
Evolutionary Implications of a Peroxidase with High Affinity for Cinnamyl Alcohols from Physcomitrium patens, a Non-Vascular Plant


Abstract
:1. Introduction
2. Results
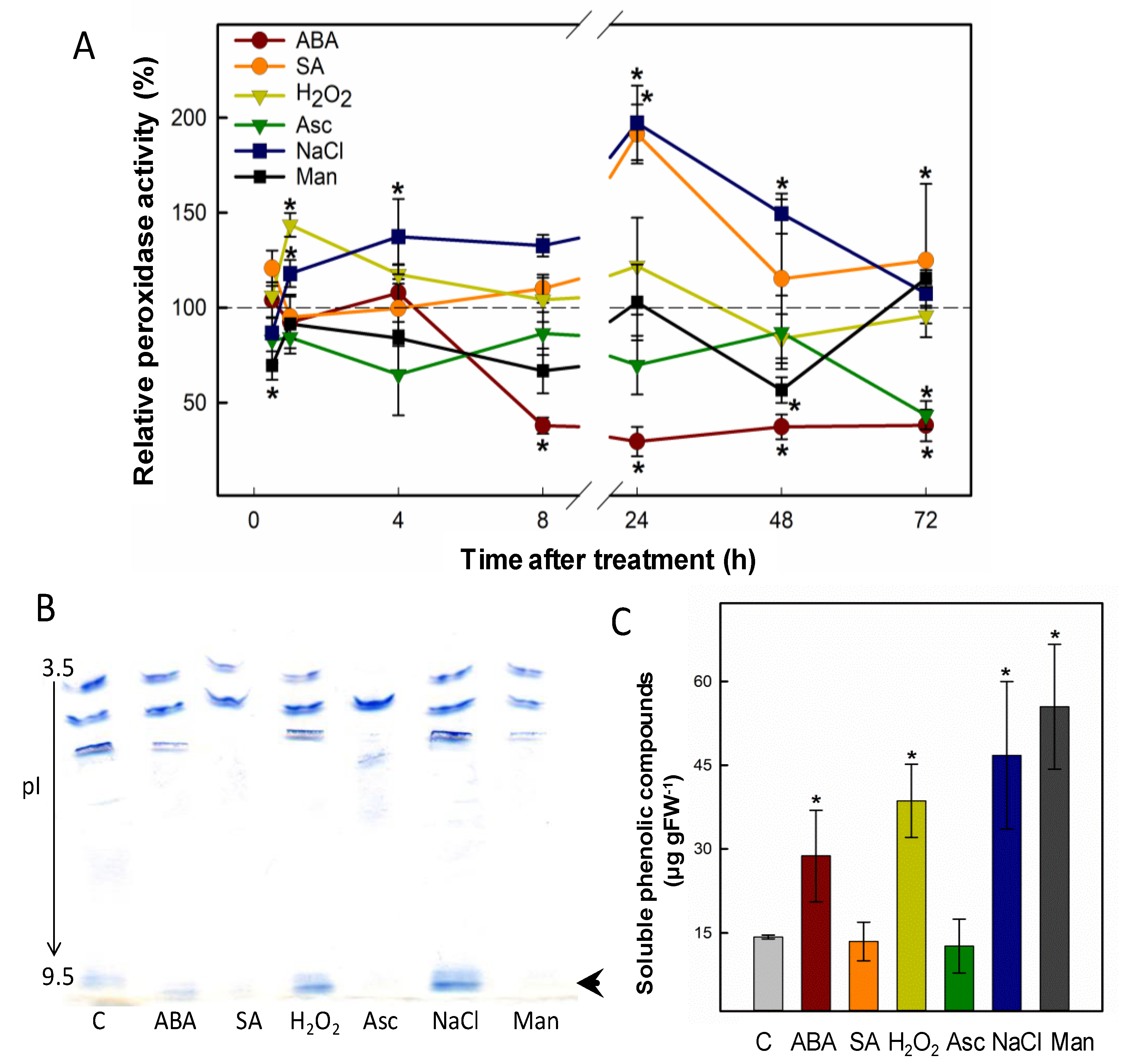
2.1. Abiotic Stress Strongly Modulates the Expression of a Basic Peroxidase in P. patens
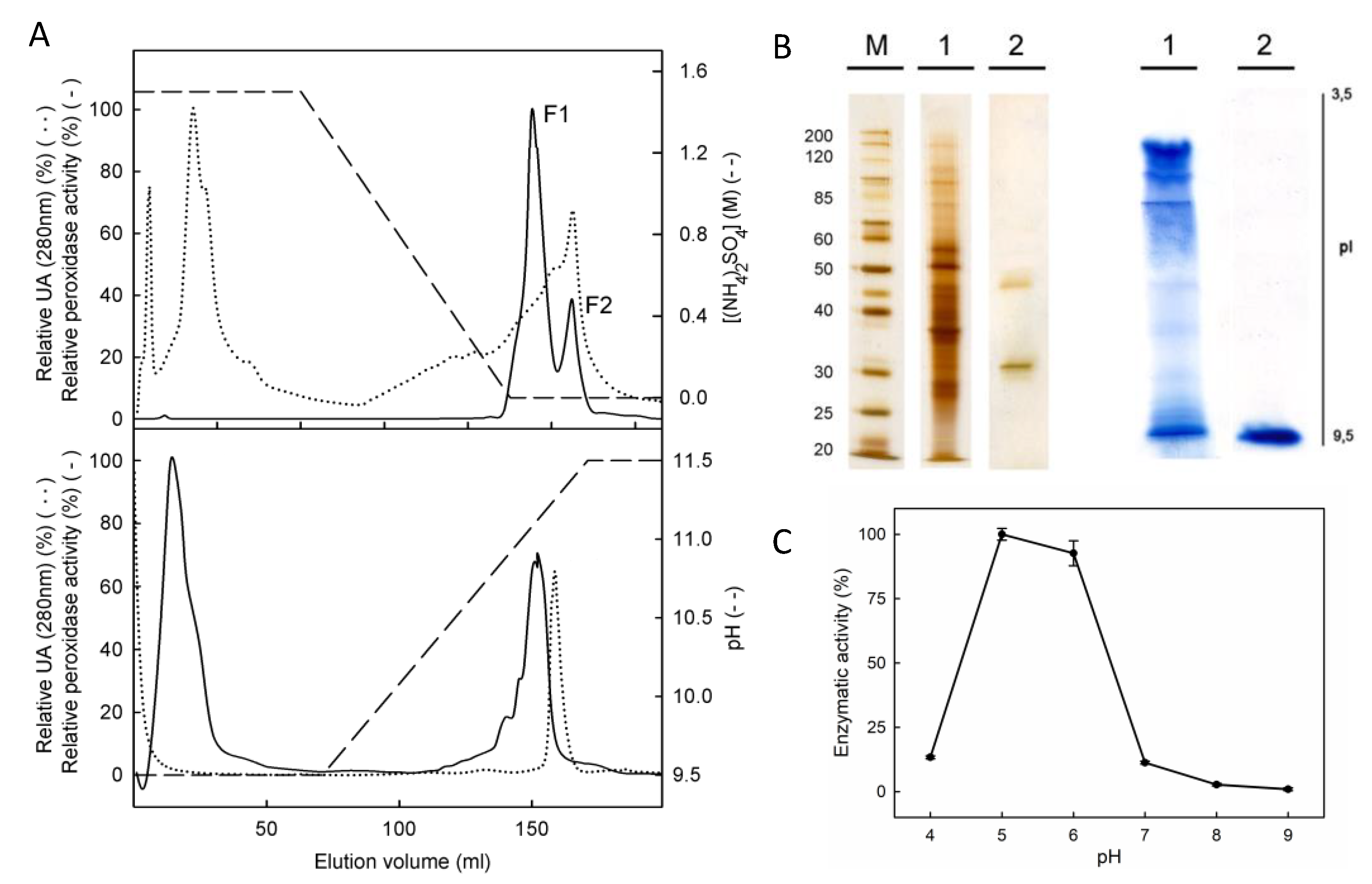
2.2. PpaPrx19 Is a 36 kDa Basic Peroxidase
2.3. PpaPrx19 Is Homologous to Peroxidases with a Role in Lignification
2.4. PpaPrx19 Shows High Affinity for Cinnamyl Alcohols
2.5. PpaPrx19 Associates with Lignin Biosynthesis Enzymes and Cell Wall-Related Proteins
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Protein Extraction and Precipitation
4.3. Purification of PpaPrx19
4.4. Peroxidase Activity Determination
4.5. Kinetic Data Analysis
4.6. Electrophoretic Analysis
4.7. Phenolic Compound Extraction and Quantification
4.8. Molecular Weight MALDI-TOF/TOF
4.9. Sequence Data Analysis
4.10. Gene Expression Analysis by RT-qPCR
4.11. Network Construction of PpaPrx19
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charron, A.J.; Quatrano, R.S. Between a rock and a dry place: The water-stressed moss. Mol. Plant 2009, 2, 478–486. [Google Scholar] [CrossRef] [Green Version]
- Kenrick, P.R.; Crane, P.R. The origin and early evolution of plants on land. Nature 1997, 389, 33–39. [Google Scholar] [CrossRef]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotechnol. 2019, 56, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, I.; Pettolino, F.A.; Bacic, A.; Ralph, J.; Lu, F.; O’Neill, M.A.; Fei, Z.; Rose, J.K.; Domozych, D.S.; Willats, W.G. The charophycean green algae provide insights into the early origins of plant cell walls. Plant J. 2011, 68, 201–211. [Google Scholar] [CrossRef]
- Espiñeira, J.M.; Novo-Uzal, E.; Gómez-Ros, L.V.; Carrión, J.S.; Merino, F.; Ros Barceló, A.; Pomar, F. Distribution of lignin monomers and the evolution of lignification among lower plants. Plant Biol. 2011, 13, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Renault, H.; Alber, A.; Horst, N.A.; Basilio Lopes, A.; Fich, E.A.; Kriegshauser, L.; Wiedemann, G.; Ullmann, P.; Herrgott, L.; Erhardt, M.; et al. A phenol-enriched cuticle is ancestral to lignin evolution in land plants. Nat. Commun. 2017, 8, 14713. [Google Scholar] [CrossRef]
- Weng, J.K.; Chapple, C. The origin and evolution of lignin biosynthesis. N. Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cézard, L.; Le Bris, P.; Borrega, N.; Hervé, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef] [Green Version]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. N. Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Welinder, K.G.; Justesen, A.F.; Kjaersgard, I.V.; Jensen, R.B.; Rasmussen, S.K.; Jespersen, H.M.; Duroux, L. Structural diversity and transcription of class III peroxidases from Arabidopsis thaliana. Eur. J. Biochem. 