
Reactive Oxygen Species in Host Plant Are Required for an Early Defense Response against Attack of Stagonospora nodorum Berk. Necrotrophic Effectors SnTox

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Necrotrophic Effector (NE) Sensitivity Genes Tsn1, Snn3-B1, and Snn1 in Three Cultivars of Bread Spring Wheat
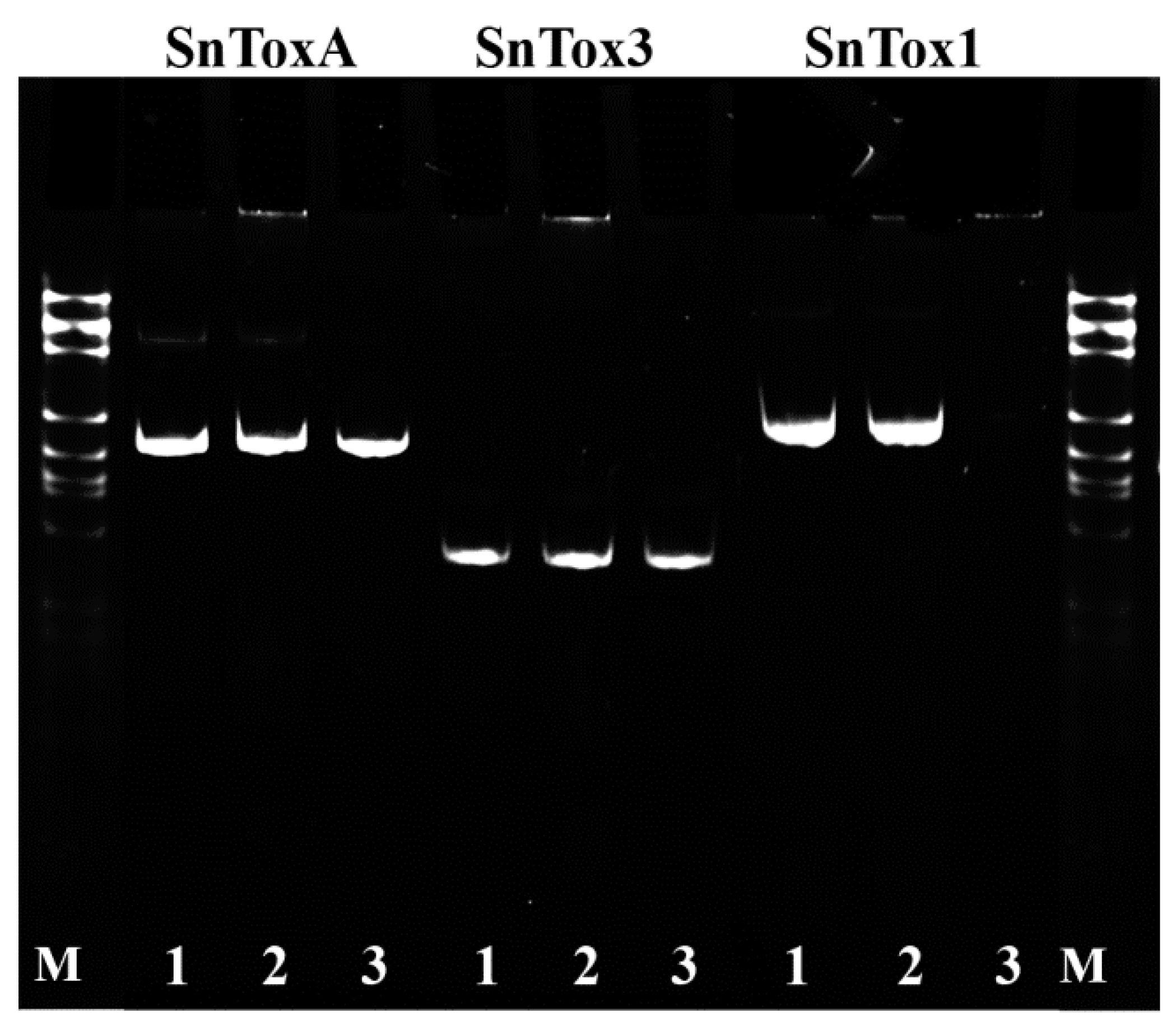
2.2. Analysis of Necrotrophic Effectors Genes of SnTox1, SnToxA, and SnTox3 in S. nodorum Isolates Sn4VD, SnB and Sn9MN
2.3. The Role of Compatible Interactions Tsn1–SnToxA, Snn3–SnTox3, Snn1–SnTox1 in Causing Disease
2.4. The Role of Compatible Interactions in Suppression of Reactive Oxygen Species (ROS) Production
3. Discussion
3.1. Expression of Necrotrophic Effectors Determines the Virulence of the S. nodorum Isolate
3.2. The S. nodorum NEs SnToxA, SnTox1 and SnTox3 Suppress the Early Defense Response of Plants Due to the Effect on the Enzymes of Redox Metabolism
4. Materials and Methods
4.1. Research Objects
4.2. Experimental Design
4.3. Seedling Resistance
4.4. Isolation of DNA and Performing the Polymerase Chain Reaction (PCR)
4.5. Gene Expression Analysis
4.6. Biochemical Parameters
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Jwa, N.-S.; Hwang, B.K. Convergent Evolution of Pathogen Effectors toward Reactive Oxygen Species Signaling Networks in Plants. Front. Plant Sci. 2017, 8, 1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, T.L.; Faris, J.D. Characterization of the wheat- Stagonospora nodorum disease system: What is the molecular basis of this quantitative necrotrophic disease interaction? Can. J. Plant Pathol. 2010, 32, 20–28. [Google Scholar] [CrossRef]
- Faris, J.D.; Zhang, Z.; Lu, H.; Lu, S.; Reddy, L.; Cloutier, S.; Fellers, J.P.; Meinhardt, S.W.; Rasmussen, J.B.; Xu, S.S.; et al. A unique wheat disease resistance-like gene governs effector-triggered susceptibility to necrotrophic pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 13544–13549. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Zhang, Z.; Friesen, T.L.; Raats, D.; Fahima, T.; Brueggeman, R.S.; Lu, S.; Trick, H.N.; Liu, Z.; Chao, W.; et al. The hijacking of a receptor kinase–driven pathway by a wheat fungal pathogen leads to disease. Sci. Adv. 2016, 2, e1600822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, H.T.; Rybak, K.; Furuki, E.; Breen, S.; Solomon, P.; Oliver, R.P.; Tan, K. Differential effector gene expression underpins epistasis in a plant fungal disease. Plant J. 2016, 87, 343–354. [Google Scholar] [CrossRef]
