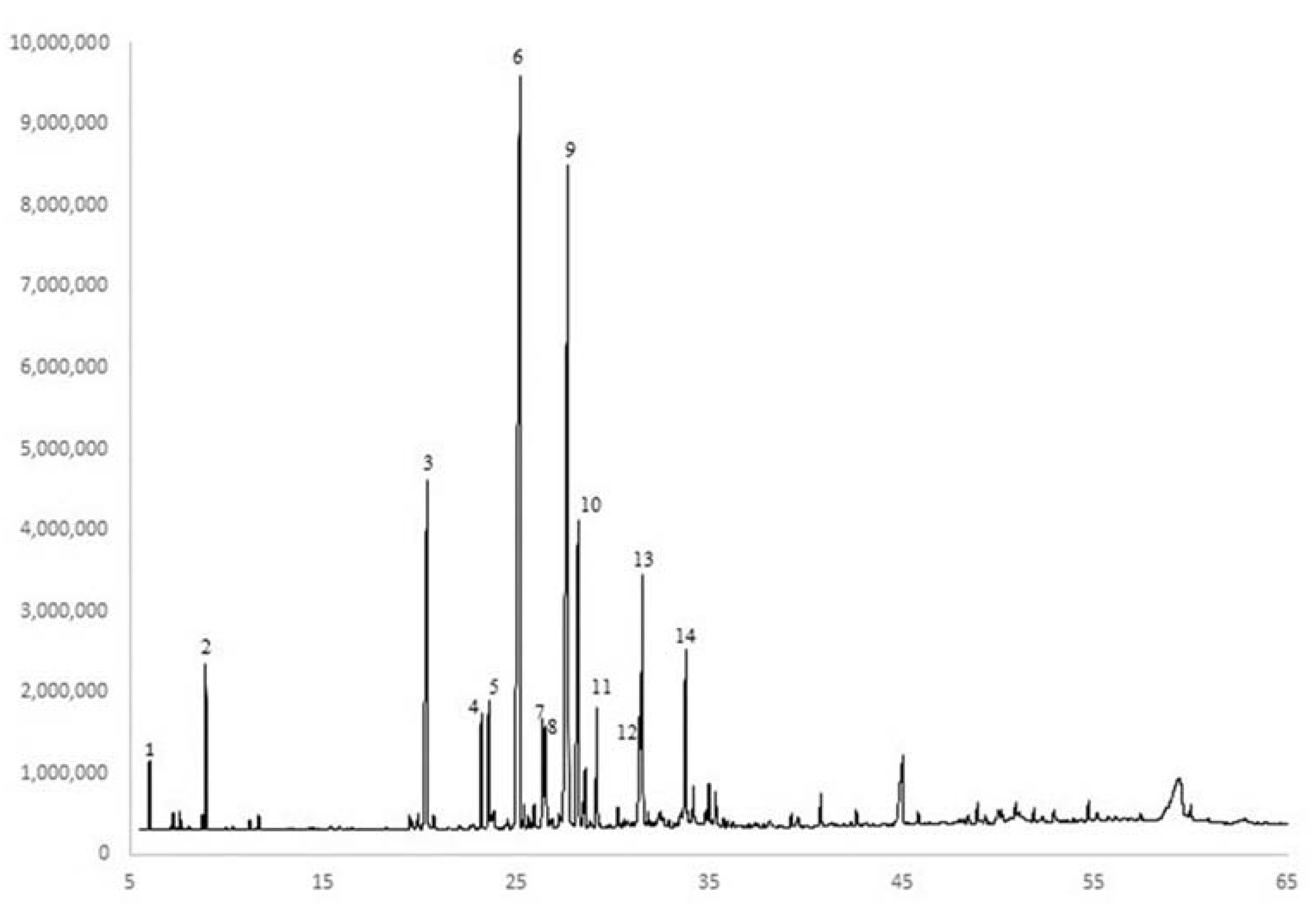
Figure 7.
GC-FID chromatogram of a representative M. friwaldskyanum essential oil with major constituents labeled. 1—α-pinene; 2—D-limonene; 3—carvacrol; 4—α-copaene; 5—β-bourbonene; 6—(E)-caryophyllene; 7—α-humulene; 8—(Z)-β-farnesene; 9—germacrene D; 10—bicylcogermacrene; 11—δ-cadinene; 12—spathulenol; 13—caryophyllene oxide; 14—τ-muurolol.
Figure 7.
GC-FID chromatogram of a representative M. friwaldskyanum essential oil with major constituents labeled. 1—α-pinene; 2—D-limonene; 3—carvacrol; 4—α-copaene; 5—β-bourbonene; 6—(E)-caryophyllene; 7—α-humulene; 8—(Z)-β-farnesene; 9—germacrene D; 10—bicylcogermacrene; 11—δ-cadinene; 12—spathulenol; 13—caryophyllene oxide; 14—τ-muurolol.
Table 1.
Mean concentration (%) of α-copaene, (E)-caryophyllene, caryophyllene oxide, and τ-muurolol obtained from the three levels of harvest stage where the effect of Harvest stage is significant.
Table 1.
