
Effects of the Simulated Enhancement of Precipitation on the Phenology of Nitraria tangutorum under Extremely Dry and Wet Years

 ,
,  and
and
Abstract
:1. Introduction
2. Results
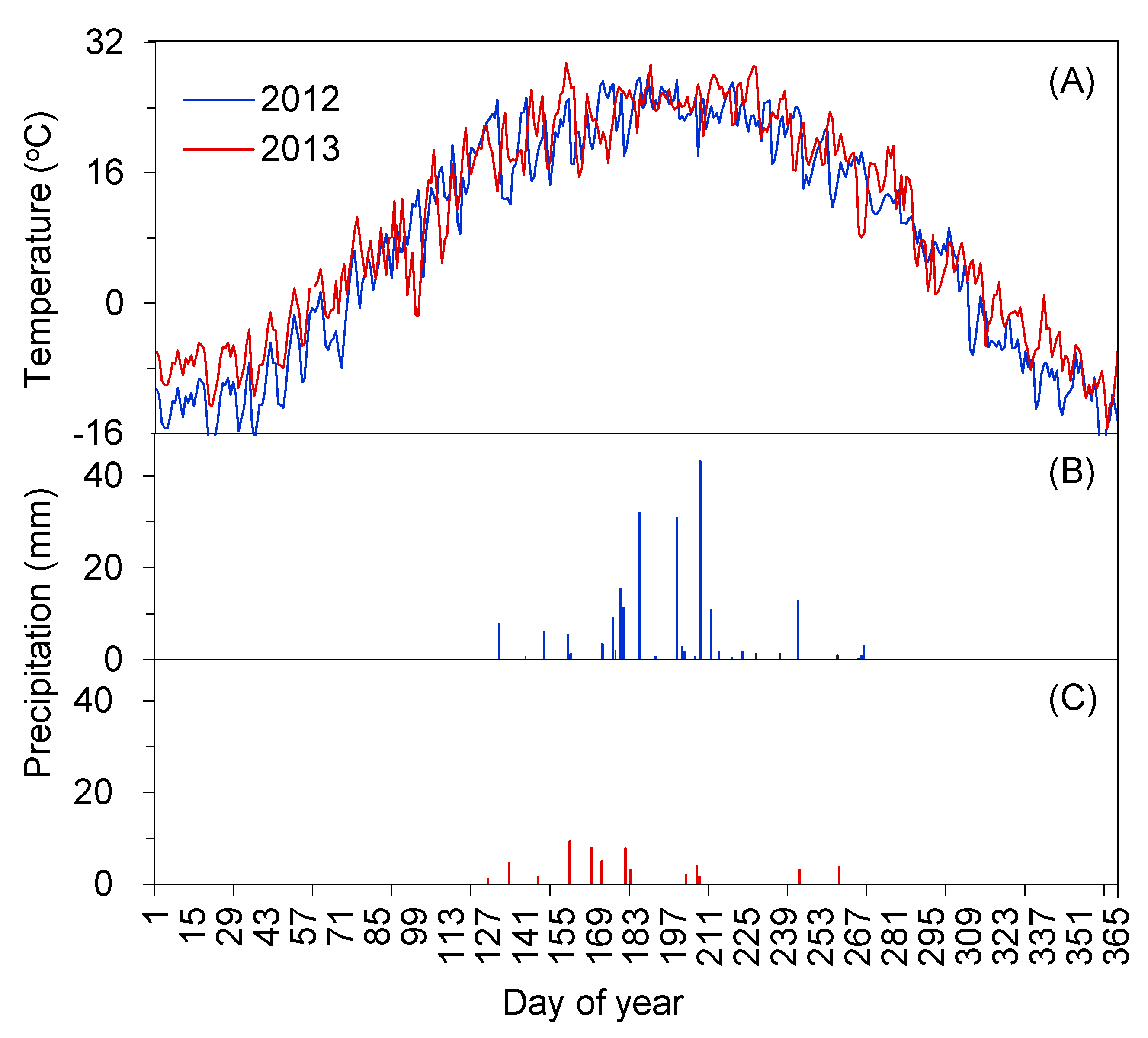
2.1. Interannual Dynamics of Meteorological Factors
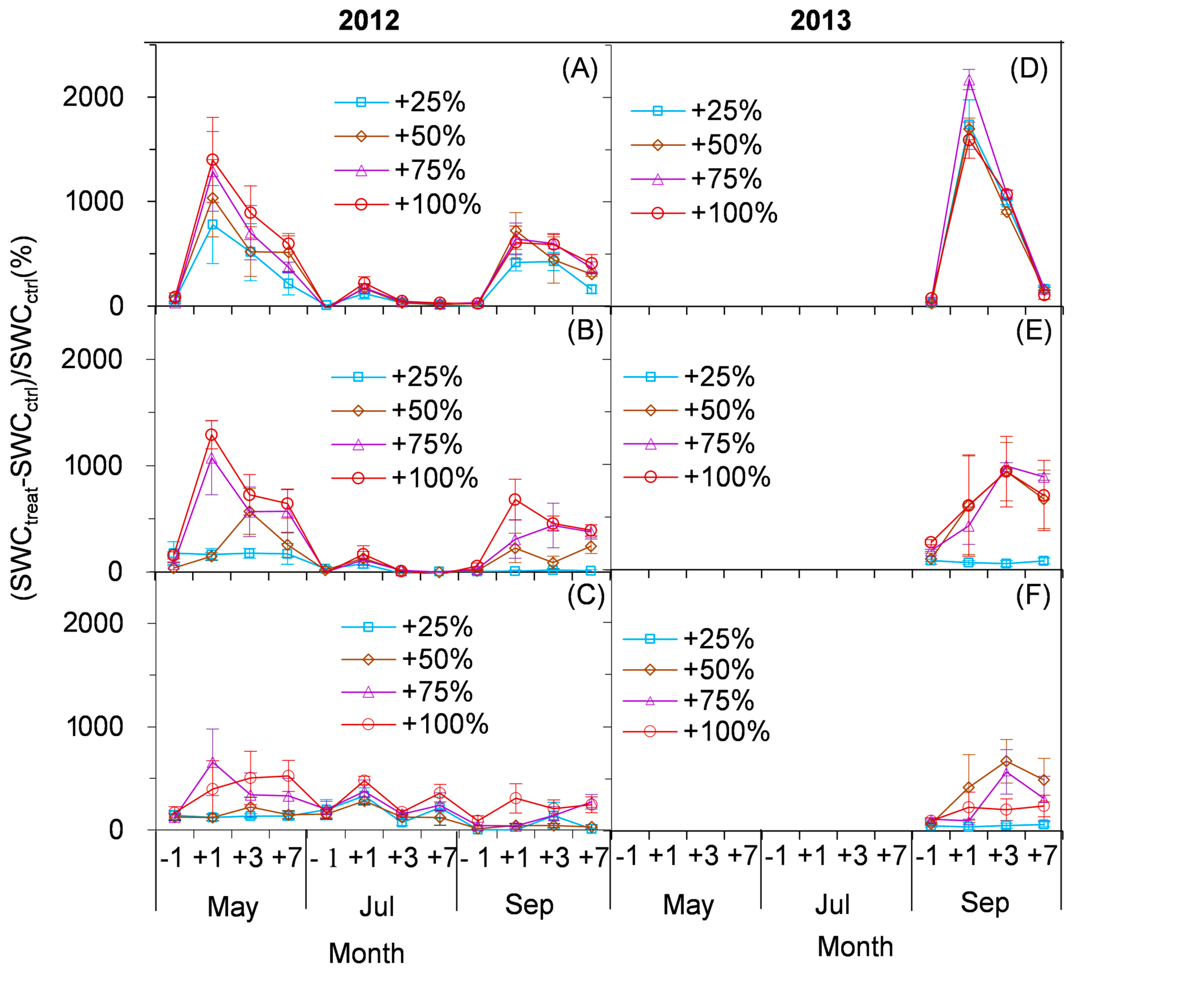
2.2. Effect of Simulated Enhancement of Precipitation on the Content of Soil Water
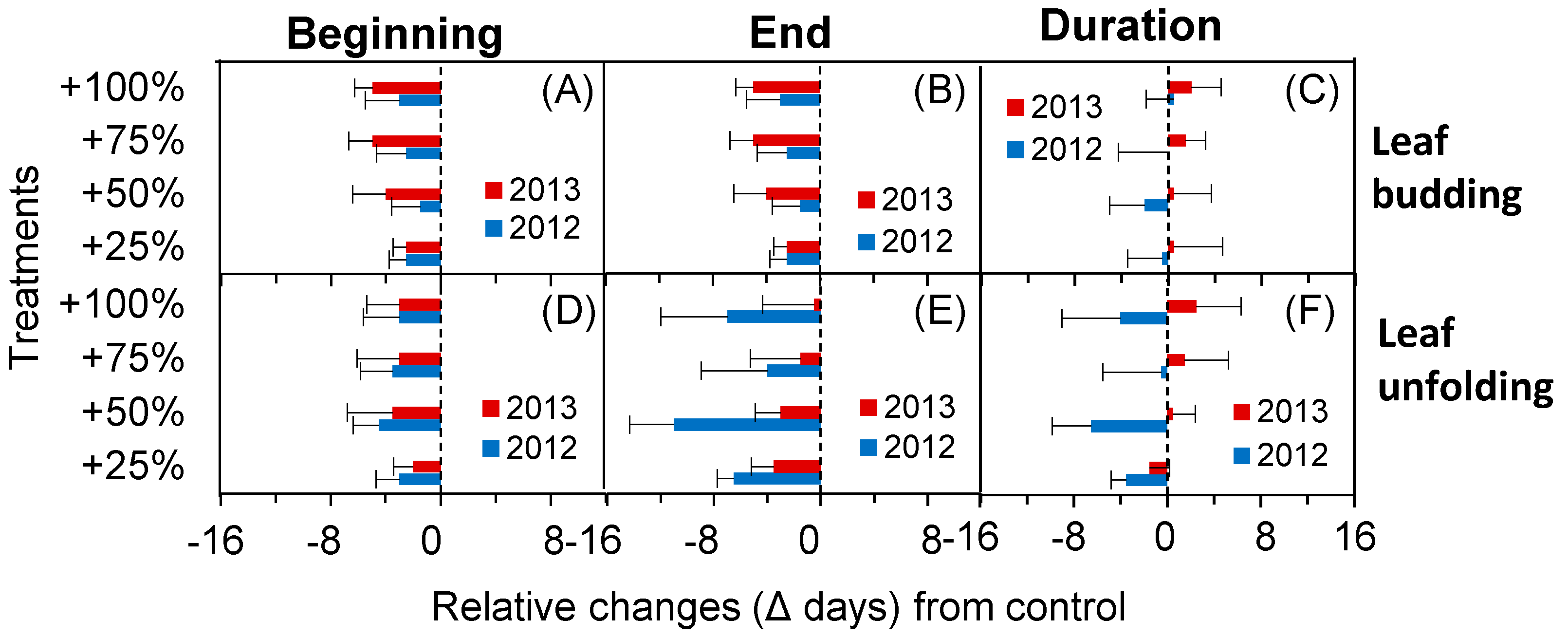
2.3. Phenological Fluctuation during the Spring in Extremely Dry and Wet Years
2.4. Phenological Fluctuation during the Autumn in Extremely Dry and Wet Years
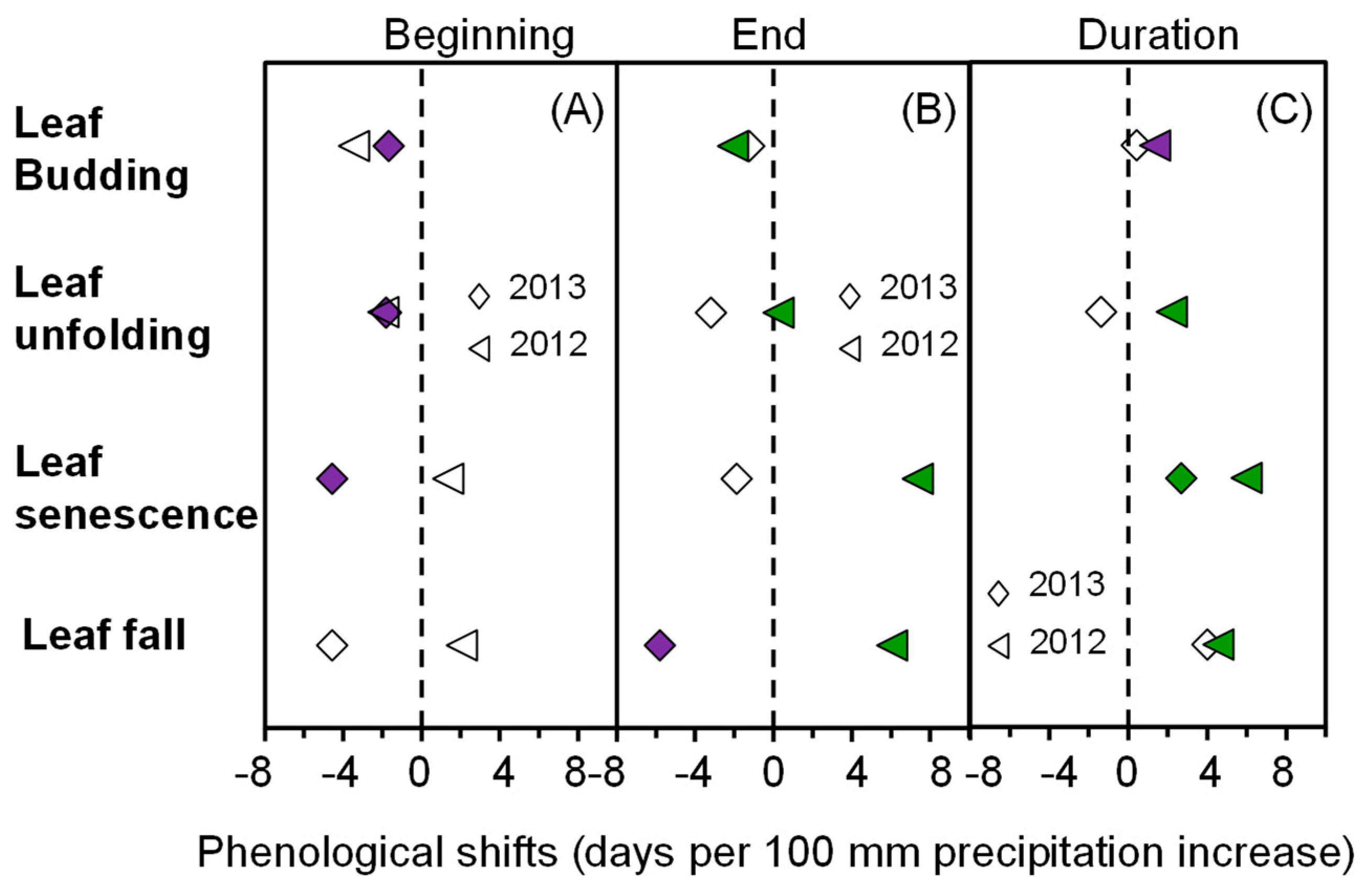
2.5. The Correlation between Phenology and the Simulated Increase in Precipitation
3. Discussion
3.1. Effect of the Simulated Enhancement of Precipitation on Phenology in the Spring
3.2. Effects of the Simulated Enhancement of Precipitation on Autumn Phenology
4. Materials and Methods
4.1. Sample Area Description
4.2. Simulated Enhancement in Precipitation
4.3. Phenological Observations
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Badeck, F.W.; Bondeau, A.; Bottcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Stephen, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Bertin, R.I. Plant phenology and distribution in relation to recent climate change. J. Torrey Bot. Soc. 2008, 135, 126–146. [Google Scholar] [CrossRef] [Green Version]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Chang. Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef]
- Nord, E.A.; Lynch, J.P. Plant phenology: A critical controller of soil resource acquisition. J. Exp. Bot. 2009, 60, 1927–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Menzel, A.; Estrella, N.; Fabian, P. Spatial and temporal variability of the phenological seasons in Germany from 1951 to 1996. Glob. Chang. Biol. 2001, 7, 657–666. [Google Scholar]
