
Effect of Cryopreservation on Proteins from the Ubiquitous Marine Dinoflagellate Breviolum sp. (Family Symbiodiniaceae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
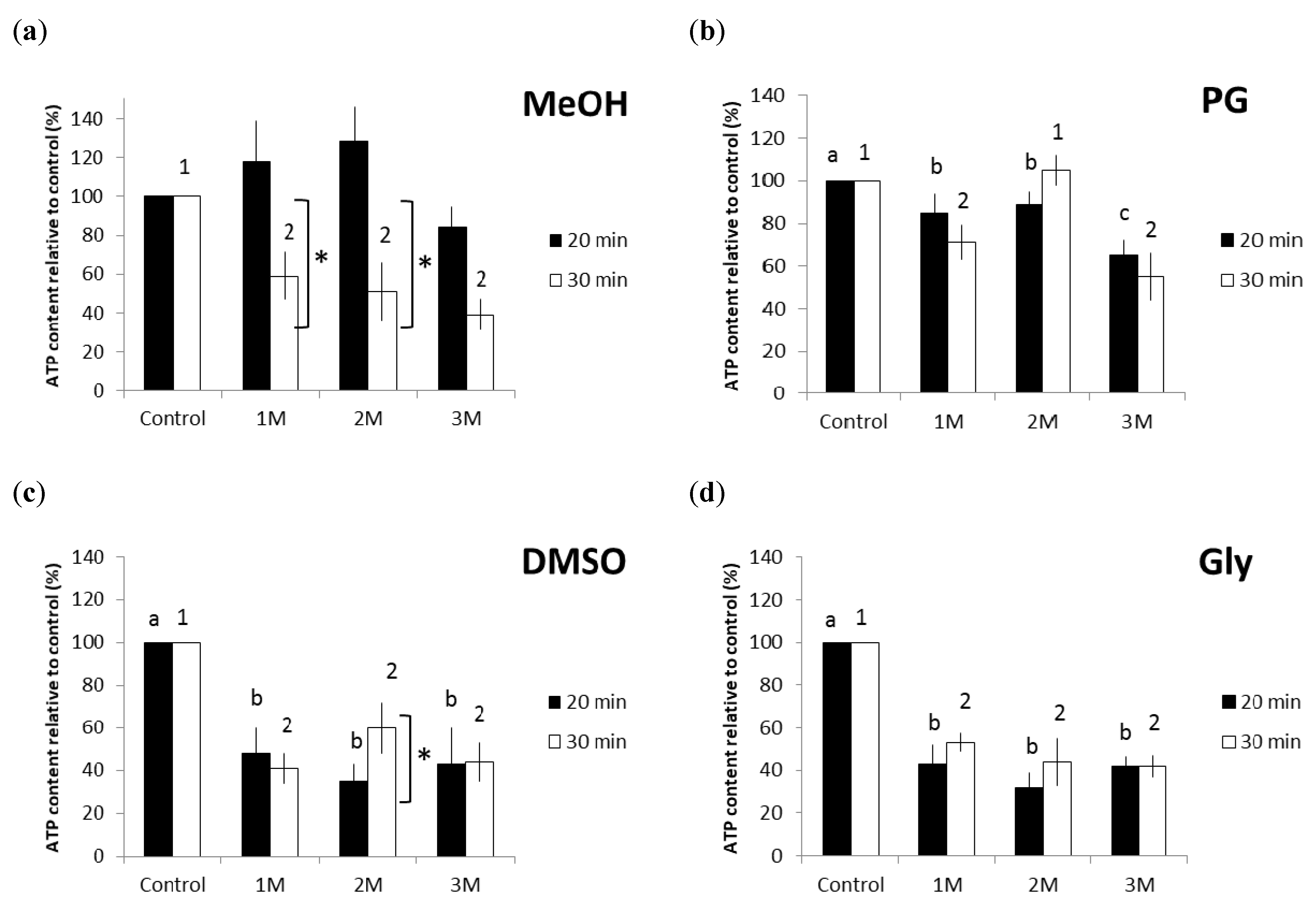
2.1. Effect of Equilibration Time and Concentration of CPAs on the Viability of Frozen-Thawed Breviolum
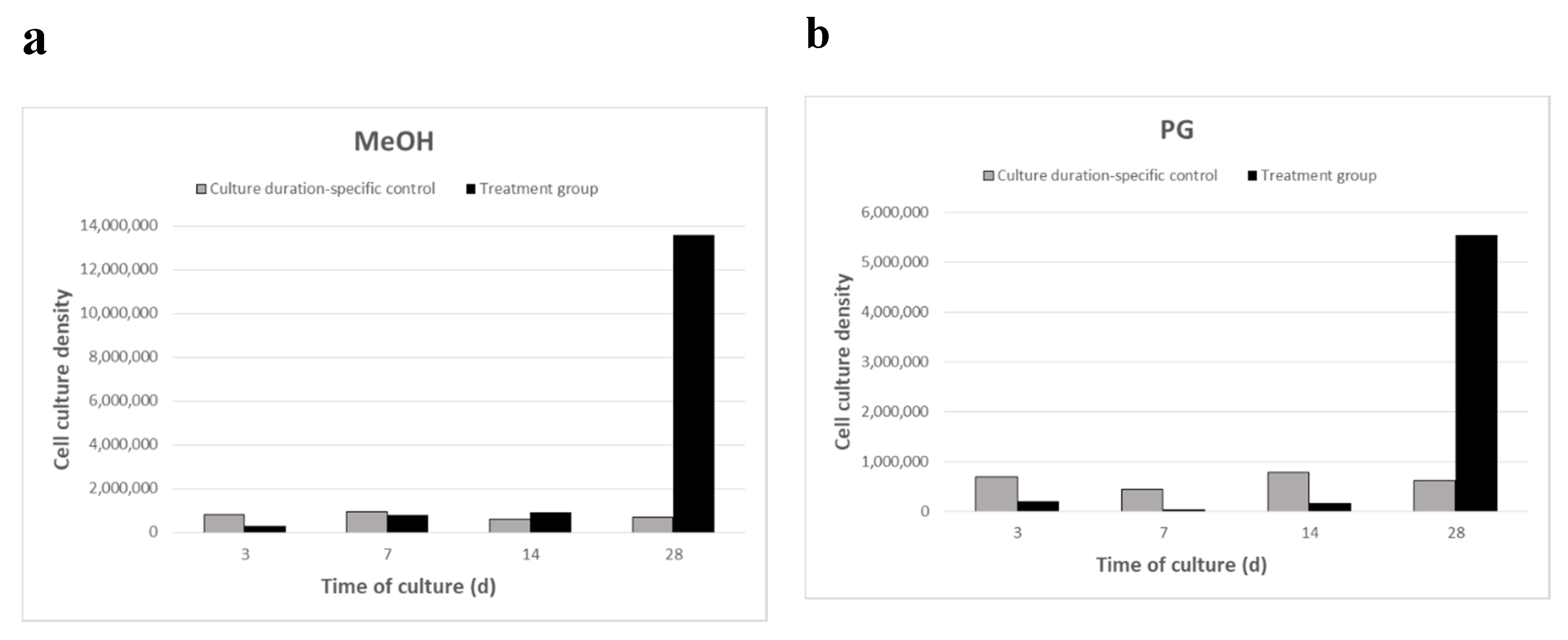
2.2. Breviolum Growth Rate after Four Culture Periods
2.3. Total Protein Expression Pattern of Breviolum
2.4. LHP and RFP Protein Expression Pattern of Breviolum
3. Discussion
4. Materials and Methods
4.1. Symbiodiniaceae Identification
4.2. Culture of Breviolum
4.3. Preparation of Artificial Sea Water
4.4. Renewal of Culture Medium
4.5. ATP Bioassay
4.6. Two-Step Freezing and Culture of Post-Thawed Breviolum
4.7. Protein Extraction
4.8. SDS-PAGE and Western Blotting
4.9. Protein Expression Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsai, S.; Lin, C. Advantages and Applications of Cryopreservation in Fisheries Science. Braz. Arch. Biol. Technol. 2012, 55, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.; Fletcher, R.L. Cryopreservation of eukaryotic algae—A review of methodologies. J. Appl. Phycol. 1998, 10, 481–501. [Google Scholar] [CrossRef]
- Ricaurte, M.; Schizas, N.V.; Ciborowski, P.; Boukli, N.M. Proteomic analysis of bleached and unbleached Acropora palmata, a threatened coral species of the Caribbean. Mar. Pollut. Bull. 2016, 107, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Baumann, J.H.; Davies, S.W.; Aichelman, H.E.; Castillo, K.D. Coral Symbiodinium Community Composition Across the Belize Mesoamerican Barrier Reef System is Influenced by Host Species and Thermal Variability. Microb. Ecol. 2018, 75, 903–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gates, R.D.; Baghdasarian, G.; Muscatine, L. Temperature stress causes host cell detachment in symbiotic cnidarians: Implications for coral bleaching. Biol. Bull. 1992, 182, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Weis, V.M. Cellular mechanisms of Cnidarian bleaching: Stress causes the collapse of symbiosis. J. Exp. Biol. 2008, 211, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Cirino, L.; Wen, Z.-H.; Hsieh, K.; Huang, C.-L.; Leong, Q.L.; Wang, L.-H.; Chen, C.-S.; Daly, J.; Tsai, S.; Lin, C. First instance of settlement by cryopreserved coral larvae in symbiotic association with dinoflagellates. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Lin, C.; Tsai, S. Fifteen years of coral cryopreservation. Platax 2020, 17, 53–76. [Google Scholar]