- Girardi, F.M.; Barra, M.B.; Zettler, C.G. Papillary thyroid carcinoma: Does the association with Hashimoto’s thyroiditis affect the clinicopathological characteristics of the disease? Braz. J. Otorhinolaryngol. 2015, 81, 283–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhang, Z.; Faris, J.; Oliver, R.P.; Syme, R.; McDonald, M.; McDonald, B.; Solomon, P.; Lu, S.; Shelver, W.L.; et al. The Cysteine Rich Necrotrophic Effector SnTox1 Produced by Stagonospora nodorum Triggers Susceptibility of Wheat Lines Harboring Snn1. PLoS Pathog. 2012, 8, e1002467. [Google Scholar] [CrossRef] [Green Version]
- Winterberg, B.; Du Fall, A.L.; Song, X.; Pascovici, D.; Care, N.; Molloy, M.; Ohms, S.; Solomon, P.S. The necrotrophic effector protein SnTox3 re-programs metabolism and elicits a strong defence response in susceptible wheat leaves. BMC Plant Biol. 2014, 14, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podgórska, A.; Burian, M.; Szal, B. Extra-Cellular but Extra-Ordinarily Important for Cells: Apoplastic Reactive Oxygen Species Metabolism. Front. Plant Sci. 2017, 8, 1353. [Google Scholar] [CrossRef] [Green Version]
- Barna, B.; Fodor, J.; Harrach, B.; Pogany, M.; Király, Z. The Janus face of reactive oxygen species in resistance and susceptibility of plants to necrotrophic and biotrophic pathogens. Plant Physiol. Biochem. 2012, 59, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Wang, J.; Gong, Z.; Zhou, J.M. Apoplastic ROS signaling in plant immunity. Curr. Opin. Plant Biol. 2017, 38, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, M.; Brown-Guedira, G.; Murphy, J.P.; Cowger, C. Genes Conferring Sensitivity to Stagonospora nodorum Necrotrophic Effectors in Stagonospora nodorum Blotch-Susceptible U.S. Wheat Cultivars. Plant Dis. 2014, 98, 746–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veselova, S.; Nuzhnaya, T.; Burkhanova, G.; Rumyantsev, S.; Khusnutdinova, E.; Maksimov, I. Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk. Biomolecules 2021, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Dmochowska-Boguta, M.; Nadolska-Orczyk, A.; Orczyk, W. Roles of peroxidases and NADPH oxidases in the oxidative response of wheat (Triticum aestivum) to brown rust (Puccinia triticina) infection. Plant Pathol. 2012, 62, 993–1002. [Google Scholar] [CrossRef]
- Veselova, S.V.; Burkhanova, G.F.; Nuzhnaya, T.V.; Maksimov, I.V. Roles of ethylene and cytokinins in development of defense responses in Triticum aestivum plants infected with Septoria nodorum. Russ. J. Plant Physiol. 2016, 63, 609–619. [Google Scholar] [CrossRef]
- Haugrud, A.R.P.; Zhang, Z.; Richards, J.; Friesen, T.L.; Faris, J.D. Genetics of Variable Disease Expression Conferred by Inverse Gene-For-Gene Interactions in the Wheat-Parastagonospora nodorum Pathosystem. Plant Physiol. 2019, 180, 420–434. [Google Scholar] [CrossRef]
