
A Comparative Study on Photo-Protective and Anti-Melanogenic Properties of Different Kadsura coccinea Extracts


 , , and
, , and 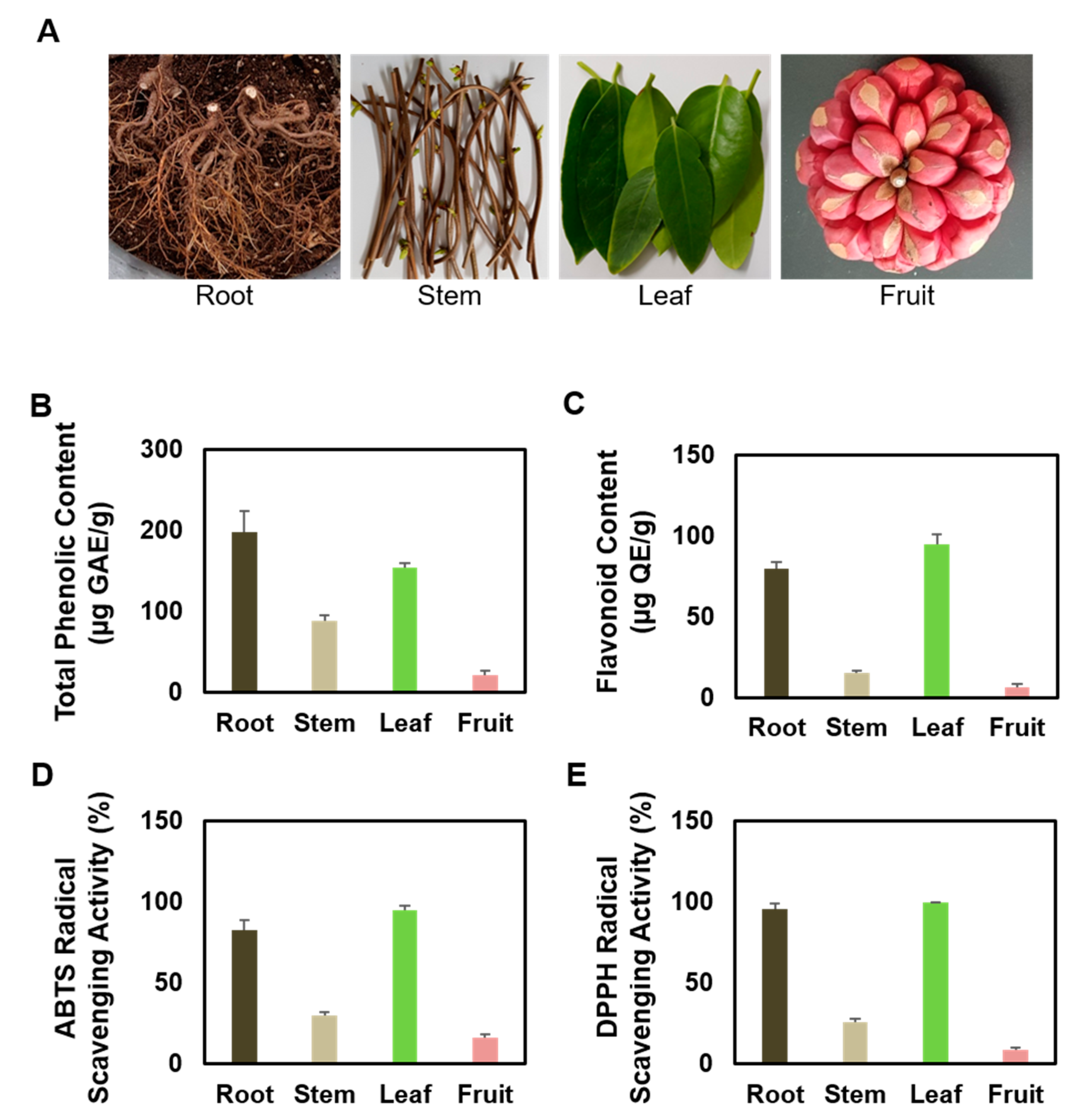{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of KC Extracts
2.2. Total Polyphenolic and Flavonoid Contents of KC Extracts
2.3. DPPH and ABTS Assay
2.4. Cell Culture
2.5. UVA and UVB Irradiation
2.6. Measurement of Cell Viability and Cytotoxicity through CCK-8 and LDH Analysis
2.7. Evaluation of Intracellular ROS Production in Keratinocytes
2.8. Analysis of Apoptosis
2.9. Analysis of Intracellular Melanin Content and Tyrosinase Activity
2.10. Quantitative Real-Time PCR
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
3.1. Comparison of Antioxidant Properties of Several Partial Extracts of KC
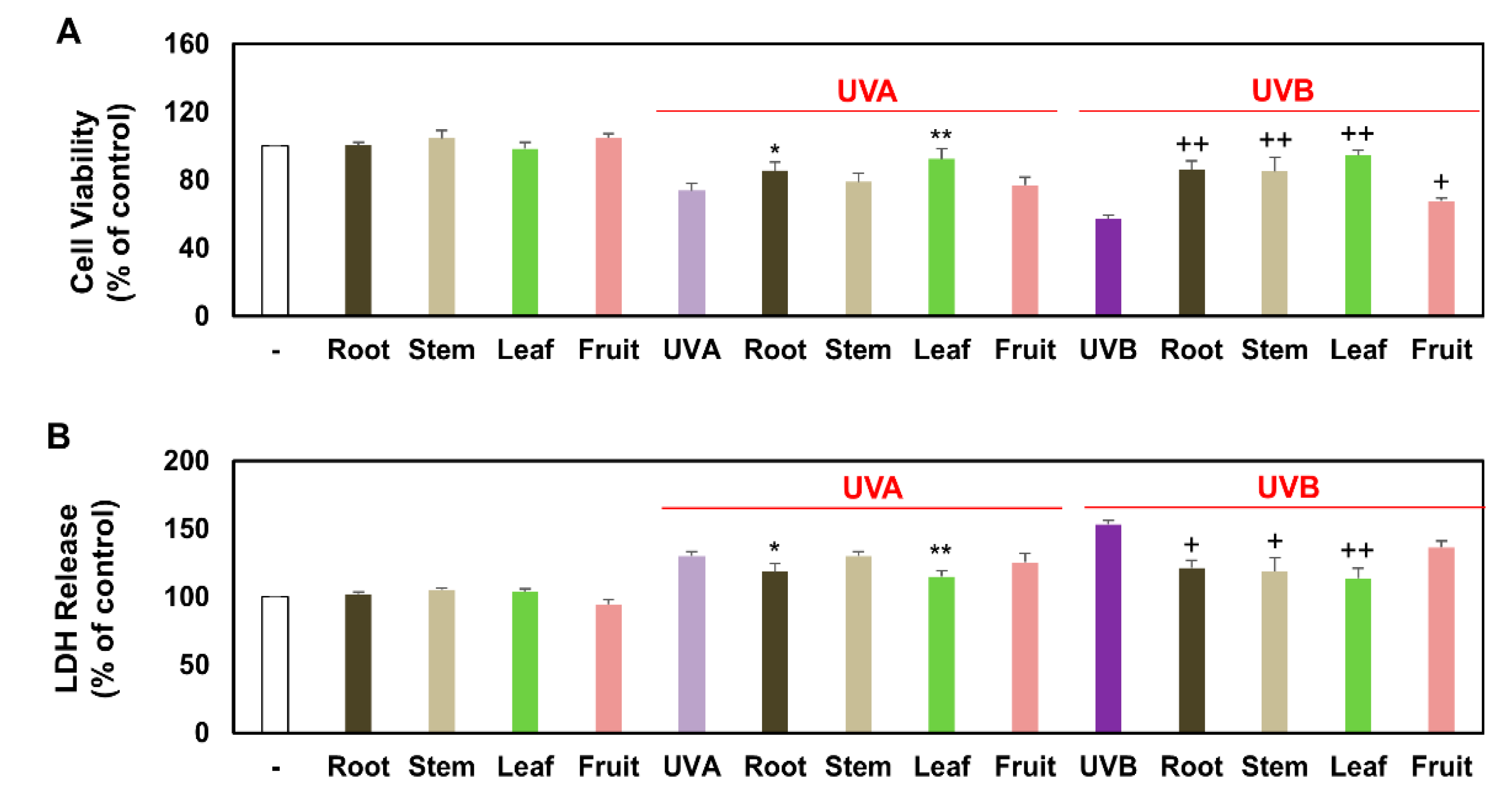
3.2. Comparison of Viable and Damaged Keratinocytes Treated with Several Partial Extracts of KC in the Presence of UVA, UVB, or Non-Irradiation
3.3. Comparison of Intracellular ROS Production in Keratinocytes Treated with Several Partial Extracts of KC in the Presence or Absence of UVA and UVB Irradiation
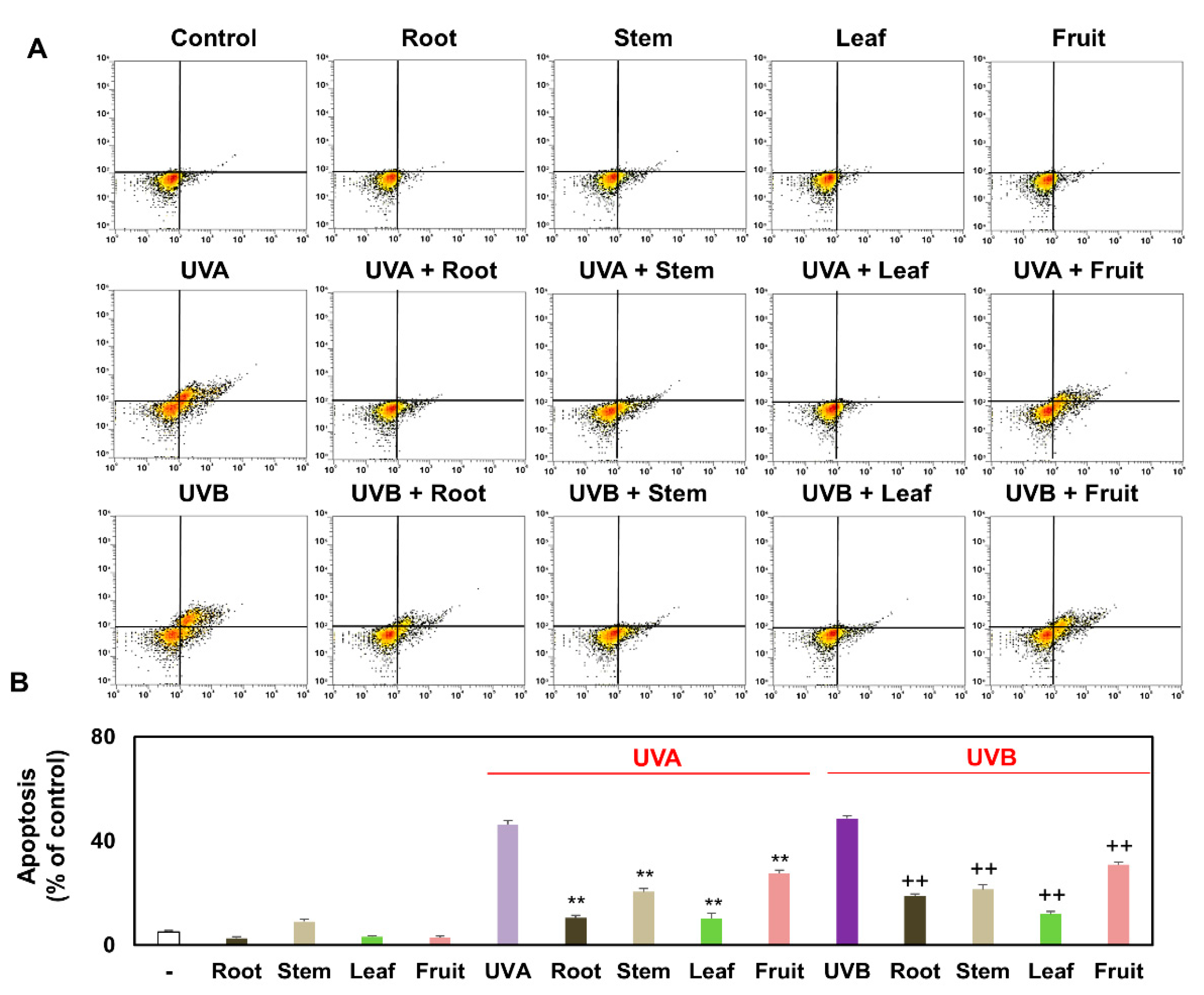
3.4. Comparison of Apoptosis of Keratinocytes Treated with Several Partial Extracts of KC in the Presence or Absence of UVA and UVB Irradiation
3.5. Comparison of Intracellular Melanin Content and Tyrosinase Activity of Melanocytes Treated with Several Partial Extracts of KC in the Presence or Absence of α-MSH Treatment
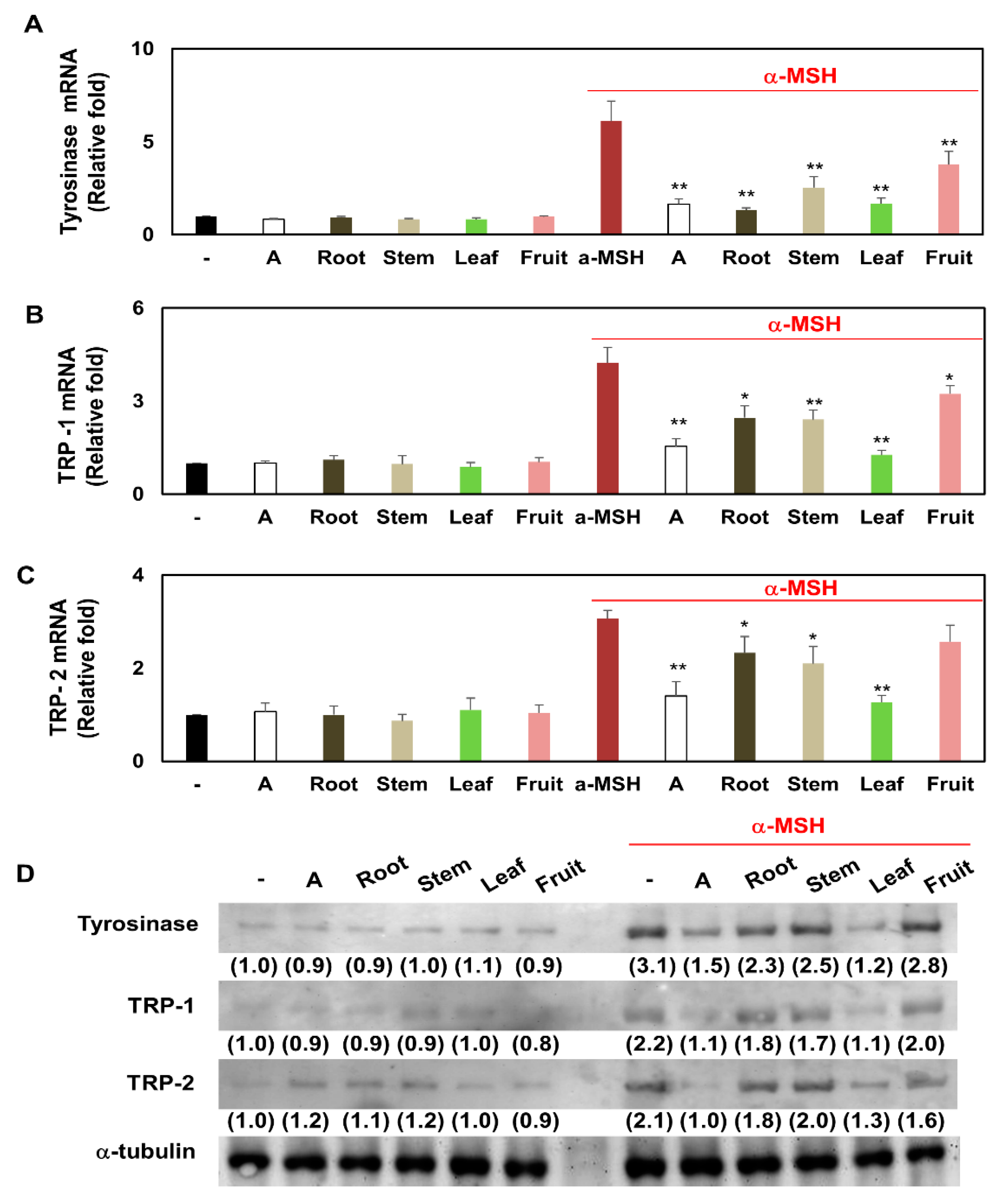
3.6. Comparison of Transcription and Translation Levels of Tyrosinase, TRP-1, and TRP-2 in Melanocytes Treated with Several Partial Extracts of KC in the Presence or Absence of α-MSH Treatment