- Di Genio, S.; Wang, L.H.; Meng, P.J.; Tsai, S.; Lin, C. Symbio-Cryobank: Towards the development of a cryogenic archive for the coral reef dinoflagellate symbiont Symbiodiniaceae. Biopreserv. Biobank. 2020, 19, 91–93. [Google Scholar] [CrossRef]
- Zhao, Y. Cryopreservation of the Gorgonian Coral Endosymbiont Symbiodinium Using Vitrification and Programmable Freezing. Master’s Thesis, National Dong Hwa University, Shoufeng, Taiwan, 2017; pp. 1–70. [Google Scholar]
- Chong, G.; Tsai, S.; Wang, L.-H.; Huang, C.-Y.; Lin, C. Cryopreservation of the gorgonian endosymbiont Symbiodinium. Sci. Rep. 2016, 6, 18816. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Vázquez, L.Z.; Newberger, N.C.; Kerr, R.G. Cryopreservation of the dinoflagellate symbiont of the octocoral Pseudopterogorgia elisabethae. Mar. Biol. 2007, 152, 549–556. [Google Scholar] [CrossRef]
- Hagedorn, M.; Carter, V.L. Seasonal Preservation Success of the Marine Dinoflagellate Coral Symbiont, Symbiodinium sp. PLoS ONE 2015, 10, e0136358. [Google Scholar] [CrossRef]
- Zhang, Q.; Cong, Y.; Qu, S.; Luo, S.; Yang, G. Cryopreservation of gametophytes of Laminaria japonica (Phaeophyta) using encapsulation-dehydration with two-step cooling method. J. Ocean Univ. China 2008, 7, 65–71. [Google Scholar] [CrossRef]
- Liu, J.; Woods, E.J.; Agca, Y.; Critser, E.S.; Critser, J.K. Cryobiology of rat embryos II: A theoretical model for the development of interrupted slow freezing procedures. Biol. Reprod. 2000, 63, 1303–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viveiros, A.; Lock, E.; Woelders, H.; Komen, J. Influence of Cooling Rates and Plunging Temperatures in an Interrupted Slow-Freezing Procedure for Semen of the African Catfish, Clarias gariepinus. Cryobiology 2001, 43, 276–287. [Google Scholar] [CrossRef]
- Morris, G.J.; Canning, C.E. The Cryopreservation of Euglena gracilis. J. Gen. Microbiol. 1978, 108, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.; Yen, W.; Chavanich, S.; Viyakarn, V.; Lin, C. Development of Cryopreservation Techniques for Gorgonian (Junceella juncea) Oocytes through Vitrification. PLoS ONE 2015, 10, e0123409. [Google Scholar] [CrossRef]
- Chong, G.; Kuo, F.-W.; Tsai, S.; Lin, C. Validation of reference genes for cryopreservation studies with the gorgonian coral endosymbiont Symbiodinium. Sci. Rep. 2017, 7, 39396. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wu, G.; Huang, X.; Liu, S.; Cong, B. Validation of housekeeping genes for gene expression studies in an ice alga Chlamydomonas during freezing acclimation. Extremophiles 2012, 16, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Prieto, R.; Trench, R. Acclimation and adaptation to irradiance in symbiotic dinoflagellates. II. Response of chlorophyll-protein complexes to different photon-flux densities. Mar. Biol. 1997, 130, 23–33. [Google Scholar] [CrossRef]