- Abeysekara, N.S.; Friesen, T.L.; Keller, B.; Faris, J.D. Identification and characterization of a novel host–toxin interaction in the wheat–Stagonospora nodorum pathosystem. Theor. Appl. Genet. 2009, 120, 117–126. [Google Scholar] [CrossRef]
- Friesen, T.L.; Chu, C.; Xu, S.S.; Faris, J. SnTox5-Snn5: A novel Stagonospora nodorume ffector-wheat gene interaction and its relationship with the SnToxA-Tsn1and SnTox3-Snn3-B1interactions. Mol. Plant Pathol. 2012, 13, 1101–1109. [Google Scholar] [CrossRef]
- Gao, Y.; Faris, J.; Liu, Z.; Kim, Y.M.; Syme, R.; Oliver, R.P.; Xu, S.S.; Friesen, T.L. Identification and Characterization of the SnTox6-Snn6 Interaction in the Parastagonospora nodorum–Wheat Pathosystem. Mol. Plant Microbe Interact. 2015, 28, 615–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.; Friesen, T.L.; Saini, J.; Xu, S.S.; Rasmussen, J.B.; Faris, J.D. The Wheat Snn7 Gene Confers Susceptibility on Recognition of the Parastagonospora nodorum Necrotrophic Effector SnTox7. Plant Genome 2015, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.T.; Furuki, E.; Hunziker, L.; Rybak, K.; Tan, K.-C. GWAS analysis reveals distinct pathogenicity profiles of Australian Parastagonospora nodorum isolates and identification of marker-trait-associations to septoria nodorum blotch. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.K.; Kariyawasam, G.K.; Seneviratne, S.; Wyatt, N.A.; Xu, S.S.; Liu, Z.; Faris, J.D.; Friesen, T.L. A triple threat: The Parastagonospora nodorum SnTox267 effector exploits three distinct host genetic factors to cause disease in wheat. New Phytol. 2021. [Google Scholar] [CrossRef]
- Rybak, K.; See, P.T.; Phan, H.T.T.; Syme, R.A.; Moffat, C.S.; Oliver, R.P.; Tan, K.-C. A functionally conserved Zn2Cys6 binuclear cluster transcription factor class regulates necrotrophic effector gene expression and host-specific virulence of two major Pleosporales fungal pathogens of wheat. Mol. Plant Pathol. 2017, 18, 420–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faris, J.D.; Zhang, Z.; Rasmussen, J.B.; Friesen, T.L. Variable Expression of the Stagonospora nodorum Effector SnToxA among Isolates Is Correlated with Levels of Disease in Wheat. Mol. Plant Microbe Interact. 2011, 24, 1419–1426. [Google Scholar] [CrossRef] [Green Version]
- Virdi, S.K.; Liu, Z.; Overlander, M.E.; Zhang, Z.; Xu, S.S.; Friesen, T.L.; Faris, J.D. New Insights into the Roles of Host Gene-Necrotrophic Effector Interactions in Governing Susceptibility of Durum Wheat to Tan Spot and Septoria Nodorum Blotch. G3 (Bethesda) 2016, 6, 4139–4150. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.-C.; Oliver, R.P. Regulation of proteinaceous effector expression in phytopathogenic fungi. PLoS Pathog. 2017, 13, e1006241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.-G.; Faris, J.D.; Xu, S.S.; Friesen, T.L. Genetic analysis of disease susceptibility contributed by the compatible Tsn1–SnToxA and Snn1–SnTox1 interactions in the wheat-Stagonospora nodorum pathosystem. Theor. Appl. Genet. 2010, 120, 1451–1459. [Google Scholar] [CrossRef]