2002, 269, 6063–6081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosio, C.; Dunand, C. Specific functions of individual class III peroxidase genes. J. Exp. Bot. 2009, 60, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Goffinet, B.; Meyberg, R.; Wu, S.-Z.; Bezanilla, M. The Moss Physcomitrium (Physcomitrella) patens: A model organism for non-seed plants. Plant Cell 2020, 32, 1361–1376. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, M.T.; Akita, M.; Kalkkinen, N.; Ahola-Iivarinen, E.; Rönnholm, G.; Somervuo, P.; Thelander, M.; Valkonen, J.P.T. Quickly-released peroxidase of moss in defense against fungal invaders. N. Phytol. 2009, 183, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, M.T.; Akita, M.; Frank, W.; Reski, R.; Valkonen, J.P. Involvement of a class III peroxidase and the mitochondrial protein TSPO in oxidative burst upon treatment of moss plants with a fungal elicitor. Mol. Plant Microbe Interact. 2012, 25, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, W.; Ratnadewi, D.; Reski, R. Physcomitrella patens is highly tolerant against drought, salt and osmotic stress. Planta 2005, 220, 384–394. [Google Scholar] [CrossRef]
- Su, P.; Yan, J.; Li, W.; Wang, L.; Zhao, J.; Ma, X.; Li, A.; Wang, H.; Kong, L. A member of wheat class III peroxidase gene family, TaPRX-2A, enhanced the tolerance of salt stress. BMC Plant Biol. 2020, 20, 392. [Google Scholar] [CrossRef]
- Novo-Uzal, E.; Gutierrez, J.; Martinez-Cortes, T.; Pomar, F. Molecular cloning of two novel peroxidases and their response to salt stress and salicylic acid in the living fossil Ginkgo biloba. Ann. Bot. 2014, 114, 923–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mika, A.; Lüthje, S. Properties of guaiacol peroxidase activities isolated from corn root plasma membranes. Plant Physiol. 2003, 132, 1489–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergio, L.; Cardinali, A.; Di Paola, A.; Di Venere, D. Biochemical properties of soluble and bound peroxidases from artichoke heads and leaves. Food Technol. Biotechnol. 2009, 47, 32–38. [Google Scholar]
- Martinez-Cortés, T.; Pomar, F.; Espiñeira, J.M.; Merino, F.; Novo-Uzal, E. Purification and kinetic characterization of two peroxidases of Selaginella martensii Spring. involved in lignification. Plant Physiol. Biochem. 2012, 52, 130–139. [Google Scholar] [CrossRef]
