
Metabolomics Reveals the Allelopathic Potential of the Invasive Plant Eupatorium adenophorum



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
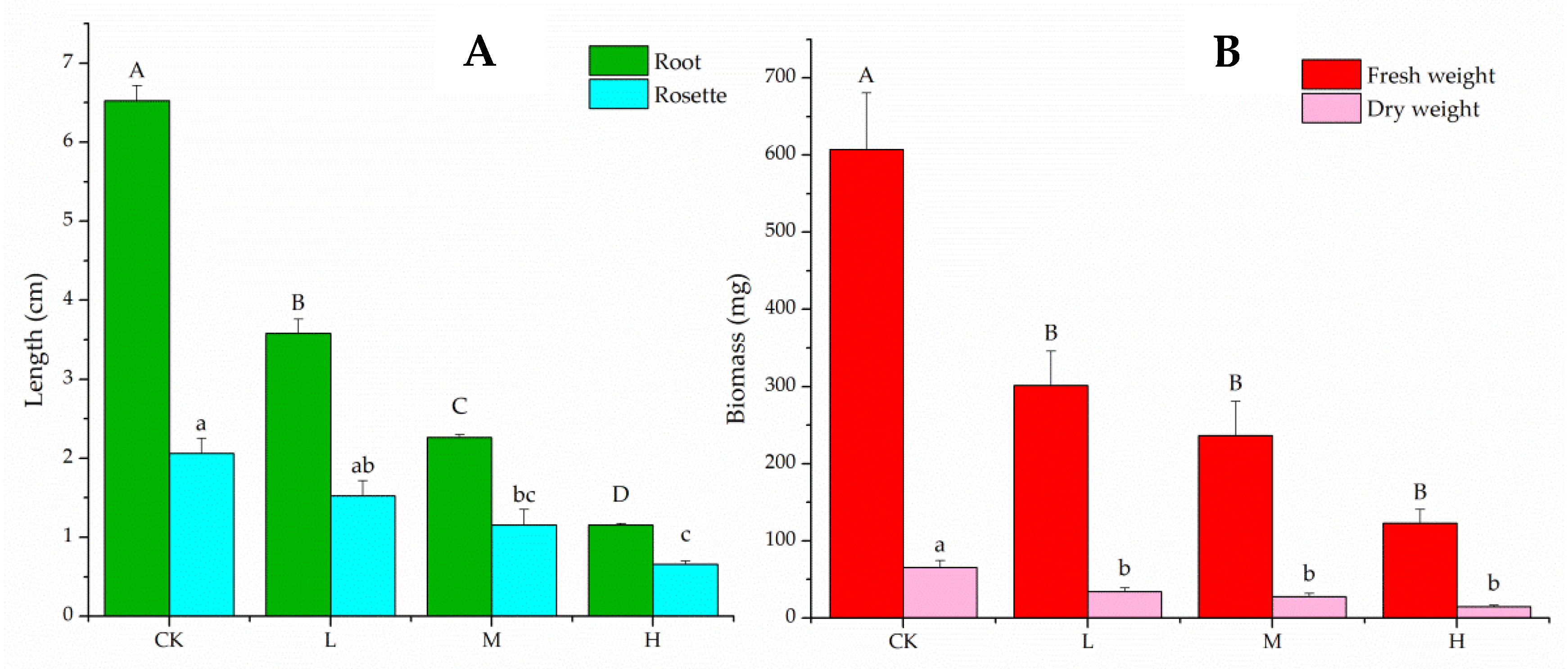
2.1. In Vitro Bioassay
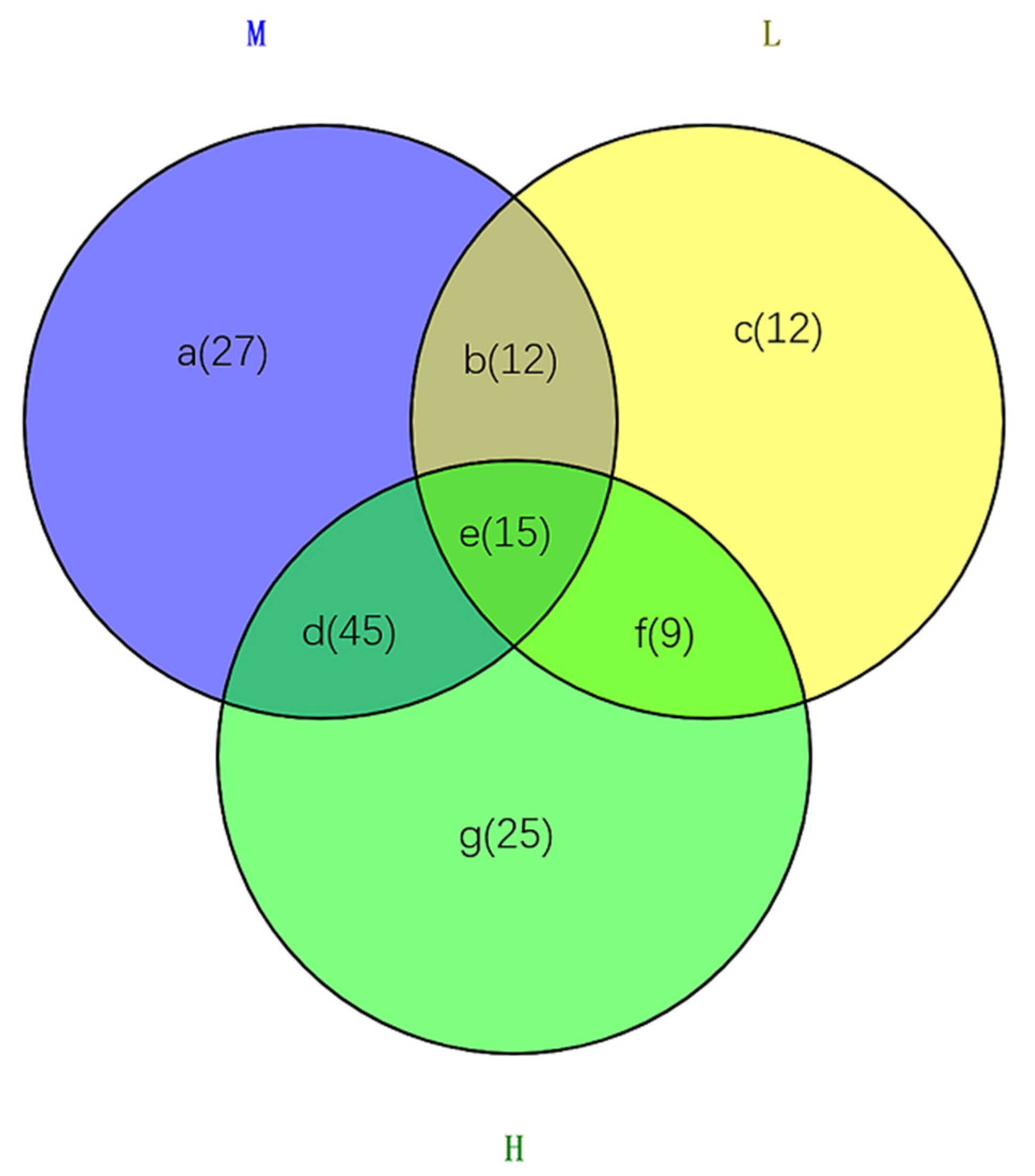
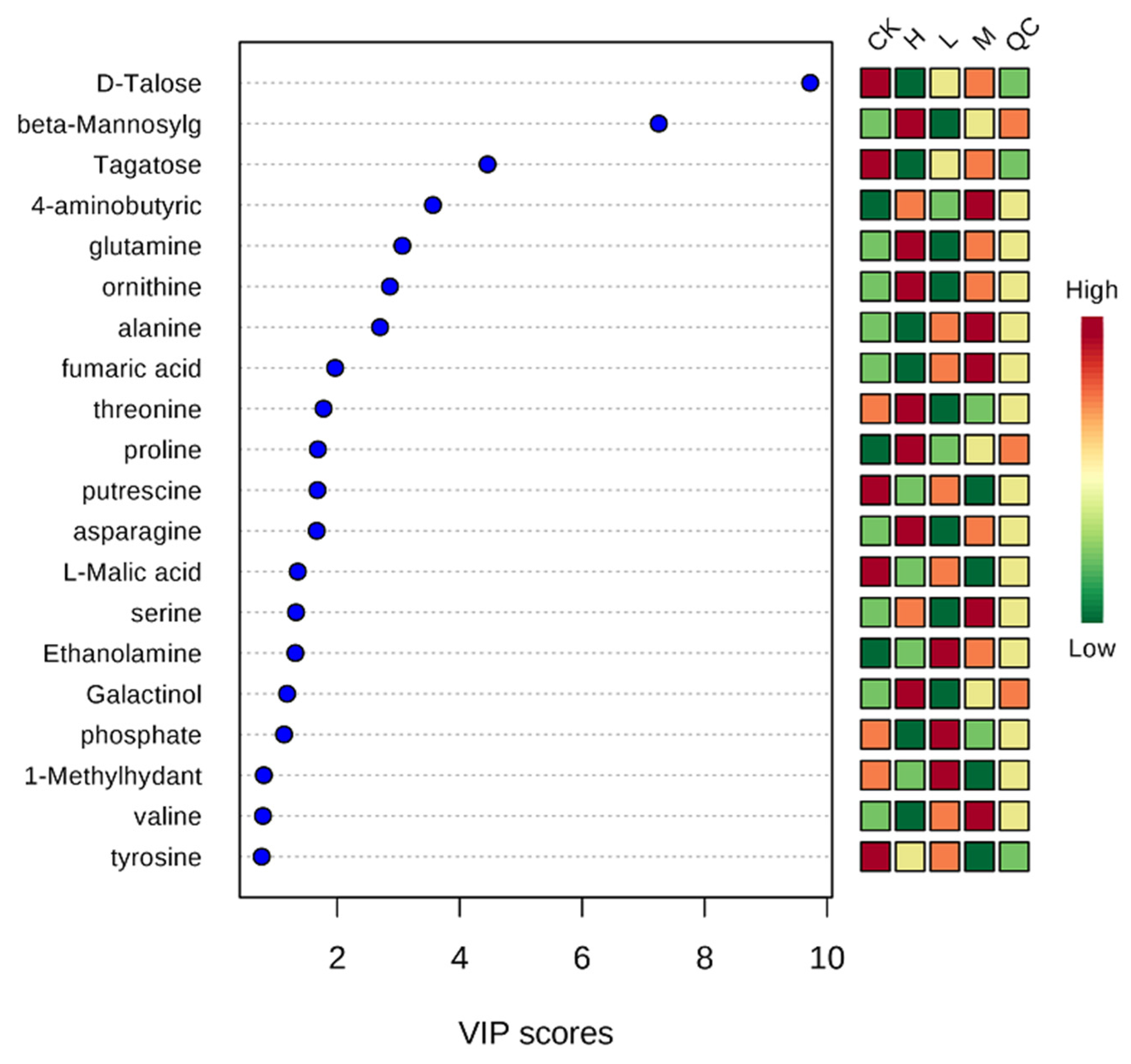
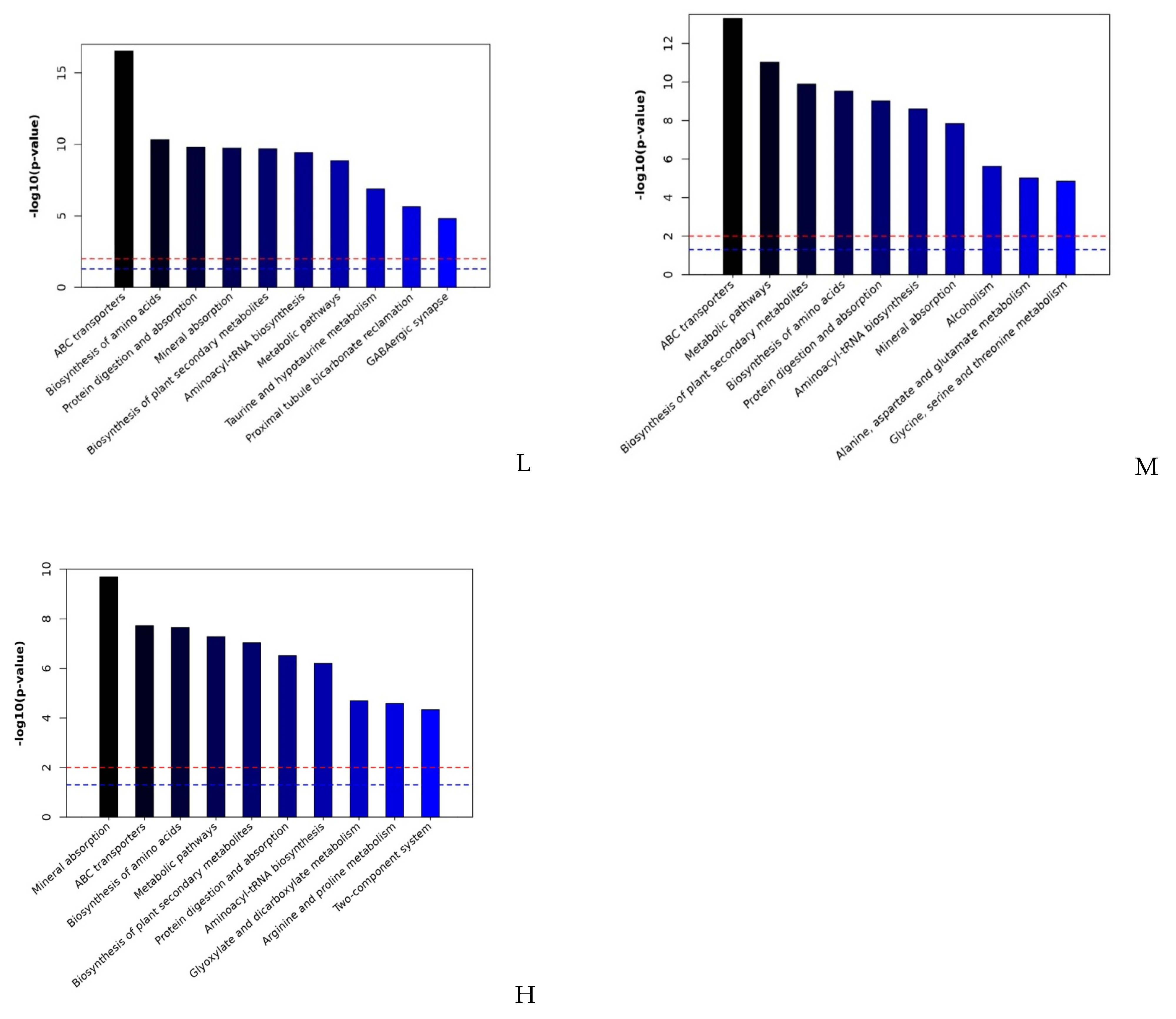
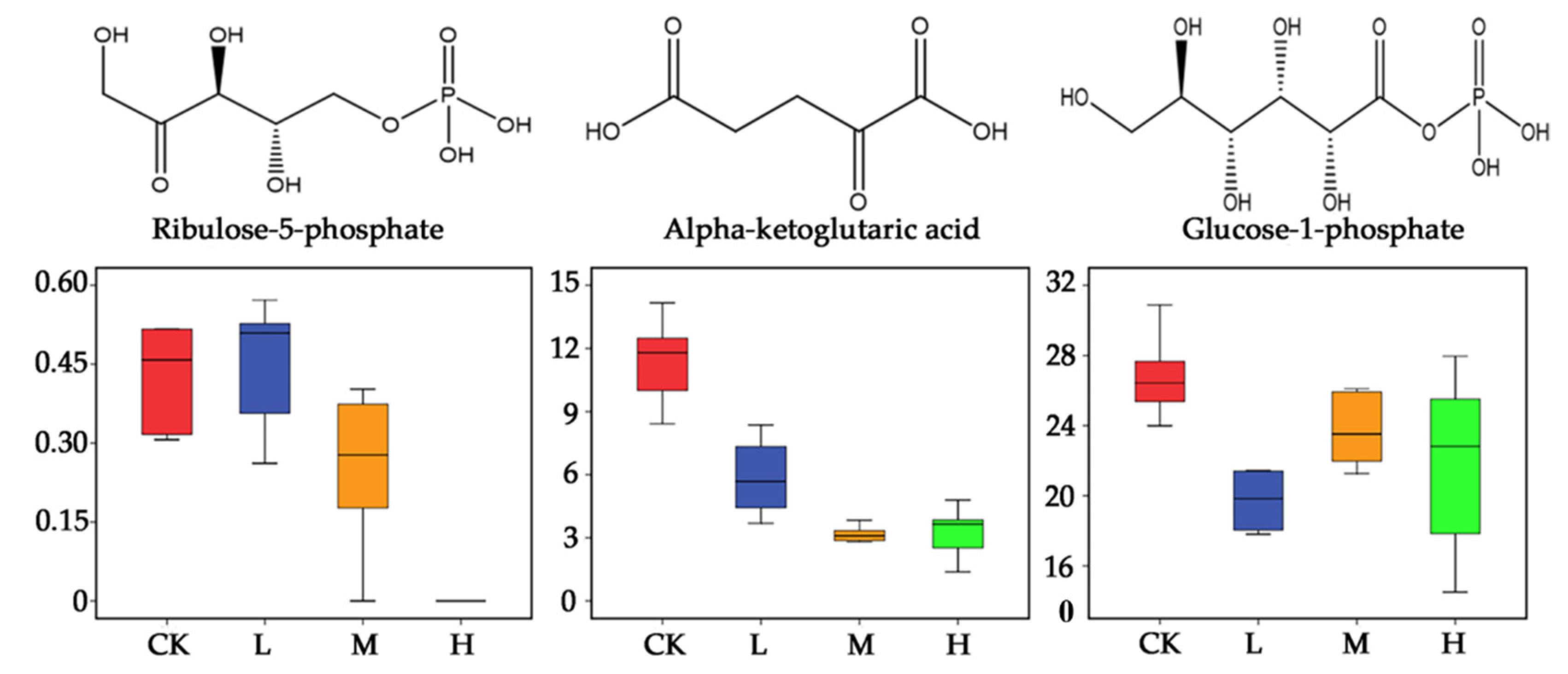
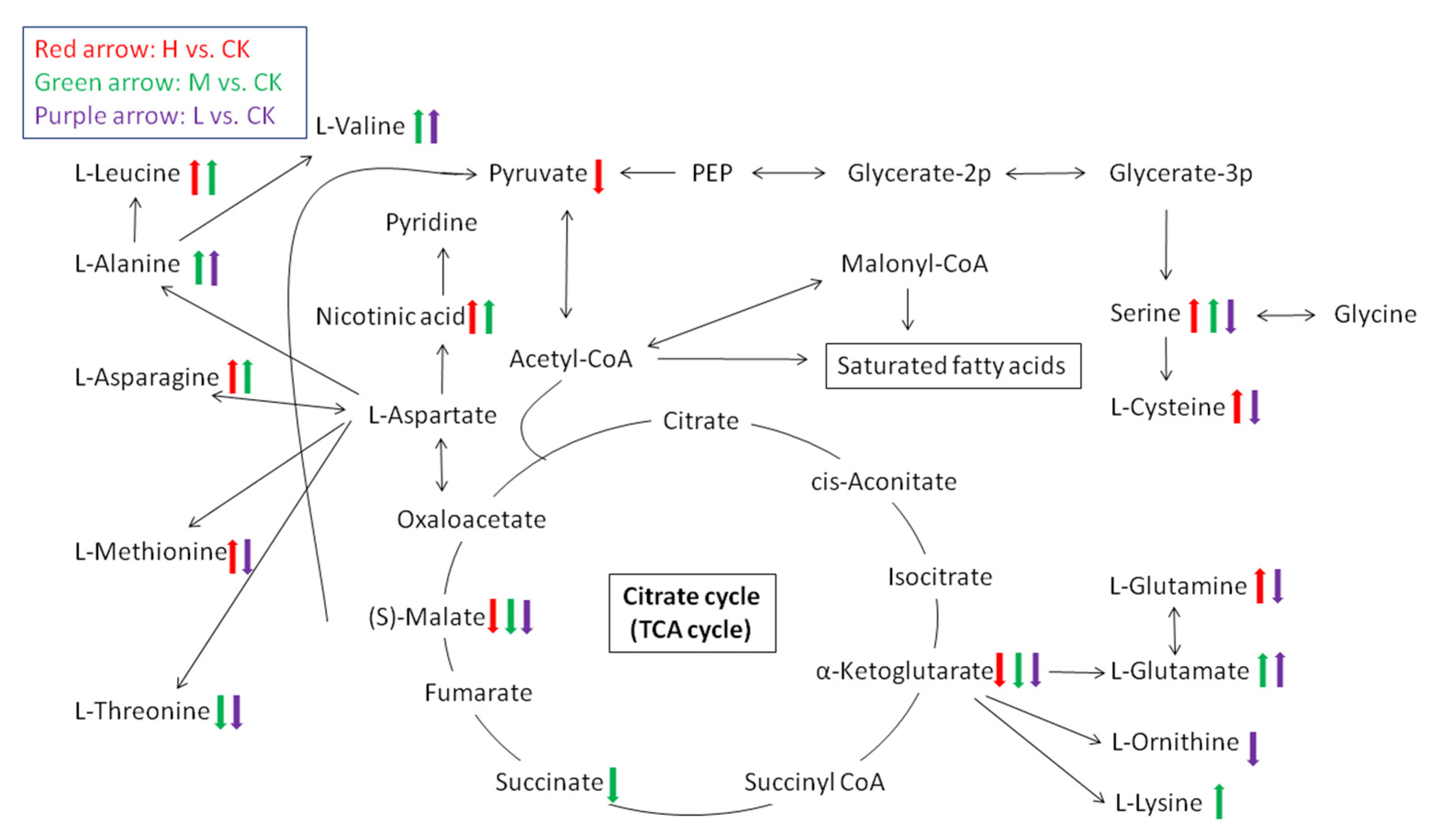
2.2. Metabolite Analysis in Arabidopsis thaliana
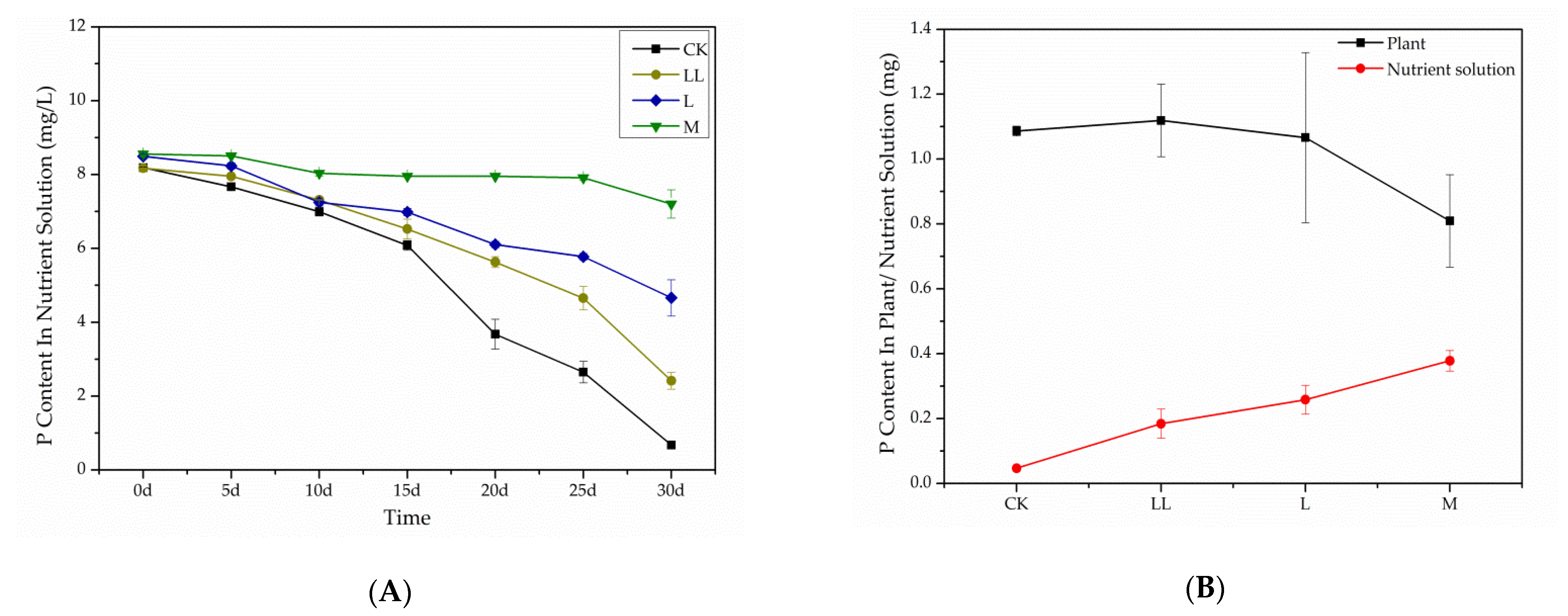
2.3. Phosphate Absorption Experiment
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material Collection and Aqueous Extracts Preparation
5.2. In Vitro Bioassay
5.3. Metabolomic Experiment
5.3.1. GC-MS Sample Preparation
5.3.2. Gas Chromatographic−Mass Spectrometric Analysis
5.3.3. Data Processing
5.4. Phosphate Absorption Experiment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shrestha, K.; Wilson, E.; Gay, H. Ecological and environmental study of Eupatorium adenophorum Sprengel (banmara) with reference to its gall formation in Gorkha-langtang route, Nepal. J. Nat. Hist. Mus. 2009, 23, 108–124. [Google Scholar] [CrossRef] [Green Version]
