
Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles

 ,
,  , ,
, ,  ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Alleviation of Heavy Metal Toxicity in Plants Using Nanoparticles
3. Alleviation of Salinity Stress in Plant Using Nanoparticles
4. Alleviation of Drought Stress in Plants Using Nanoparticles
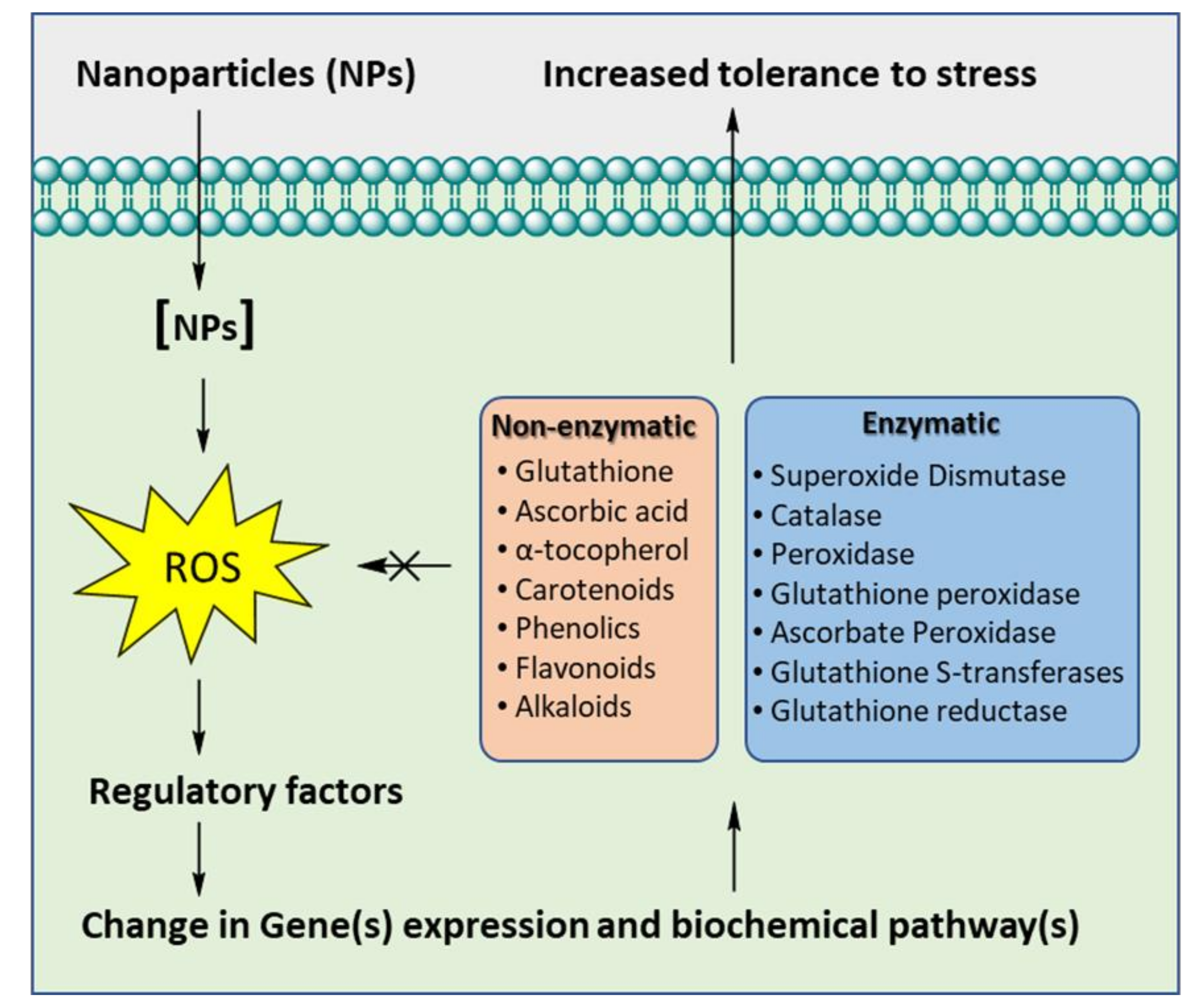
5. Mechanism of Stress Alleviation in Plants under Nanoparticles Application
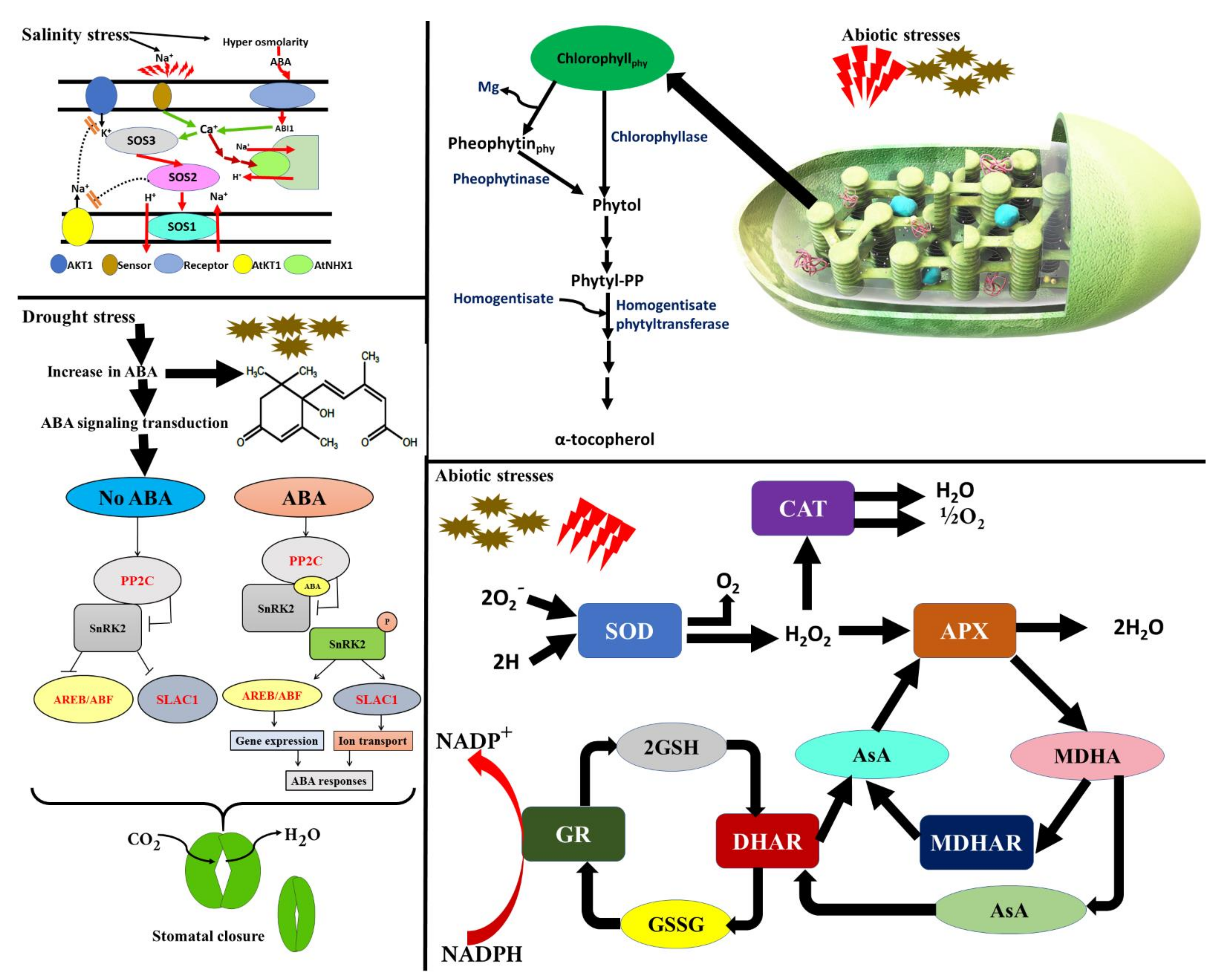
5.1. Impact on Antioxidative Enzymes
5.2. Impact on Cellular Osmolyte Pool
5.3. Molecular Insights
5.4. Impact on Secondary Metabolites
5.5. Activation of Salt Overly Sensitive (SOS) Pathway and Roles of Abscisic Acid under Salt Stress
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Calicioglu, O.; Flammini, A.; Bracco, S.; Bellù, L.; Sims, R. The Future Challenges of Food and Agriculture: An Integrated Analysis of Trends and Solutions. Sustainability 2019, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, S.A.; Siddiqui, A.J.; Elkhalifa, A.E.O.; Khan, M.I.; Patel, M.; Alreshidi, M.; Moin, A.; Singh, R.; Snoussi, M.; Adnan, M. Innovations in Nanoscience dor the Sustainable Development of Food and Agriculture with Implications on Health and Environment. Sci. Total Environ. 2021, 768, 144990. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-Fertilization as an Emerging Fertilization Technique: Why Can Modern Agriculture Benefit from Its Use? Plants 2020, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Rajput, V.D.; Minkina, T.; Bauer, T.; Chauhan, A.; Jindal, T. Nanoparticles Induced Stress and Toxicity in Plants. Environ. Nanotechnol. Monit. Manag. 2021, 15, 100457. [Google Scholar] [CrossRef]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Alam Cheema, S.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium Toxicity in Plants: Impacts and Remediation Strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.; Jogawat, A.; Rahman, S.; Narayan, O.P. Secondary Metabolites in the Drought Stress Tolerance of Crop Plants: A Review. Gene Rep. 2021, 23, 101040. [Google Scholar] [CrossRef]