- Adams, J.B. Regeneration and kinetics of peroxidase inactivation. Food Chem. 1997, 60, 201–206. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von Hijne, G.; Nielsen, H. SIGNALP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; Heijne, G.V. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Ligrone, R.; Ducket, J.G.; Renzaglia, K.S. Conducting tissues and phyletic relationships of bryophytes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 795–813. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Han, X.; Xu, X.; Shao, Y.; Zhu, Q.; Liu, Y.; Du, J.; Xu, J.; Zhang, S. WRKY15 suppresses tracheary element differentiation upstream of VND7 during xylem formation. Plant Cell 2020, 32, 2307–2324. [Google Scholar] [CrossRef] [Green Version]
- Herrero, J.; Esteban-Carrasco, A.; Zapata, J.M. Looking for Arabidopsis thaliana peroxidases involved in lignin biosynthesis. Plant Physiol. Biochem. 2013, 67, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, L.; Teilum, K.; Mirza, O.; Mattsson, O.; Petersen, M.; Welinder, K.G.; Mundy, J.; Gajhede, M.; Henriksen, A. Arabidopsis ATP A2 peroxidase. Expression and high-resolution structure of a plant peroxidase with implications for lignification. Plant Mol. Biol. 2000, 44, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Kakrana, A.; Kumar, A.; Satheesh, V.; Abdin, M.Z.; Subramaniam, K.; Bhattacharya, R.C.; Srinivasan, R.; Sirohi, A.; Jain, P.K. Identification, validation and utilization of novel nematode-responsive root-specific promoters in Arabidopsis for inducing host-delivered RNAi mediated root-knot nematode resistance. Front. Plant Sci. 2017, 8, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.H.; Kim, S.Y.; Nam, K.H. Genes encoding plant-specific class III peroxidases are responsible for increased cold tolerance of the brassinosteroid-insensitive 1 mutant. Mol. Cells 2012, 34, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, F.; Pomar, F.; Pedreño, M.A.; Novo-Uzal, E. The suppression of AtPrx52 affects fibers but not xylem lignification in Arabidopsis by altering the proportion of syringyl units. Physiol. Plant 2015, 154, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Irshad, M.; Canut, H.; Borderies, G.; Pont-Lezica, R.; Jamet, E. A new picture of cell wall protein dynamics in elongating cells of Arabidopsis thaliana: Confirmed actors and newcomers. BMC Plant Biol. 2008, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Bindschedler, L.V.; Whitelegge, J.P.; Millar, D.J.; Bolwell, G.P. A two component chitin-binding protein from French bean: Association of a proline-rich protein with a cysteine-rich polypeptide. FEBS Lett. 2006, 580, 1541–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gara, L. Class III peroxidases and ascorbate metabolism in plants. Phytochem. Rev. 2004, 3, 195–205. [Google Scholar] [CrossRef]
- Liszkay, A.; van der Zalm, E.; Schopfer, P. Production of reactive oxygen intermediates (O2-), H2O2, and OH) by maize roots and their role in wall loosening and elongation growth. Plant Physiol. 2004, 36, 3114–3123. [Google Scholar] [CrossRef] [Green Version]