- Fu, Y.S.H.; Piao, S.L.; Op de Beeck, M.; Cong, N.; Zhao, H.F.; Zhang, Y.; Menzel, A.; Janssens, I.A. Recent spring phenology shifts in western Central Europe based on multiscale observations. Glob. Ecol. Biogeogr. 2014, 23, 1255–1263. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Reiter, B.E. Changes in North American spring. Int. J. Climatol. 2000, 20, 929–932. [Google Scholar] [CrossRef]
- Jeong, S.J.; Ho, C.H.; Gim, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Fridley, J.D. Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 2012, 485, 359–362. [Google Scholar] [CrossRef]
- Ge, Q.S.; Wang, H.J.; Zheng, J.Y.; This, R.; Dai, J.H. A 170 year spring phenology index of plants in eastern China. J. Geophys. Res. Biogeosci. 2014, 119, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.S.; Wang, H.J.; Rutishauser, T.; Dai, J.H. Phenological response to climate change in China: A meta-analysis. Glob. Chang. Biol. 2015, 21, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhu, W.; Chen, G.; Jiang, N.; Fan, D.; Zhang, D. Continuous but diverse advancement of spring-summer phenology in response to climate warming across the Qinghai-Tibetan Plateau. Agric. For. Meterol. 2016, 223, 194–202. [Google Scholar] [CrossRef]
- Chambers, L.E.; Altwegg, R.; Barbraud, C.; Barnard, P.; Beaumont, L.J.; Crawford, R.J.; Durant, J.M.; Hughes, L.; Keatley, M.R.; Low, M.; et al. Phenological changes in the southern hemisphere. PLoS ONE 2013, 8, e75514. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.L.; Huete, A.; Yu, Q.; Coupe, N.R.; Davies, K.; Broich, M.; Ratana, P.; Beringer, J.; Hutley, L.B.; Cleverly, J.; et al. Spatial patterns and temporal dynamics in savanna vegetation phenology across the North Australian Tropical Transect. Remote Sens. Environ. 2013, 139, 97–115. [Google Scholar] [CrossRef]
- Moore, L.M.; Lauenroth, W.K.; Bell, D.M.; Schlaepfer, D.R. Soil Water and Temperature Explain Canopy Phenology and Onset of Spring in a Semiarid Steppe. Great Plains Res. 2015, 25, 121–138. [Google Scholar] [CrossRef]