- Takahashi, S.; Whitney, S.; Itoh, S.; Maruyama, T.; Badger, M. Heat stress causes inhibition of the de novo synthesis of antenna proteins and photobleaching in cultured Symbiodinium. Proc. Natl. Acad. Sci. USA 2008, 105, 4203–4208. [Google Scholar] [CrossRef] [Green Version]
- Prézelin, B.B.; Haxo, F.T. Purification and characterization of peridinin-chlorophyll a-proteins from the marine dinoflag-ellates Glenodinium sp. and Gonyaulax polyedra. Planta 1976, 128, 133–141. [Google Scholar] [CrossRef]
- Gross, L.A.; Baird, G.S.; Hoffman, R.C.; Baldridge, K.K.; Tsien, R.Y. The structure of the chromophore within DsRed, a red fluorescent protein from coral. Proc. Natl. Acad. Sci. USA 2000, 97, 11990–11995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevis, B.J.; Glick, B.S. Rapidly maturing variants of the Discosoma red fluorescent protein (DsRed). Nat. Biotechnol. 2002, 20, 83–87. [Google Scholar] [CrossRef]
- Huang, K.-J.; Huang, Z.-Y.; Lin, C.-Y.; Wang, L.-H.; Chou, P.-H.; Chen, C.-S.; Li, H.-H. Generation of clade- and symbiont-specific antibodies to characterize marker molecules during Cnidaria-Symbiodinium endosymbiosis. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stauber, E.J.; Fink, A.; Markert, C.; Kruse, O.; Johanningmeier, U.; Hippler, M. Proteomics of Chlamydomonas reinhardtii Light-Harvesting Proteins. Eukaryot. Cell 2003, 2, 978–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldt, L.; Yellowlees, D.; Leggat, W. Hyperdiversity of Genes Encoding Integral Light-Harvesting Proteins in the Dinoflagellate Symbiodinium sp. PLoS ONE 2012, 7, e47456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, M.; Carter, V.; Leong, J.; Kleinhans, F. Physiology and cryosensitivity of coral endosymbiotic algae (Symbiodinium). Cryobiology 2010, 60, 147–158. [Google Scholar] [CrossRef]
- Lin, C.; Thongpoo, P.; Juri, C.; Wang, L.-H.; Meng, P.-J.; Kuo, F.-W.; Tsai, S. Cryopreservation of a Thermotolerant Lineage of the Coral Reef Dinoflagellate Symbiodinium. Biopreserv. Biobank. 2019, 17, 520–529. [Google Scholar] [CrossRef]
- Cañavate, J.P.; Lubian, L.M. Tolerance of six marine microalgae to the cryoprotectants dimethyl sulfoxide and methanol. J. Phycol. 1994, 30, 559–565. [Google Scholar] [CrossRef]
- Mitbavkar, S.; Anil, A.C. Cell damage and recovery in cryopreserved microphytobenthic diatoms. Cryobiology 2006, 53, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Tsuru, S. Preservation of marine and fresh water algae by means of freezing and freeze-drying. Cryobiology 1973, 10, 445–452. [Google Scholar] [CrossRef]