3.7. Comparison of MITF Expression and CREB Phosphorylation of Melanocytes Treated with Several Partial Extracts of KC in the Presence or Absence of α-MSH Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, J.; Wang, X.A.; Gao, M.Y.; Wei, X.X. The complete chloroplast genome of ‘black tiger 2’ (Kadsura coccinea (lem.) A.C. Smith) in southeast of China and phylogenetic relationshipsAQ1. Mitochondrial DNA B Resour. 2019, 5, 296–297. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wei, J.G.; Wang, K.W.; Luo, J.; Wu, Y.J.; Luo, J.T.; Yang, X.H.; Yang, X.B. Three phytotoxins produced by Neopestalotiopsis clavispora, the causal agent of ring spot on Kadsura coccinea. Microbiol. Res. 2020, 238, 126531. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.H.; Nguyen, D.H.; Choi, J.S.; Park, S.E.; Thuong, P.T.; Min, B.S.; Le, D.D. Chemical constituents from the roots of Kadsura coccinea with their protein tyrosine phosphatase 1B and acetylcholinesterase inhibitory activities. Arch. Pharm. Res. 2020, 43, 204–213. [Google Scholar] [CrossRef]
- Huang, S.Z.; Duan, L.P.; Wang, H.; Mei, W.L.; Dai, H.F. Two New AChE Inhibitors Isolated from Li Folk Herb Heilaohu “Kadsura coccinea” Stems. Molecules 2019, 24, 3628. [Google Scholar] [CrossRef] [Green Version]
- Sritalahareuthai, V.; Aursalung, A.; On-Nom, N.; Temviriyanukul, P.; Charoenkiatkul, S.; Suttisansanee, U. Nutritional composition of conserved Kadsura spp. plants in Northern Thailand. Heliyon 2020, 6, e04451. [Google Scholar] [CrossRef]