- Gazaryan, I.G.; Lagrimini, L.M.; Ashby, G.A.; Thorneley, N.F. Mechanism of indole-3-acetic acid oxidation by plant peroxidases: Anaerobic stopped-flow spectrophotometric studies on horseradish and tobacco peroxidases. Biochem. J. 1996, 313, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A large family of class III plant peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, R.D.; Rancour, D.M.; Marita, J.M. Grass cell walls: Story of cross-linking. Front. Plant Sci. 2017, 7, 2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerstedt, K.V.; Kukkola, E.M.; Koistinen, V.V.; Takahashi, J.; Marjamaa, K. Cell wall lignin is polymerised by class III secretable plant peroxidases in Norway spruce. J. Integr. Plant Biol. 2010, 52, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Ros Barceló, A.; Gómez Ros, L.V.; Esteban-Carrasco, A. Looking for syringyl peroxidases. Trends Plant Sci. 2007, 12, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Kim, K.W.; Cho, M.H.; Franceschi, V.R.; Davin, L.D.; Lewis, N.G. Expression of cinnamyl alcohol dehydrogenases and their putative homologues during Arabidopsis thaliana growth and development: Lessons for database annotations? Phytochemistry 2007, 68, 1957–1974. [Google Scholar] [CrossRef] [PubMed]
- Moinuddin, S.G.; Jourdes, M.; Laskar, D.D.; Ki, C.; Cardenas, C.L.; Kim, K.W.; Zhang, D.; Davin, L.B.; Lewis, N.G. Insights into lignin primary structure and deconstruction from Arabidopsis thaliana COMT (caffeic acid O-methyl transferase) mutant Atomt1. Org. Biomol. Chem. 2010, 8, 3928–3946. [Google Scholar] [CrossRef] [PubMed]
- Møller, S.; Yi, X.; Velásquez, S.M.; Gille, S.; Hansen, P.L.M.; Poulsen, C.P.; Olsen, C.E.; Rejzek, M.; Parsons, H.; Yang, Z.; et al. Identification and evolution of a plant cell wall specific glycoprotein glycosyl transferase, ExAD. Sci. Rep. 2017, 7, 45341. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Takada, R.; Tobimatsu, Y.; Suzuki, S.; Yamamura, M.; Osakabe, K.; Osakabe, Y.; Sakamoto, M.; Umezawa, T. Double knockout of OsWRKY36 and OsWRKY102 boosts lignification with altering culm morphology of rice. Plant Sci. 2020, 296, 110466. [Google Scholar] [CrossRef]
- Martone, P.T.; Estevez, J.M.; Lu, F.; Ruel, K.; Denny, M.W.; Somerville, C.; Ralph, J. Discovery of lignin in seaweed reveals convergent evolution of cell-wall architecture. Curr. Biol. 2009, 19, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutaniemi, S.; Toikka, M.M.; Kärkönen, A.; Mustonen, M.; Lundell, T.; Simola, L.K.; Kilpeläinen, I.A.; Teeri, T.H. Characterization of basic p-coumaryl and coniferyl alcohol peroxidases from a lignin-forming Picea abies suspension culture. Plant Mol. Biol. 2005, 58, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, M.; Guerrero, C.; Botella, M.A.; Barcelo, A.; Amaya, I.; Medina, M.I.; Alonso, F.J.; de Forchetti, S.M.; Tigier, H.; Valpuesta, V. A tomato peroxidase involved in the synthesis of lignin and suberin. Plant Physiol. 