- Lin, C.; Chong, G.; Wang, L.; Kuo, F.; Tsai, S. Use of luminometry and flow cytometry for evaluating the effects of cryoprotectants in the gorgonian coral endosymbiont Symbiodinium. Phycol. Res. 2019, 67, 320–326. [Google Scholar] [CrossRef]
- Gwo, J.C.; Chiu, J.Y.; Chou, C.C.; Cheng, H.Y. Cryopreservation of a marine microalga, Nannochloropsis oculata (Eustigmatophyceae). Cryobiology 2005, 50, 338–343. [Google Scholar] [CrossRef]
- Morris, G. Cryopreservation: An Introduction to Cryopreservation in Culture Collections; Institute of Terrestrial Ecology: Cambridge, UK, 1981; pp. 1–27. [Google Scholar]
- Yang, D.; Li, W. Methanol-Promoted Lipid Remodelling during Cooling Sustains Cryopreservation Survival of Chlamydomonas reinhardtii. PLoS ONE 2016, 11, e0146255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G. Cryopreservation of 250 strains of Chlorococcales by the method of two-step cooling. Br. Phycol. J. 1978, 13, 15–24. [Google Scholar] [CrossRef]
- Fenwick, C.; Day, J.G. Cryopreservation of Tetraselmis suecica cultured under different nutrients regimes. J. Appl. Phycol. 1992, 4, 105–109. [Google Scholar] [CrossRef]
- Chong, G.; Tsai, S.; Lin, C. Factors responsible for successful cryopreservation of algae. J. Fish Soc. Taiwan. 2016, 43, 153–162. [Google Scholar]
- Box, J. Cryopreservation of the blue-green alga Microcystis aeruginosa. Eur. J. Phycol. 1988, 23, 385–386. [Google Scholar] [CrossRef]
- Hubálek, Z. Protectants used in the cryopreservation of microorganisms. Cryobiology 2003, 46, 205–229. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, J.; Tervit, R.; Roberts, R.; Adamson, J.; Adams, S.; Decker, M. Cryopreservation of economically valuable marine micro-algae in the classes Bacillariophyceae, Chlorophyceae, Cyanophyceae, Dinophyceae, Haptophyceae, Prasinophyceae, and Rhodophyceae. Cryobiology 2006, 52, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Tsai, S. The effect of chilling and cryoprotectants on hard coral (Echinopora spp.) oocytes during short-term low temperature preservation. Theriogenology 2012, 77, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Zhang, S.C.; Liu, X.Z.; Xu, Y.J.; Hu, J.H.; Xu, Y.Y.; Li, J.; Chen, S.L. Toxicity and protective efficiency of cryo-protectants to flounder (Paralichthys olivaceus) embryos. Theriogenology. 2005, 63, 763–773. [Google Scholar] [CrossRef]
- Chong, G.; Tsai, S.; Lin, C. Cryopreservation and its molecular impacts on microorganisms. J. Fish Soc. Taiwan 2016, 43, 263–272. [Google Scholar]