- Bao, F.; Liu, M.; Cao, Y.; Li, J.; Yao, B.; Xin, Z.; Lu, Q.; Wu, B. Water addition prolonged the length of the growing season of the desert shrub Nitraria tangutorum in a temperate desert. Front. Plant Sci. 2020, 11, 1099. [Google Scholar] [CrossRef]
- Ghazahfar, S.A. The phenology of desert plants: A 3-year study in a gravel desert wadi in northern Oman. J. Arid Environ. 1997, 35, 407–417. [Google Scholar] [CrossRef]
- Gebauer, R.L.E.; Ehleringer, J.R. Water and nitrogen uptake patterns following moisture pulses in a cold desert community. Ecology 2000, 81, 1415–1424. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K. Modifying the ‘pulse–reserve’ paradigm for deserts of North America: Precipitation pulses, soil water, and plant responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef]
- Patrick, L.D.; Ogle, K.; Bell, C.W.; Zak, J.; Tissue, D. Physiological responses of two contrasting desert plant species to precipitation variability are differentially regulated by soil moisture and nitrogen dynamics. Glob. Chang. Biol. 2009, 15, 1214–1229. [Google Scholar] [CrossRef]
- Sakkir, S.; Shah, J.N.; Cheruth, A.J.; Kabshawi, M. Phenology of desert plants from an arid gravel plain in eastern United Arab Emirates. J. Arid Land 2014, 7, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Zhang, X.; Yu, Y.; Guo, W.; Hanan, N.P. Characterizing land surface phenology and responses to precipitation in the Sahara Desert. J. Geophys. Res. Biogeosci. 2016, 121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Cao, J.; Bai, Y.; Zhou, X.; Ning, Z.; Yang, S.; Hu, L. Effects of precipitation amount and frequency on vegetation growth in a Tibetan alpine meadow. Clim. Chang. 2013, 118, 197–212. [Google Scholar] [CrossRef]
- Shen, M.; Tang, Y.; Chen, J.; Zhu, X.; Zheng, Y. Influences of temperature and precipitation before the growing season on spring phenology in grasslands of the central and eastern Qinghai-Tibetan Plateau. Agric. For. Meterol. 2011, 151, 1711–1722. [Google Scholar] [CrossRef]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Chang. Biol. 2015, 21, 467–3656. [Google Scholar] [CrossRef] [Green Version]