- Liu, J.; Wei, X.; Zhang, X.; Qi, Y.; Zhang, B.; Liu, H.; Xiao, P. A Comprehensive Comparative Study for the Authentication of the Kadsura Crude Drug. Front. Pharmacol. 2019, 9, 1576. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Daniyal, M.; Yu, H.; Xie, Q.; Li, B.; Jian, Y.; Man, R.; Wang, S.; Zhou, X.; et al. New Lignans from roots of Kadsura coccinea. Fitoterapia 2019, 139, 104368. [Google Scholar] [CrossRef]
- Wang, P.W.; Hung, Y.C.; Lin, T.Y.; Fang, J.Y.; Yang, P.M.; Chen, M.H.; Pan, T.L. Comparison of the Biological Impact of UVA and UVB upon the Skin with Functional Proteomics and Immunohistochemistry. Antioxidants 2019, 8, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, H.; Li, H.; Ho, Z.Y.; Dai, X.Y.; Chen, Q.; Li, R.; Liang, B.; Zhu, H. Pterostilbene’s protective effects against photodamage caused by UVA/UVB irradiation. Pharmazie 2018, 73, 651–658. [Google Scholar]
- Gęgotek, A.; Ambrożewicz, E.; Jastrząb, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Rutin and ascorbic acid cooperation in antioxidant and antiapoptotic effect on human skin keratinocytes and fibroblasts exposed to UVA and UVB radiation. Arch. Dermatol. Res. 2019, 311, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Jastrząb, A.; Jarocka-Karpowicz, I.; Muszyńska, M.; Skrzydlewska, E. The Effect of Sea Buckthorn (Hippophae rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells. Antioxidants 2018, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marabini, L.; Melzi, G.; Lolli, F.; Dell’Agli, M.; Piazza, S.; Sangiovanni, E.; Marinovich, M. Effects of Vitis vinifera L. leaves extract on UV radiation damage in human keratinocytes (HaCaT). J. Photochem. Photobiol. B 2020, 204, 111810. [Google Scholar] [CrossRef]