- Rajput, V.; Harish; Singh, R.; Verma, K.; Sharma, L.; Quiroz-Figueroa, F.; Meena, M.; Gour, V.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Rajput, V.; Minkina, T.; Semenkov, I.; Klink, G.; Tarigholizadeh, S.; Sushkova, S. Phylogenetic Analysis of Hyperaccumulator Plant Species for Heavy Metals and Polycyclic Aromatic Hydrocarbons. Environ. Geochem. Health 2021, 43, 1629–1654. [Google Scholar] [CrossRef]
- Minkina, T.; Rajput, V.; Fedorenko, G.; Fedorenko, A.; Mandzhieva, S.; Sushkova, S.; Morin, T.; Yao, J. Anatomical and Ultrastructural Responses of Hordeum Sativum to the Soil Spiked by Copper. Environ. Geochem. Health 2020, 42, 45–58. [Google Scholar] [CrossRef]
- Verma, K.K.; Anas, M.; Chen, Z.; Rajput, V.D.; Malviya, M.K.; Verma, C.L.; Singh, R.K.; Singh, P.; Song, X.-P.; Li, Y.-R. Silicon Supply Improves Leaf Gas Exchange, Antioxidant Defense System and Growth in Saccharum officinarum Responsive to Water Limitation. Plants 2020, 9, 1032. [Google Scholar] [CrossRef]
- Ayup, M.; Chen, Y.-N.; Nyongesah, M.J.; Zhang, Y.-M.; Rajput, V.D.; Zhu, C.-G. Xylem Anatomy and Hydraulic Traits of Two Co-Occurring Riparian Desert Plants. IAWA J. 2015, 36, 69–83. [Google Scholar] [CrossRef]
- Zulkafflee, N.S.; Redzuan, N.A.M.; Selamat, J.; Ismail, M.R.; Praveena, S.M.; Razis, A.F.A. Evaluation of Heavy Metal Contamination in Paddy Plants at the Northern Region of Malaysia Using ICPMS and Its Risk Assessment. Plants 2020, 10, 3. [Google Scholar] [CrossRef]
- Li, Q.; Li, C.; Wang, H.; Wei, X.; Liu, Y.; Yang, R.; Wen, X. Geochemical Characteristics of Heavy Metals in Soil and Blueberries of the Core Majiang Blueberry Production Area. Bull. Environ. Contam. Toxicol. 2021, 106, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.; Minkina, T.; Mazarji, M.; Shende, S.; Sushkova, S.; Mandzhieva, S.; Burachevskaya, M.; Chaplygin, V.; Singh, A.; Jatav, H. Accumulation of Nanoparticles in the Soil-plant Systems and Their Sffects on Human Health. Ann. Agric. Sci. 2020, 65, 137–143. [Google Scholar] [CrossRef]