- Muniappan, R.; Raman, A.; Reddy, G.V.P. Biological Control of Tropical Weeds Using Arthropods, 1st ed.; Cambridge University Press: Cambridge, UK, 2009; pp. 63–73. [Google Scholar]
- Evans, H.; Crocoll, C.; Bajpai, D.; Kaur, R.; Feng, Y.L.; Silva, C.; Trevino, C.J.; Valiente, B.A.; Gershenzon, J. Volatile chemicals from leaf litter are associated with invasiveness of a Neotropical weed in Asia. Ecology 2011, 92, 316–324. [Google Scholar]
- Wang, W.B.; Wang, R.F.; Lei, Y.B.; Liu, C.; Han, L.H.; Shi, X.D.; Feng, Y.L. High resource capture and use efficiency and prolonged growth season contribute to invasiveness of Eupatorium adenophorum. Plant Ecol. 2013, 214, 857–868. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Song, Q.S.; Fu, Y.; Tang, J.W.; Feng, Z.L.; Yang, C.R. Allelopathic potential of Eupatorium adenophorum. Chin. J. Plant Ecol. 2000, 24, 362–365. [Google Scholar]
- Baruah, N.C.; Sarma, J.C.; Sarma, S.; Sharma, R.P. Seed germination and growth inhibitory cadinenes from Eupatorium adenophorum Spreng. J. Chem. Ecol. 1994, 20, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Q.; Wan, F.H.; Liu, W.X.; Zhang, X.W. Physiological effects of allelochemicals from leachates of Ageratina adenophora (Spreng.) on rice seedlings. Allelopath. J. 2006, 18, 237–245. [Google Scholar]