- Muzaffer, U.; Paul, V.I.; Prasad, N.R.; Karthikeyan, R.; Agilan, B. Protective effect of Juglans regia L. against ultraviolet B radiation induced inflammatory responses in human epidermal keratinocytes. Phytomedicine 2018, 42, 100–111. [Google Scholar] [CrossRef]
- Maciel, B.; Moreira, P.; Carmo, H.; Gonçalo, M.; Lobo, J.M.S.; Almeida, I.F. Implementation of an in vitro methodology for phototoxicity evaluation in a human keratinocyte cell line. Toxicol In Vitro 2019, 61, 104618. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, J.E.; Kim, T.; Yeom, M.H.; Park, J.S.; di Luccio, E.; Chen, H.; Dong, Z.; Lee, K.W.; Kang, N.J. 7,3’,4’-Trihydroxyisoflavone, a Metabolite of the Soy Isoflavone Daidzein, Suppresses α-Melanocyte-Stimulating Hormone-Induced Melanogenesis by Targeting Melanocortin 1 Receptor. Front. Mol. Biosci. 2020, 7, 577284. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.C.; Chen, X.Z.; Vudhya Gowrisankar, Y.; Yen, H.R.; Chuang, J.Y.; Yang, H.L. The Skin-Whitening Effects of Ectoine via the Suppression of α-MSH-Stimulated Melanogenesis and the Activation of Antioxidant Nrf2 Pathways in UVA-Irradiated Keratinocytes. Antioxidants 2020, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Seo, G.Y.; Ha, Y.; Park, A.H.; Kwon, O.W.; Kim, Y.J. Leathesia difformis Extract Inhibits α-MSH-Induced Melanogenesis in B16F10 Cells via Down-Regulation of CREB Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 536. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, X.; Jiang, R.; Sun, L.; Zhao, D. Vanillic acid in Panax ginseng root extract inhibits melanogenesis in B16F10 cells via inhibition of the NO/PKG signaling pathway. Biosci. Biotechnol. Biochem. 2019, 83, 1205–1215. [Google Scholar] [CrossRef]