- Bao, F.; He, J.; Cao, Y.; Liu, M.; Xin, Z.; Wu, B. Response and acclimation of photosynthesis in Nitraria tangutorum to rain addition treatments in temperate desert in Northwest China. J. Univ. Chin. Acad. Sci. 2017, 34, 508–514. [Google Scholar]
- Han, J.; Chen, J.; Xia, J.; Li, L. Grazing and watering alter plant phenological processes in a desert steppe community. Plant Ecol. 2015, 216, 599–613. [Google Scholar] [CrossRef]
- Huang, J.; Ji, M.; Xie, Y.; Wang, S.; He, Y.; Ran, J. Global semi-arid climate change over last 60 years. Clim. Dyn. 2015, 46, 1131–1150. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2015, 6, 166–172. [Google Scholar] [CrossRef]
- Gao, X.J.; Shi, Y.; Zhang, D.; Giorgi, F. Climate change in China in the 21st century as simulated by a high-resolution regional climate model. Chin. Sci. Bull. 2012, 57, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Chen, H. Projected change in extreme precipitation events in China by the end of the21st century using CMIP5 models. Chin. Sci. Bull. 2013, 58, 1462–1472. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachaur, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Li, B.; Chen, Y.; Chen, Z.; Xiong, H.; Lian, L. Why does precipitation in northwest China show a significant increasing trend from 1960 to 2010? Atmos. Res. 2016, 167, 275–284. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, B.; Qin, D.; Wu, J.; Gao, R.; Song, L. Changes in mean and extreme temperature and precipitation over the arid region of northwestern China: Observation and projection. Adv. Atmos. Sci. 2017, 34, 289–305. [Google Scholar] [CrossRef]