- Yang, G.Q.; Wan, F.H.; Liu, W.X.; Guo, J. Influence of two allelochemicals from Ageratina adenophora Sprengel on ABA, IAA and ZR contents in roots of upland rice seedlings. Allelopath. J. 2008, 21, 253–262. [Google Scholar]
- Liao, F.; Hu, Y.; Tan, H.; Wu, L.; Wang, Y.; Huang, Y.; Mo, Q.; Wei, Y. Acaricidal activity of 9-oxo-10,11-dehydroageraphorone extracted from Eupatorium adenophorum in vitro. Exp. Parasitol. 2014, 140, 8–11. [Google Scholar] [CrossRef]
- Zhang, F.; Guo, J.; Chen, F.; Liu, W.; Wan, F. Identification of Volatile Compounds Released by Leaves of the Invasive Plant Croftonweed (Ageratina adenophora, Compositae), and their Inhibition of Rice Seedling Growth. Weed Sci. 2012, 60, 205–211. [Google Scholar] [CrossRef]
- Gui, F.R.; Jiang, Z.L.; Jin, J.B.; He, S.Q.; Jiang, X.C. Allelopathic effect of Ageratina adenophora on seed germination of nine herbaceous species. J. Biosaf. 2011, 20, 331–336. [Google Scholar]
- Jia, H.J.; Li, X.K.; Tang, S.C.; Tang, S.Q.; Lu, S.H.; Xu, X.L. Allelopathic effects of Eupatorium adenophorum on seed germination of three woody plants in karst region. Guihaia 2009, 29, 631–634. [Google Scholar]
- Yang, G.Q.; Qiu, W.R.; Jin, Y.N.; Wan, F.H. Potential allelochemicals from root exudates of invasive Ageratina adenophora. Allelopath. J. 2013, 32, 233–241. [Google Scholar]