- Singh, R.; Dangol, S.; Chen, Y.; Choi, J.; Cho, Y.-S.; Lee, J.-E.; Choi, M.-O.; Jwa, A.N.-S. Magnaporthe oryzae Effector AVR-Pii Helps to Establish Compatibility by Inhibition of the Rice NADP-Malic Enzyme Resulting in Disruption of Oxidative Burst and Host Innate Immunity. Mol. Cells 2016, 39, 426–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkhanova, G.F.; Yarullina, L.G.; Maksimov, I. The control of wheat defense responses during infection with Bipolaris sorokiniana by chitooligosaccharides. Russ. J. Plant Physiol. 2007, 54, 104–110. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Winterberg, B.; Kobe, B.; Solomon, P.S. Wheat PR-1 proteins are targeted by necrotrophic pathogen effector proteins. Plant J. 2016, 88, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis Effector Pep1 Suppresses Plant Immunity by Inhibition of Host Peroxidase Activity. PLoS Pathog. 2012, 8, e1002684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Henderson, C.; Gurr, S.J. Blumeria graminis secretes an extracellular catalase during infection of barley: Potential role in suppression of host defence. Mol. Plant Pathol. 2004, 5, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Maksimov, I.V.; Yarullina, L.G.; Burkhanova, G.F.; Zaikina, E.A. Relationship between the aggressiveness and catalase activity of Septoria nodorum Berk. in wheat. Biol. Bull. 2013, 40, 441–446. [Google Scholar] [CrossRef]
- de Jonge, R.; van Esse, H.P.; Kombrink, A.; Shinya, T.; Desaki, Y.; Bours, R.; van der Krol, S.; Shibuya, N.; Joosten, M.H.A.J.; Thomma, B.P.H.J. Conserved Fungal LysM Effector Ecp6 Prevents Chitin-Triggered Immunity in Plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]
- Maksimov, I.V.; Cherepanova, E.; Khairullin, R.M. “Chitin-Specific” Peroxidases in Plants. Biochemistry 2003, 68, 111–115. [Google Scholar] [CrossRef]
- Veselova, S.V.; Nuzhnaya, T.V.; Maksimov, I. The effect of 1-methylcyclopropene on the components of pro- and antioxidant systems of wheat and the development of defense reactions in fungal pathogenesis. Appl. Biochem. Microbiol. 2014, 50, 516–523. [Google Scholar] [CrossRef]
- Liu, Z.H.; Friesen, T.L.; Rasmussen, J.B.; Ali, S.; Meinhardt, S.W.; Faris, J. Quantitative Trait Loci Analysis and Mapping of Seedling Resistance to Stagonospora nodorum Leaf Blotch in Wheat. Phytopathology 2004, 94, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Maniatis, T.; Fritsch, E.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 1982; p. 545. [Google Scholar]