- Huang, G.; Li, C.H.; Li, Y. Phenological responses to nitrogen and water addition are linked to plant growth patterns in a desert herbaceous community. Ecol. Evol. 2018, 8, 5139–5152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogle, K.; Reynolds, J.F. Plant responses to precipitation in desert ecosystems: Integrating functional types, pulses, thresholds, and delays. Oecologia 2004, 141, 282–294. [Google Scholar] [CrossRef]
- Golluscio, R.A.; Sala, O.E.; Lauenroth, W.K. Differential use of large summer precipitation events by shrubs and grasses: A manipulative experiment in the Patagonian steppe. Oecologia 1998, 115, 17–25. [Google Scholar] [CrossRef]
- Fracheboud, Y.; Luquez, V.; BjÖrken, L.; SjÖdin, A.; Tuominen, H.; Jansson, S. The control of autumn senescence in European aspen. Plant Physiol. 2009, 149, 1982–1991. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wang, X.; Wilsonb, A.M.; Silander, J.A., Jr. Predicting autumn phenology: How deciduous tree species respond to weather stressors. Agric. For. Meteorol. 2018, 250, 127–137. [Google Scholar] [CrossRef]
- Zhou, Y.; Jia, G. Precipitation as a control of vegetation phenology for temperate steppes in China. Atmos. Ocean. Sci. Lett. 2016, 9, 162–168. [Google Scholar] [CrossRef]
- Keenan, T.F.; Richardson, A.D. The timing of autumn senescence is affected by the timing of spring phenology: Implications for predictive models. Glob. Chang. Biol. 2015, 21, 2634–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.S.H.; Campioli, M.; Vitasse, Y.; De Boeck, H.J.; Van den Berge, J.; AbdElgawad, H.; Asard, H.; Piao, S.; Deckmyn, G.; Janssens, I.A. Variation in leaf flushing date influences autumnal senescence and next year’s flushing date in two temperate tree species. Proc. Natl. Acad. Sci. USA 2014, 111, 7355–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Wang, X.; Silander, J.A., Jr. Deciduous forest responses to temperature, precipitation, and drought imply complex climate change impacts. Proc. Natl. Acad. Sci. USA 2015, 112, 13585–13590. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Bao, F.; Wu, B.; Zhang, J.; Jia, Z. Photosynthetic acclimation of a desert species N. tangutorum to long term rain addition in north-western china. Int. J. Agric. Biol. 2019, 22, 1050–1058. [Google Scholar]