2000, 122, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Gabaldón, C.; López-Serrano, M.; Pedreño, M.A.; Ros Barceló, A. Cloning and molecular characterization of the basic peroxidase isoenzyme from Zinnia elegans, an enzyme involved in lignin biosynthesis. Plant Physiol. 2005, 139, 1138–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moural, T.W.; Lewis, K.M.; Barnaba, C.; Zhu, F.; Palmer, N.A.; Sarath, G.; Scully, E.D.; Jones, J.P.; Sattler, S.E.; Kang, C. Characterization of class III peroxidases from switchgrass. Plant Physiol. 2017, 173, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, F.C.; Davin, L.B.; Lewis, N.G. The Arabidopsis phenylalanine ammonia lyase gene family: Kinetic characterization of the four PAL isoforms. Phytochemistry 2004, 65, 1557–1564. [Google Scholar] [CrossRef]
- Baltas, M.; Lapeyre, C.; Bedos-Belval, F.; Maturano, M.; Saint-Aguet, P.; Roussel, L.; Duran, H.; Grima-Pettenati, J. Kinetic and inhibition studies of cinnamoyl-CoA reductase 1 from Arabidopsis thaliana. Plant Physiol. Biochem. 2005, 43, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Naik, P.P.; Chen, H.C.; Shi, R.; Lin, C.Y.; Liu, J.; Shuford, C.M.; Li, Q.; Sun, Y.H.; Tunlaya-Anukit, S.; et al. Complete proteomic-based enzyme reaction and inhibition kinetics reveal how monolignol biosynthetic enzyme families affect metabolic flux and lignin in Populus trichocarpa. Plant Cell 2014, 26, 894–914. [Google Scholar] [CrossRef] [Green Version]
- Solé-Gil, A.; Hernández-García, J.; López-Gresa, M.P.; Blázquez, M.A.; Agustí, J. Conservation of thermospermine synthase activity in vascular and non-vascular plants. Front. Plant Sci. 2019, 10, 663. [Google Scholar] [CrossRef]
- Moghe, G.D.; Last, R.L. Something old, something new: Conserved enzymes and the evolution of novelty in plant specialized metabolism. Plant Physiol. 2015, 169, 1512–1523. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhang, D.; Hu, J.; Zhou, X.; Ye, X.; Reichel, K.L.; Stewart, N.R.; Syrenne, R.D.; Yang, X.; Gao, P.; et al. Comparative genome analysis of lignin biosynthesis gene families across the plant kingdom. BMC Bioinform. 2009, 10 (Suppl. 11), S3. [Google Scholar] [CrossRef] [Green Version]
- Labeeuw, L.; Martone, P.T.; Boucher, Y.; Case, R.J. Ancient origin of the biosynthesis of lignin precursors. Biol. Direct. 2015, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, H.; Werck-Reichhart, D.; Weng, J.K. Harnessing lignin evolution for biotechnological applications. Curr. Opin. Biotechnol. 2019, 56, 105–111. [Google Scholar] [CrossRef]