- Yang, G.Q.; Guo, J.; Zhu, X.Z.; Shao, H.; Gao, T. Soil Chemicals from Croftonweed (Ageratina adenophora) Are Phytotoxic. Weed Sci. 2016, 64, 223–230. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, W.X.; Zheng, M.F.; Xu, Q.L.; Wan, F.H.; Wang, J.; Lei, T.; Zhou, Z.Y.; Tan, J.W. Bioactive Quinic Acid Derivatives from Ageratina adenophora. Molecules 2013, 18, 14096–14104. [Google Scholar] [CrossRef] [Green Version]
- Fiehn, O. Metabolomics-the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Rischer, H.; Oksman, C.K.M. Unintended effects in genetically modified crops: Revealed by metabolomics. Trends Biotechnol. 2006, 24, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Furumichi, M.; Tanabe, M.; Hirakawa, M. KEGG for representation and analysis of molecular networks involving diseases and drugs. Nucleic Acids Res. 2010, 38, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.Z.; Zhang, K.M.; Tang, S.J. Allelopathic plants 29: Eupatorium adenophorum Sprengel. Allelopath. J. 2021, 53, 15–22. [Google Scholar] [CrossRef]
- Zhou, Z.Y.; Liu, W.X.; Pei, G.; Ren, H.; Wang, J.; Xu, Q.L.; Xie, H.H.; Wan, F.H.; Tan, J.W. Phenolics from Ageratina adenophora roots and their phytotoxic effects on Arabidopsis thaliana seed germination and seedling growth. Agric. Food Chem. 2013, 61, 11792–11799. [Google Scholar] [CrossRef]