- Kim, B.H.; Hong, S.N.; Ye, S.K.; Park, J.Y. Evaluation and Optimization of the Anti-Melanogenic Activity of 1-(2-Cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone Derivatives. Molecules 2019, 24, 1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.J.; Lee, A.K.; Park, Y.J.; Lee, S.; Kang, D.; Jung, Y.S.; Chung, H.Y.; Moon, H.R. (2E,5E)-2,5-Bis(3-hydroxy-4-methoxybenzylidene) cyclopentanone Exerts Anti-Melanogenesis and Anti-Wrinkle Activities in B16F10 Melanoma and Hs27 Fibroblast Cells. Molecules 2018, 23, 1415. [Google Scholar] [CrossRef] [Green Version]
- Azam, M.S.; Kwon, M.; Choi, J.; Kim, H.R. Sargaquinoic acid ameliorates hyperpigmentation through cAMP and ERK-mediated downregulation of MITF in α-MSH-stimulated B16F10 cells. Biomed. Pharmacother. 2018, 104, 582–589. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Oh, S.W.; Park, S.H.; Lee, J.; Yoo, J.A.; Kwon, K.; Park, S.J.; Kim, J.; Yu, E.; Cho, J.Y.; et al. Melanogenic Effects of Maclurin Are Mediated through the Activation of cAMP/PKA/CREB and p38 MAPK/CREB Signaling Pathways. Oxid. Med. Cell Longev. 2019, 2019, 9827519. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.; Kim, Y.; Park, T.J.; Kang, H.Y. Dasatinib, a second-generation tyrosine kinase inhibitor, induces melanogenesis via ERK-CREB-MITF-tyrosinase signaling in normal human melanocytes. Biochem. Biophys. Res. Commun. 2020, 523, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jin, M.L.; Kim, Y.H.; Kim, Y.; Lee, S.J. Aromatic-turmerone inhibits α-MSH and IBMX-induced melanogenesis by inactivating CREB and MITF signaling pathways. Arch. Dermatol. Res. 2011, 303, 737–744. [Google Scholar] [CrossRef]