- Fraaije, B.; Lovell, D.; Baldwin, S. Septoria epidemics on wheat: Combined use of visual assessment and PCR-based diagnostics to identify mechanisms of disease escape. Plant Prot. Sci. 2017, 38, 421–424. [Google Scholar] [CrossRef]
- Giovanini, M.P.; Puthoff, D.P.; Nemacheck, J.A.; Mittapalli, O.; Saltzmann, K.D.; Ohm, H.W.; Shukle, R.H.; Williams, C.E. Gene-for-Gene Defense of Wheat against the Hessian Fly Lacks a Classical Oxidative Burst. Mol. Plant Microbe Interact. 2006, 19, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Maksimov, I.V.; Valeev, A.S.; Cherepanova, E.A.; Burkhanova, G.F. Effect of chitooligosaccharides with different degrees of acetylation on the activity of wheat pathogen-inducible anionic peroxidase. Appl. Biochem. Microbiol. 2013, 50, 82–87. [Google Scholar] [CrossRef]
- Gimenez, M.J.; Pistón, F.; Atienza, S.G. Identification of suitable reference genes for normalization of qPCR data in comparative transcriptomics analyses in the Triticeae. Planta 2010, 233, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Bindschedler, L.; Minibayeva, F.; Gardner, S.L.; Gerrish, C.; Davies, D.R.; Bolwell, G.P. Early signalling events in the apoplastic oxidative burst in suspension cultured French bean cells involve cAMP and Ca2+. New Phytol. 2001, 151, 185–194. [Google Scholar] [CrossRef] [PubMed]





 —positive regulation,
—positive regulation,  —negative regulation, ???—unknown signal transduction pathway.
—positive regulation, —negative regulation, ???—unknown signal transduction pathway.
—negative regulation, ???—unknown signal transduction pathway.
—positive regulation, —negative regulation, ???—unknown signal transduction pathway.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate of S. nodorum | Necrotrophic Effectors | Cultivars | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Omskaya 35 (tsn1/snn3/Snn1) | Kazahstanskaya 10 (tsn1/Snn3/Snn1) | Zhnitsa (Tsn1/Snn3/Snn1) | ||||||||
| Time after Infection, Hours | ||||||||||
| 6 | 24 | 72 | 6 | 24 | 72 | 6 | 24 | 72 | ||
| Sn4VD (toxA/tox3/tox1) | SnToxA | 0.05 ± 0.02 a | 0.01 ± 0.01 a | 0.0 ± 0.0 a | 0.07 ± 0.05 a | 0.09 ± 0.4 a | 0.0 ± 0.0 a | 0.04 ± 0.02 a | 0.01 ± 0.12 a | 0.0 ± 0.0 a |
| SnTox3 | 0.01 ± 0.01 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.03 ± 0.02 a | 0.05 ± 0.04 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.07 ± 0.05 a | |
| SnTox1 | 0.0 ± 0.0 a | 0.04 ± 0.03 a | 0.03 ± 0.01 a | 0.04 ± 0.02 a | 0.04 ± 0.02 a | 0.01 ± 0.01 a | 0.04 ± 0.02 a | 0.01 ± 0.01 a | 0.01 ± 0.02 a | |