- Zhu, X.Z.; Zhang, J.T.; Ma, K.P. Soil biota reduce allelopathic effects of the invasive Eupatorium adenophorum. PLoS ONE 2011, 6, e25393. [Google Scholar] [CrossRef] [PubMed]
- Florencio, O.V.; Selles, M.S.; Zubcoff, V.J.; Jander, G.; Casas, J.L. Changes in the free amino acid composition of Capsicum annuum (pepper) leaves in response to Myzus persicae (green peach aphid) infestation. A comparison with water stress. PLoS ONE 2018, 13, e0198093. [Google Scholar] [CrossRef] [Green Version]
- Galili, G.; Amir, R.; Fernie, A.R. The regulation of essential amino acid synthesis and accumulation in plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Antikainen, M.; Griffith, M. Antifreeze protein accumulation in freezing-tolerant cereals. Physiol. Plantarum. 1997, 99, 423–432. [Google Scholar] [CrossRef]
- Waters, E.R.; Lee, G.J.; Vierling, E. Evolution, structure and function of the small heat shock proteins in plants. J. Exp. Bot. 1996, 47, 325–338. [Google Scholar] [CrossRef]
- Robinson, S.A.; Stewart, G.R.; Phillips, R. Regulation of glutamate dehydrogenase activity in relation to carbon limitation and protein catabolism in carrot cell suspension cultures. Plant Physiol. 1992, 98, 1190–1195. [Google Scholar] [CrossRef] [Green Version]
- Bourguignon, J.; Rebeille, F.; Douce, R. Serine and Glycine Metabolism in Higher Plants; Plant Amino Acids Biochemistry & Biotechnology Marcel Dekker Inc: New York, NY, USA, 1998; pp. 111–146. [Google Scholar]
- Diaz, C.; Purdy, S.; Christ, A.; Morot-Gaudry, J.-F.; Wingler, A.; Masclaux-Daubresse, C. Characterization of Markers to Determine the Extent and Variability of Leaf Senescence in Arabidopsis. A Metabolic Profiling Approach. Plant Physiol. 2005, 138, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.O.; Oliva, A. Mode of Action of Phytotoxic Terpenoids, 1st ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 201–206. [Google Scholar]
- Araniti, F.; Grana, E.; Krasuska, U.; Bogatek, R.; Reigosa, M.J.; Abenavoli, M.R.; Sanchez, M.A.M. Loss of Gravitropism in Farnesene-Treated Arabidopsis Is Due to Microtubule Malformations Related to Hormonal and ROS Unbalance. PLoS ONE 2016, 11, e0160202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaimovitsh, D.; Abu, A.M.; Belausov, E.; Rubin, B.; Dudai, N.; Sadot, E. Microtubules are an intracellular target of the plant terpene citral. Plant