- Park, S.Y.; Lu, G.; Kim, B.; Song, W.C.; Park, G.; Choi, Y.W. A Comparative Study on Physicochemical, Photocatalytic, and Biological Properties of Silver Nanoparticles Formed Using Extracts of Different Parts of Cudrania tricuspidata. Nanomaterials 2020, 10, 1350. [Google Scholar] [CrossRef] [PubMed]
- Zaid, A.N.; Al Ramahi, R. Depigmentation and Anti-aging Treatment by Natural Molecules. Curr. Pharm. Des. 2019, 25, 2292–2312. [Google Scholar] [CrossRef] [PubMed]
- Desmedt, B.; Courselle, P.; De Beer, J.O.; Rogiers, V.; Grosber, M.; Deconinck, E.; De Paepe, K. Overview of skin whitening agents with an insight into the illegal cosmetic market in Europe. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 943–950. [Google Scholar] [CrossRef]
- Qian, W.; Liu, W.; Zhu, D.; Cao, Y.; Tang, A.; Gong, G.; Su, H. Natural skin-whitening compounds for the treatment of melanogenesis (Review). Exp. Ther. Med. 2020, 20, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, Y.; Tasneem, S.; Hussain, N.; Daniyal, M.; Yuan, H.; Xie, Q.; Liu, B.; Sun, J.; Jian, Y.; et al. Lignans from Tujia Ethnomedicine Heilaohu: Chemical Characterization and Evaluation of Their Cytotoxicity and Antioxidant Activities. Molecules 2018, 23, 2147. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Qi, Y.; Lai, H.; Zhang, J.; Jia, X.; Liu, H.; Zhang, B.; Xiao, P. Genus Kadsura, a good source with considerable characteristic chemical constituents and potential bioactivities. Phytomedicine 2014, 21, 1092–1097. [Google Scholar] [CrossRef]
- Yang, Y.; Hussain, N.; Zhang, L.; Jia, Y.; Jian, Y.; Li, B.; Choudhary, M.I.; Rahman, A.; Wang, W. Kadsura coccinea: A rich source of structurally diverse and biologically important compounds. Chin. Herbal Med. 2020, 12, 214–223. [Google Scholar] [CrossRef]