| SnB (ToxA/Tox3/tox1) | SnToxA | 0.01 ± 0.01 a | 0.4 ± 0.1 c | 0.17 ± 0.11 b | 0.0 ± 0.0 a | 0.9 ± 0.1 d | 1.2 ± 0.8 d | 1.72 ± 0.8 d | 1.4 ± 0.9 d | 2.8 ± 1.2 e |
| SnTox3 | 0.14 ± 0.01 b | 0.5 ± 0.1 c | 0.16 ± 0.1 b | 0.7 ± 0.01 c | 1.5 ± 0.3 d | 11.0 ± 3.6 j | 0.9 ± 0.2 d | 1.2 ± 0.4 d | 12.1 ± 3.5 k | |
| SnTox1 | 0.01 ± 0.01 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| Sn9MN (ToxA/Tox3/Tox1) | SnToxA | 0.01 ± 0.01 a | 0.08 ± 0.02 a | 0.27 ± 0.1 b | 0.0 ± 0.0 a | 0.1 ± 0.01 b | 1.1 ± 0.5 d | 3.5 ± 1.8 f | 2.4 ± 0.2 e | 5.12 ± 0.8 g |
| SnTox3 | 0.03 ± 0.01 a | 0.2 ± 0.03 b | 0.13 ± 0.04 b | 0.0 ± 0.0 a | 0.2 ± 0.02 b | 0.55 ± 0.1 c | 0.5 ± 0.2 c | 2.73 ± 0.1 e | 17.0 ± 1.6 l | |
| SnTox1 | 0.01 ± 0.01 a | 0.9 ± 0.4 d | 0.5 ± 0.1 c | 0.5 ± 0.1 c | 9.8 ± 2.4 i | 2.85 ± 1.5 e | 5.6 ± 1.3 g | 2.6 ± 0.3 e | 7.6 ± 2.2 h | |
| Cultivar | Damage Reaction | Isolate of S. nodorum | ||
|---|---|---|---|---|
| Sn4VD (toxa/tox3/tox1) | SnB (ToxA/Tox3/tox1) | Sn9MN (ToxA/Tox3/Tox1) | ||
| Omskaya 35 (tsn1/snn3/Snn1) | Necrosis, % | 0.05 ± 0.002 a | 5 ± 0.7 bc | 23 ± 2 e |
| Chlorosis, % | 0 ± 0 a | 3 ± 0.5 b | 0 ± 0 a | |
| Damage zone, % | 0.05 ± 0.001 a | 8 ± 1 c | 23 ± 2 e | |
| Damage score | 1 | 2 | 3 | |
| Resistance group * | RR | R | M | |
| Kazahstanskaya 10 (tsn1/Snn3/Snn1) | Necrosis, % | 0.05 ± 0.002 a | 16 ± 2 d | 31 ± 3 f |
| Chlorosis, % | 0 ± 0 a | 35 ± 3 f | 25 ± 2 e | |
| Damage zone, % | 0.05 ± 0.001 a | 51 ± 5 g | 56 ± 4 g | |
| Damage score | 1 | 4 | 4 | |
| Resistance group * | RR | S | S | |
| Zhnitsa (Tsn1/Snn3/Snn1) | Necrosis, % | 1 ± 0.1 b | 18 ± 2 d | 27 ± 2 e |
| Chlorosis, % | 2 ± 0.2 b | 55 ± 4 g | 57 ± 5 g | |
| Damage zone, % | 3 ± 0.3 b | 73 ± 6 h | 84 ± 6 i | |
| Damage score | 1 | 5 | 5 | |
| Resistance group * | RR | SS | SS | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veselova, S.; Nuzhnaya, T.; Burkhanova, G.; Rumyantsev, S.; Maksimov, I. Reactive Oxygen Species in Host Plant Are Required for an Early Defense Response against Attack of Stagonospora nodorum Berk. Necrotrophic Effectors SnTox. Plants 2021, 10, 1586. https://doi.org/10.3390/plants10081586
Veselova S, Nuzhnaya T, Burkhanova G, Rumyantsev S, Maksimov I. Reactive Oxygen Species in Host Plant Are Required for an Early Defense Response against Attack of Stagonospora nodorum Berk. Necrotrophic Effectors SnTox. Plants. 2021; 10(8):1586. https://doi.org/10.3390/plants10081586
Chicago/Turabian StyleVeselova, Svetlana, Tatyana Nuzhnaya, Guzel Burkhanova, Sergey Rumyantsev, and Igor Maksimov. 2021. "Reactive Oxygen Species in Host Plant Are Required for an Early Defense Response against Attack of Stagonospora nodorum Berk. Necrotrophic Effectors SnTox" Plants 10, no. 8: 1586. https://doi.org/10.3390/plants10081586