J. 2010, 61, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Sanchez, M.A.M.; Grana, E.; Reigosa, M.J.; Abenavoli, M.R. Terpenoid trans-caryophyllene inhibits weed germination and induces plant water status alteration and oxidative damage in adult Arabidopsis. Plant Biol. 2017, 19, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Grana, E.; Diaz, T.C.; Lopez, G.D.; Martinez, P.A.; Reigosa, M.J.; Sanchez, M.A.M. The plant secondary metabolite citral alters water status and prevents seed formation in Arabidopsis thaliana. Plant Biol. 2016, 18, 423–432. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. Characterization of Xanthophyll Pigments, Photosynthetic Performance, Photon Energy Dissipation, Reactive Oxygen Species Generation and Carbon Isotope Discrimination during Artemisinin-Induced Stress in Arabidopsis thaliana. PLoS ONE 2015, 10, e0114826. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.Z.; Li, Y.P.; Feng, Y.L.; Ma, K.P. Response of soil bacterial communities to secondary compounds released from Eupatorium adenophorum. Biol. Invasions 2017, 19, 1471–1481. [Google Scholar] [CrossRef]
- Zhang, H.L.; Du, W.C.; Peralta, V.J.R.; Gardea, T.J.L.; White, J.C.; Keller, A.; Guo, H.Y.; Ji, R.; Zhao, L.J. Metabolomics Reveals How Cucumber (Cucumis sativus) Reprograms Metabolites to Cope with Silver Ions and Silver Nanoparticle-Induced Oxidative Stress. Environ. Sci. Technol. 2018, 52, 14. [Google Scholar] [CrossRef]
- Zhang, H.; Kariuki, S.; Schroder, J.L.; Payton, M.E.; Focht, C. Interlaboratory Validation of the Mehlich 3 Method for Extraction of Plant-Available Phosphorus. J. AOAC Int. 2009, 92, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, C.C. Direct analysis of plant minerals and comparison of extraction processes using ICP-AES. Food Chem. 1992, 45, 145–149. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Yi, Y.; Huang, L.; Zhang, C.; Shao, H. Metabolomics Reveals the Allelopathic Potential of the Invasive Plant Eupatorium adenophorum. Plants 2021, 10, 1473. https://doi.org/10.3390/plants10071473
Zhu X, Yi Y, Huang L, Zhang C, Shao H. Metabolomics Reveals the Allelopathic Potential of the Invasive Plant Eupatorium adenophorum. Plants. 2021; 10(7):1473. https://doi.org/10.3390/plants10071473
Chicago/Turabian StyleZhu, Xunzhi, Yangmin Yi, Ling Huang, Chi Zhang, and Hua Shao. 2021. "Metabolomics Reveals the Allelopathic Potential of the Invasive Plant Eupatorium adenophorum" Plants 10, no. 7: 1473. https://doi.org/10.3390/plants10071473