- Sritalahareuthai, V.; Temviriyanukul, P.; On-Nom, N.; Charoenkiatkul, S.; Suttisansanee, U. Phenolic Profiles, Antioxidant, and Inhibitory Activities of Kadsura heteroclita (Roxb.) Craib and Kadsura coccinea (Lem.) A.C. Sm. Foods 2020, 9, 1222. [Google Scholar] [CrossRef] [PubMed]
- Tasneem, S.; Yang, Y.; Liu, B.; Choudhary, M.I.; Wang, W. Cytotoxicity of Schisandronic Acid from Kadsura coccinea by Activation of Caspase-3, Cleavage of poly-ADP Ribose Polymerase, and Reduction of Oxidative Stress. Rev. Bras. Farmacogn. 2021, 31, 51–58. [Google Scholar] [CrossRef]
- Tram, L.H.; Thu Huong, T.; Thi Thuy, L.; Van Thong, N.; Tuan Anh, N.; Hoang Minh, N.; Thu Ha, T.; Anh Dung, D.; Thao, N.P.; Thuong, P.T.; et al. A new triterpenoid from the stems of Kadsura coccinea with antiproliferative activity. Nat. Prod. Res. 2021, 1–5. [Google Scholar] [CrossRef]
- Zuo, Y.H.; Liu, Y.B.; Cheng, C.S.; Yang, Y.P.; Xie, Y.; Luo, P.; Wang, W.; Zhou, H. Isovaleroylbinankadsurin A ameliorates cardiac ischemia/reperfusion injury through activating GR dependent RISK signaling. Pharmacol. Res. 2020, 158, 104897. [Google Scholar] [CrossRef] [PubMed]
- Amar, S.K.; Goyal, S.; Srivastav, A.K.; Chopra, D.; Ray, R.S. Combined effect of Benzophenone-2 and ultraviolet radiation promote photogenotoxicity and photocytotoxicity in human keratinocytes. Regul. Toxicol. Pharmacol. 2018, 95, 298–306. [Google Scholar] [CrossRef]
- Dubey, D.; Srivastav, A.K.; Singh, J.; Chopra, D.; Qureshi, S.; Kushwaha, H.N.; Singh, N.; Ray, R.S. Photoexcited triclosan induced DNA damage and oxidative stress via p38 MAP kinase signaling involving type I radicals under sunlight/UVB exposure. Ecotoxicol. Environ. Saf. 2019, 174, 270–282. [Google Scholar] [CrossRef]