- Chaplygin, V.A.; Rajput, V.D.; Mandzhieva, S.S.; Minkina, T.M.; Nevidomskaya, D.G.; Nazarenko, O.G.; Kalinitchenko, V.P.; Singh, R.; Maksimov, A.Y.; Popova, V.A. Comparison of Heavy Metal Content in Artemisia austriaca in Various Impact Zones. ACS Omega 2020, 5, 23393–23400. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Manikandan, R.; Favas, P.J.; Sahi, S.V.; Venkatachalam, P. Effect of Lead on Phytotoxicity, Growth, Biochemical Alterations and Its Role on Genomic Template Stability in Sesbania grandiflora: A Potential Plant for Phytoremediation. Ecotoxicol. Environ. Saf. 2014, 108, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, H.; Pour, B.E.; Rizwan, M. Foliar Application of Silicon Nanoparticles Affected the Growth, Vitamin C, Flavonoid, and Antioxidant Enzyme Activities of Coriander (Coriandrum sativum L.) Plants Grown in Lead (Pb)-Spiked Soil. Environ. Sci. Pollut. Res. 2021, 28, 1417–1425. [Google Scholar] [CrossRef]
- Bidi, H.; Fallah, H.; Niknejad, Y.; Tari, D.B. Iron Oxide Nanoparticles Alleviate Arsenic Phytotoxicity In Rice by Improving Iron Uptake, Oxidative Stress Tolerance and Diminishing Arsenic Accumulation. Plant Physiol. Biochem. 2021, 163, 348–357. [Google Scholar] [CrossRef]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.-T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. Int. J. Mol. Sci. 2019, 21, 148. [Google Scholar] [CrossRef] [Green Version]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant Survival Under Drought Stress: Implications, Adaptive Responses, and Integrated Rhizosphere Management Strategy for Stress Mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Rajput, V.D.; Yaning, C.; Ayup, M.; Minkina, T.; Sushkova, S.; Mandzhieva, S. Physiological and Hydrological Changes in Populus euphratica Seedlings Under Salinity Stress. Acta Ecol. Sin. 2017, 37, 229–235. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the Physiological and Biochemical Impacts of Salt Stress on Plant Growth and Development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Yaning, C.; Sushkova, S.; Chapligin, V.A.; Mandzhieva, S. A Review on Salinity Adaptation Mechanism and Characteristics of Populus euphratica, A Boon for Arid Ecosystems. Acta Ecol. Sin. 2016, 36, 497–503. [Google Scholar] [CrossRef]
- Ibrahim, W.; Zhu, Y.-M.; Chen, Y.; Qiu, C.-W.; Zhu, S.; Wu, F.; Shuijin, Z. Genotypic Differences in Leaf Secondary Metabolism, Plant Hormones and Yield Under Alone and Combined Stress of Drought and Salinity In Cotton Genotypes. Physiol. Plant 2018, 165, 343–355. [Google Scholar] [CrossRef]
- Rajput, V.; Minkina, T.; Ahmed, B.; Sushkova, S.; Singh, R.; Soldatov, M.; Laratte, B.; Fedorenko, A.; Mandzhieva, S.; Blicharska, E.; et al. Interaction of Copper-Based Nanoparticles to Soil, Terrestrial, and Aquatic Systems: Critical Review of the State of the Science and Future Perspectives. In Reviews of Environmental Contamination and Toxicology; de Voogt, P., Ed.; Springer: Cham, Switzerland, 2019; Volume 250, pp. 51–96. [Google Scholar]
- Pelegrino, M.T.; Kohatsu, M.Y.; Seabra, A.B.; Monteiro, L.R.; Gomes, D.G.; Oliveira, H.C.; Rolim, W.R.; De Jesus, T.A.; Batista, B.L.; Lange, C.N. Effects of Copper Oxide Nanoparticles on Growth of Lettuce (Lactuca sativa L.) Seedlings and Possible Implications of Nitric Oxide in Their Antioxidative Defense. Environ. Monit. Assess. 2020, 192, 1–14. [Google Scholar] [CrossRef]
- Rajput, V.; Chaplygin, V.; Gorovtsov, A.; Fedorenko, A.; Azarov, A.; Chernikova, N.; Barakhov, A.; Minkina, T.; Maksimov, A.; Mandzhieva, S.; et al. Assessing the Toxicity and Accumulation of Bulk- and Nano-Cuo in Hordeum sativum L. Environ. Geochem. Health 2020, 43, 2443–2454. [Google Scholar] [CrossRef]
- Youssef, M.S.; Elamawi, R. Evaluation of Phytotoxicity, Cytotoxicity, and Genotoxicity of ZnO Nanoparticles in Vicia faba. Environ. Sci. Pollut. Res. 2018, 27, 18972–18984. [Google Scholar] [CrossRef]