- Zilli, L.; Beirão, J.; Schiavone, R.; Herraez, P.; Gnoni, A.; Vilella, S. Comparative Proteome Analysis of Cryopreserved Flagella and Head Plasma Membrane Proteins from Sea Bream Spermatozoa: Effect of Antifreeze Proteins. PLoS ONE 2014, 9, e99992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voolstra, C.R.; Schnetzer, J.; Peshkin, L.; Randall, C.J.; Szmant, A.M.; Medina, M. Effects of temperature on gene expression in embryos of the coral Montastraea faveolata. BMC Genom. 2009, 10, 627. [Google Scholar] [CrossRef] [Green Version]
- Weston, A.J.; Dunlap, W.C.; Shick, J.M.; Klueter, A.; Iglic, K.; Vukelic, A.; Starcevic, A.; Ward, M.; Wells, M.L.; Trick, C.G.; et al. A Profile of an Endosymbiont-enriched Fraction of the Coral Stylophora pistillata Reveals Proteins Relevant to Microbial-Host Interactions. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Shaner, N.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef]
- Shikina, S.; Chiu, Y.-L.; Chung, Y.-J.; Chen, C.-J.; Lee, Y.-H.; Chang, C.-F. Oocytes express an endogenous red fluorescent protein in a stony coral, Euphyllia ancora: A potential involvement in coral oogenesis. Sci. Rep. 2016, 6, 25868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, A.H.; Bhagooli, R.; Ralph, P.; Takahashi, S. Coral bleaching: The role of the host. Trends Ecol. Evol. 2009, 24, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Grajales, A.; Rodríguez, E. Morphological revision of the genus Aiptasia and the family Aiptasiidae (Cnidaria, Actiniaria, Metridioidea). Zootaxa 2014, 3826, 55–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, S.R.; Taylor, D.J.; Kinzie, R.A., III; Hidaka, M.; Sakai, K.; Coffroth, M.A. Molecular phylogeny of symbiotic di-noflagel-lates inferred from partial chloroplast large subunit (23S)-rDNA sequences. Mol. Phylogenet. Evol. 2002, 23, 97–111. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.-H.; Lu, J.-L.; Lo, H.-E.; Tsai, S.; Lin, C. Effect of Cryopreservation on Proteins from the Ubiquitous Marine Dinoflagellate Breviolum sp. (Family Symbiodiniaceae). Plants 2021, 10, 1731. https://doi.org/10.3390/plants10081731
Li H-H, Lu J-L, Lo H-E, Tsai S, Lin C. Effect of Cryopreservation on Proteins from the Ubiquitous Marine Dinoflagellate Breviolum sp. (Family Symbiodiniaceae). Plants. 2021; 10(8):1731. https://doi.org/10.3390/plants10081731
Chicago/Turabian StyleLi, Hsing-Hui, Jia-Lin Lu, Hui-Esther Lo, Sujune Tsai, and Chiahsin Lin. 2021. "Effect of Cryopreservation on Proteins from the Ubiquitous Marine Dinoflagellate Breviolum sp. (Family Symbiodiniaceae)" Plants 10, no. 8: 1731. https://doi.org/10.3390/plants10081731