- Benito, B.; Rodriguez Navarro, A. Molecular cloning and characterization of a sodium-pump ATPase of the moss Physcomitrella patens. Plant J. 2003, 36, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, M.; Dreyer, M.K.; Wendt, K.U. APRV—A program for automated data processing, refinement and visualization. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 1679–1682. [Google Scholar] [CrossRef]
- Minami, A.; Nagao, M.; Arakawa, K.; Fujikawa, S.; Takezawa, D. Abscisic acid-induced freezing tolerance in the moss Physcomitrella patens is accompanied by increased expression of stress-related genes. J. Plant Physiol. 2003, 160, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Richardt, S.; Timmerhaus, G.; Lang, D.; Qudeimat, E.; Corrêa, L.G.G.; Reski, R.; Rensing, S.A.; Frank, W. Microarray analysis of the moss Physcomitrella patens reveals evolutionarily conserved transcriptional regulation of salt stress and abscisic acid signaling. Plant Mol. Biol. 2010, 72, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhong, N.Q.; Wang, Q.L.; Li, L.J.; Liu, X.L.; He, Y.K.; Xia, G.X. Cloning and functional characterization of PpDBF1 gene encoding a DRE-binding transcription factor from Physcomitrella patens. Planta 2007, 226, 827–838. [Google Scholar] [CrossRef]
- de Marco, A.; Guzzardi, P.; Jamet, E. Isolation of tobacco isoperoxidases accumulated in cell-suspension culture medium and characterization of activities related to cell wall metabolism. Plant Physiol. 1999, 120, 371–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. BioTechniques 1993, 15, 532–534. [Google Scholar] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Peroxidase Activity (nkat) | Specific Activity (nkat mg−1 Protein) | Purification Fold | Yield (%) | |
|---|---|---|---|---|
| 95% (NH4)2SO4 precipitation | 698 | 28 | 1 | 100 |
| Phenyl Sepharose chromatography | 483 | 266 | 10 | 69 |
| SP Sepharose chromatography | 218 | 23,644 | 850 | 31 |
| Prx Name | Species | Taxonomical Group | Identity (%) | E-Value |
|---|---|---|---|---|
| PpaPrx18 | Physcomitrium patens | Bryophyte | 66 | 3 × 10−144 |
| TruPrx01 | Tortula ruralis (star moss) | Bryophyte | 65 | 3 × 10−133 |
| PpaPrx09 | Physcomitrium patens | Bryophyte | 61 | 1 × 10−124 |
| CppPrx02 | Citrus x Paradisi x Poncirus | Angiosperm | 49 | 3 × 10−97 |
| PtaPrx102 | Pinus taeda (loblolly pine) | Gymnosperm | 50 | 8 × 10−97 |
| CsPrx62 | Citrus sinensis | Angiosperm | 49 | 2 × 10−96 |
| PabPrx05 | Picea abies (Norway spruce) | Gymnosperm | 49 | 6 × 10−95 |
| TsPrx15 | Thellungiella salsuginea | Angiosperm | 47 | 1 × 10−94 |
| BrPrx15-1 | Brassica rapa | Angiosperm | 48 | 6 × 10−94 |
| PtaPrx28 | Pinus taeda (loblolly pine) | Gymnosperm | 46 | 9 × 10−94 |