- Wan, M.; Liu, X. Plant Phenology Observation Method in China; Science Press: Beijing, China, 1979. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Budding | Leaf Unfolding | Leaf Senescence | Leaf Fall | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | Duration | Start | End | Duration | Start | End | Duration | Start | End | Duration | |
| Year | <0.01 | <0.01 | <0.01 | 0.37 | 0.06 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Treatment | <0.01 | 0.11 | 0.70 | 0.02 | 0.12 | 0.84 | 0.33 | 0.42 | 0.17 | 0.10 | 0.03 | 0.18 |
| Year∗Treatment | 0.97 | 0.97 | 0.93 | 0.59 | 0.77 | 0.29 | 0.06 | 0.50 | 0.74 | 0.97 | 0.01 | 0.89 |
| Year | Treatments | Spring Phenology | Autumn Phenology | ||
|---|---|---|---|---|---|
| Leaf Budding | Leaf Unfolding | Leaf Senescence | Leaf Fall | ||
| 2012 | Control | 24.67 ± 1.26 | 20.50 ± 3.30 | 80.50 ± 0.96 | 88.50 ± 1.26 |
| +25% | 21.00 ± 1.73 | 17.00 ± 3.11 | 84.00 ± 1.63 | 90.50 ± 1.71 | |
| +50% | 19.50 ± 2.50 | 14.00 ± 0.82 | 80.50 ± 3.77 | 90.00 ± 3.16 | |
| +75% | 21.50 ± 3.10 | 20.00 ± 3.56 | 79.75 ± 7.17 | 94.00 ± 3.92 | |
| +100% | 22.00 ± 2.83 | 16.50 ± 2.63 | 87.50 ± 3.20 | 95.00 ± 2.89 | |
| 2013 | Control | 21.50 ± 0.96 | 16.00 ± 1.41 | 76.50 ± 2.75 | 79.75 ± 4.73 |
| +25% | 22.00 ± 0.82 | 14.50 ± 0.50 | 82.50 ± 1.71 | 83.75 ± 1.18 | |
| +50% | 22.00 ± 2.45 | 16.50 ± 1.26 | 83.00 ± 1.29 | 82.00 ± 1.83 | |
| +75% | 23.00 ± 2.38 | 17.50 ± 1.71 | 82.25 ± 2.66 | 85.75 ± 3.64 | |
| +100% | 23.50 ± 2.06 | 18.50 ± 2.63 | 87.50 ± 5.74 | 86.00 ± 2.92 | |
| Phenological Events | Observation Methods | |
|---|---|---|
| Leaf budding | Beginning period | In all leaf buds on the whole sandbag, as long as the leaf budding was observed, the beginning of leaf budding period was recorded. |