- Dubey, D.; Chopra, D.; Singh, J.; Srivastav, A.K.; Kumari, S.; Verma, A.; Ray, R.S. Photosensitized methyl paraben induces apoptosis via caspase dependent pathway under ambient UVB exposure in human skin cells. Food Chem. Toxicol. 2017, 108, 171–185. [Google Scholar] [CrossRef]
- Park, J.U.; Yang, S.Y.; Guo, R.H.; Li, H.X.; Kim, Y.H.; Kim, Y.R. Anti-Melanogenic Effect of Dendropanax morbiferus and Its Active Components via Protein Kinase A/Cyclic Adenosine Monophosphate-Responsive Binding Protein- and p38 Mitogen-Activated Protein Kinase-Mediated Microphthalmia-Associated Transcription Factor Downregulation. Front. Pharmacol. 2020, 11, 507. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, J.S.; Kang, H.M.; Park, J.H.; Kang, J.S.; Lee, Y.J.; Park, Y.H.; Je, B.I.; Park, S.Y.; Choi, Y.W. A Comparative Study on Photo-Protective and Anti-Melanogenic Properties of Different Kadsura coccinea Extracts. Plants 2021, 10, 1633. https://doi.org/10.3390/plants10081633
Jeon JS, Kang HM, Park JH, Kang JS, Lee YJ, Park YH, Je BI, Park SY, Choi YW. A Comparative Study on Photo-Protective and Anti-Melanogenic Properties of Different Kadsura coccinea Extracts. Plants. 2021; 10(8):1633. https://doi.org/10.3390/plants10081633
Chicago/Turabian StyleJeon, Joong Suk, He Mi Kang, Ju Ha Park, Jum Soon Kang, Yong Jae Lee, Young Hoon Park, Byoung Il Je, Sun Young Park, and Young Whan Choi. 2021. "A Comparative Study on Photo-Protective and Anti-Melanogenic Properties of Different Kadsura coccinea Extracts" Plants 10, no. 8: 1633. https://doi.org/10.3390/plants10081633