| PabPrx125 | Picea abies (Norway spruce) | Gymnosperm | 48 | 4 × 10−93 |
| GbPrx04 | Ginkgo biloba | Gymnosperm | 49 | 6 × 10−93 |
| Peroxidase Name | TAIR Gene ID | Identity (%) | E-Value | Function | Reference |
|---|---|---|---|---|---|
| AtPrx15 | At2g18150 | 47 | 4 × 10−93 | Lignification/abiotic stress | [25] |
| AtPrx49 | At4g36430 | 47 | 6 × 10−93 | Lignification | [26] |
| AtPrx53 | At5g06720 | 46 | 1 × 10−92 | Lignification | [27] |
| AtPrx14 | At2g18140 | 48 | 2 × 10−92 | Biotic stress | [28] |
| AtPrx22 | At2g38380 | 47 | 5 × 10−91 | Cold tolerance | [29] |
| AtPrx52 | At5g05340 | 48 | 1 × 10−89 | Lignification | [30] |
| AtPrx32 | At3g32980 | 45 | 5 × 10−88 | Cell elongation | [31] |
| AtPrx34 | At3g49120 | 45 | 7 × 10−88 | Oxidative burst | [32] |
| Substrate | Peroxidase Activity |
|---|---|
| Ascorbic acid | n.d. |
| NADH | n.d. |
| Indole-3-acetic acid (IAA) | 0.01 ± 0.00 |
| Ferulic acid | 0.53 ± 0.07 |
| Coniferyl alcohol | 1.46 ± 0.13 |
| Sinapyl alcohol | 0.08 ± 0.01 |
| Substrate | KM (μM) | Kcat (s−1) | Kcat/KM (μM−1 s−1) |
|---|---|---|---|
| CA | 16.7 | 3940.0 | 235.8 |
| SA | 20.8 | 281.0 | 13.5 |
| P. patens Protein ID | A. thaliana Homolog | Function | Score | |
|---|---|---|---|---|
| PP1S84_209V6.1 | CADG | AT1G72680.1 | Cinnamyl alcohol dehydrogenase | 0.650 |
| PP1S126_185V6.1 | CAD9 | AT4G39330.1 | Cinnamyl alcohol dehydrogenase | 0.650 |
| PP1S163_63V6.1 | ELI3-2 | AT4G37990.1 | Cinnamyl alcohol dehydrogenase | 0.650 |
| PP1S56_71V6.1 | OMT1 | AT5G54160.1 | O-methyltransferase | 0.650 |
| PP1S123_38V6.1 | AT1G78780 | PR protein | 0.546 | |
| PP1S34_74V6.1 | AT1G29850 | DNA binding | 0.546 | |
| PP1S141_102V6.3 | MBF1A | AT2G42680.1 | Multiprotein bridging factor | 0.531 |
| CHI | AT2G43590 | Chitinase | 0.521 | |
| PP1S96_94V6.1 | MBF1B | AT3G58680.1 | Multiprotein bridging factor | 0.400 |
| PP1S35_215V6.1 | MBF1B | AT3G58680.1 | Multiprotein bridging factor | 0.400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Cortés, T.; Pomar, F.; Novo-Uzal, E. Evolutionary Implications of a Peroxidase with High Affinity for Cinnamyl Alcohols from Physcomitrium patens, a Non-Vascular Plant. Plants 2021, 10, 1476. https://doi.org/10.3390/plants10071476
Martínez-Cortés T, Pomar F, Novo-Uzal E. Evolutionary Implications of a Peroxidase with High Affinity for Cinnamyl Alcohols from Physcomitrium patens, a Non-Vascular Plant. Plants. 2021; 10(7):1476. https://doi.org/10.3390/plants10071476
Chicago/Turabian StyleMartínez-Cortés, Teresa, Federico Pomar, and Esther Novo-Uzal. 2021. "Evolutionary Implications of a Peroxidase with High Affinity for Cinnamyl Alcohols from Physcomitrium patens, a Non-Vascular Plant" Plants 10, no. 7: 1476. https://doi.org/10.3390/plants10071476