| End period | Most of the leaf buds on the whole sandbag had split, and the tips of leaflets were clearly visible. | |
| Leaf unfolding | Beginning period | The beginning of leaf unloading was recorded as long as at least one young leaf had completely extended and spread out completely from one or more leaf buds had been observed on the whole sandbag. |
| End period | More than 90% of the young leaves on leaf buds of the whole sandbag have been completely spread, which is recorded as the end period of leaf unfolding. | |
| Leaf senescence | Beginning period | Approximately 5% of all the leaves on the whole sandbag have begun to turn yellow, which was recorded as the beginning of leaf senescence. |
| End period | More than 90% of the leaves on the whole sandbag have turned yellow, which was recorded as the end period of leaf senescence. | |
| Leaf fall | Beginning period | When approximately 5% of the leaves on the whole sandbag fell off, the beginning of leaf fall was recorded. |
| End period | When more than 90% of the leaves on the whole sandbag had fallen off, the end stage of leaf fall was recorded. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, F.; Xin, Z.; Li, J.; Liu, M.; Cao, Y.; Lu, Q.; Gao, Y.; Wu, B. Effects of the Simulated Enhancement of Precipitation on the Phenology of Nitraria tangutorum under Extremely Dry and Wet Years. Plants 2021, 10, 1474. https://doi.org/10.3390/plants10071474
Bao F, Xin Z, Li J, Liu M, Cao Y, Lu Q, Gao Y, Wu B. Effects of the Simulated Enhancement of Precipitation on the Phenology of Nitraria tangutorum under Extremely Dry and Wet Years. Plants. 2021; 10(7):1474. https://doi.org/10.3390/plants10071474
Chicago/Turabian StyleBao, Fang, Zhiming Xin, Jiazhu Li, Minghu Liu, Yanli Cao, Qi Lu, Ying Gao, and Bo Wu. 2021. "Effects of the Simulated Enhancement of Precipitation on the Phenology of Nitraria tangutorum under Extremely Dry and Wet Years" Plants 10, no. 7: 1474. https://doi.org/10.3390/plants10071474