Mean concentration (%) of α-copaene, (E)-caryophyllene, caryophyllene oxide, and τ-muurolol obtained from the three levels of harvest stage where the effect of Harvest stage is significant.
| Harvest Stage | α-Copaene | (E)-Caryophyllene | Caryophyllene Oxide | τ-Muurolol |
|---|
| 2–3 pair of leaves | 1.83 a | 41.0 a | 11.81 a | 2.67 a |
| Before flowering | 1.44 ab | 39.3 a | 9.98 ab | 2.80 a |
| Flowering | 1.26 b | 30.8 b | 6.35 b | 1.32 b |
Table 2.
Overall mean concentration (%) of the 6 compounds (β-bourbonene, α-humulene, germacrene D, bicyclogermacrene, δ-cadinene, and spathulenol) where there was no significant difference among the harvest stages.
Table 2.
Overall mean concentration (%) of the 6 compounds (β-bourbonene, α-humulene, germacrene D, bicyclogermacrene, δ-cadinene, and spathulenol) where there was no significant difference among the harvest stages.
| Compound | Overall Mean Concentration (%) |
|---|
| β-bourbonene | 1.10 |
| α-humulene | 2.83 |
| germacrene D | 23.30 |
| bicyclogermacrene | 2.85 |
| δ-cadinene | 1.07 |
| spathulenol | 2.83 |
Table 3.
Mean concentration (%) of D-limonene and bicyclogermacrene obtained from the four levels of Grinding where the effect of Grinding is significant.
Table 3.
Mean concentration (%) of D-limonene and bicyclogermacrene obtained from the four levels of Grinding where the effect of Grinding is significant.
| Grinding | D-Limonene | Bicyclogermacrene |
|---|
| | ------ (%) is the GC-FID area 1 ----- |
| Whole plant | 1.88 b | 9.14 a |
| Grinded/without water | 0.24 d | 6.93 ab |
| Fresh/ground with water | 1.08 c | 3.55 b |
| Ground/immediately extracted | 3.25 a | 6.02 ab |
Table 4.
Germination energy (%) and germination (%) of M. friwaldskyanum.
Table 4.
Germination energy (%) and germination (%) of M. friwaldskyanum.
| Variants | Germination Energy (%) ± SD | Germination (%) ± SD |
|---|
| Variant 1 (N) | 24.0 ± 1.0 | 48.0 ± 2.0 |
| Variant 2 (WR7:B1) | 26.0 ± 1.0 | 56.0 ± 2.1 |
| Variant 3 (WR4:B1) | 26.0 ± 1.0 | 52.0 ± 1.0 |
| Variant 4 (W) | 27.0 ± 0.6 | 58.0 ± 2.0 |
Table 5.
Phenological stage, samples size (g), moisture (%), and oil yield (%) of Marrubium friwaldskyanum in Bulgaria.
Table 5.
Phenological stage, samples size (g), moisture (%), and oil yield (%) of Marrubium friwaldskyanum in Bulgaria.
| Phenologycal Stage | Sample Size/g | Moisture Content (%) | Yield of EO (% v/w) | Yield of EO (%) |
|---|
| (1) 2–3 pairs of leaves | 97.5 | 9.23 | 0.097 | 0.107 |
| (2) pre-flowering flowering | 86.5 | 9.45 | 0.074 | 0.082 |
| (3) Flowering | 159.5 | 62.19 | 0.012 | 0.034 |
Table 6.
Results from the second experiment; treatments, samples size (g), and oil yield (%) of Marrubium friwaldskyanum in Bulgaria.
Table 6.
Results from the second experiment; treatments, samples size (g), and oil yield (%) of Marrubium friwaldskyanum in Bulgaria.
| Treatments | Sample Size (g) | Water
(mL) | Yield of EO, % Volume/Weight |
|---|
| (1) Fresh samples, whole plants, without grinding | 200 | 1100 | 0.05 |
| (2) Fresh, grinding without water | 200 | 1100 | 0.03 |
| (3) Fresh, ground with water | 200 | 1400 | 0.04 |
| (4) Fresh, ground with water, macerated with 0.1% Tween® 20 for 12 h prior to the extraction | 200 | 1400 | 0.04 |
| (5) Fresh, ground with water and macerated with 0.1% Tween® 20 and immediately extracted | 200 | 1400 | 0.03 |
Table 7.
LED bulb specifications.
Table 7.
LED bulb specifications.
| Light Colour | Technical Specification |
|---|
| Power (W) | Tension (V) | Wavelength nm | Light Flux |
|---|
| Red/blue 7:1 | 200 W | 220 V (85−265 V) | 660 nm: 460 nm, (64 diode) of 3 W | 50 sm–250 µmol/m2s |
| Red/blue 4:1 | 200 W | 220 V (85–265 V) | 660 nm: 460 nm, (64diode) of 3 W; 730 nm-2 pcs; | 50 sm–250 µmol/m2s |
| White | 200 W | 220 V (85–265 V) | 660 nm-26 pcs; 630 nm-16 pcs460 nm-9 pcs; 440 nm-6 pcs; 610 nm-2 pcs; 380 nm-1 pcs; 3500 k-2 pcs; (64 diode) of 3 W | 50 sm–250 µmol/m2s |
| daylight | | | | 300 µmol/m2s–2000 µmol/m2s |


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}