- Adrees, M.; Khan, Z.S.; Ali, S.; Hafeez, M.; Khalid, S.; Rehman, M.Z.U.; Hussain, A.; Hussain, K.; Chatha, S.A.S.; Rizwan, M. Simultaneous Mitigation of Cadmium and Drought Stress in Wheat by Soil Application of Iron Nanoparticles. Chemosphere 2020, 238, 124681. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikov, S.; Tsepina, N.; Minnikova, T.; Kazeev, K.; Mandzhieva, S.; Sushkova, S.; Minkina, T.; Mazarji, M.; Singh, R.; Rajput, V. Influence of Silver Nanoparticles on the Biological Indicators of Haplic Chernozem. Plants 2021, 10, 1022. [Google Scholar] [CrossRef]
- Arya, A.; Mishra, V.; Chundawat, T.S. Green Synthesis of Silver Nanoparticles from Green Algae (Botryococcus braunii) and Its Catalytic Behavior for the Synthesis of Benzimidazoles. Chem. Data Collect. 2019, 20, 100190. [Google Scholar] [CrossRef]
- Staroń, A.; Długosz, O.; Pulit-Prociak, J.; Banach, M. Analysis of the Exposure of Organisms to the Action of Nanomaterials. Materials 2020, 13, 349. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of Carbon Nanomaterials in the Plant System: A Perspective View on the Pros and Cons. Sci. Total Environ. 2019, 667, 485–499. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Potentials of Engineered Nanoparticles as Fertilizers for Increasing Agronomic Productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Lombi, E.; Zhao, F.-J.; Kopittke, P.M. Nanotechnology: A New Opportunity in Plant Sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, A.; Nangia, A.; Prasad, M. A Green Synthetic Route to Phenolics Fabricated Magnetite Nanoparticles From Coconut Husk Extract: Implications to Treat Metal Contaminated Water and Heavy Metal Stress in Oryza sativa L. J. Clean. Prod. 2018, 174, 355–366. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Hussain, A.; Rehman, M.Z.U.; Ali, B.; Yousaf, B.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Silicon Nanoparticles Enhanced the Growth and Reduced the Cadmium Accumulation in Grains of Wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2019, 140, 1–8. [Google Scholar] [CrossRef]
- Singh, D.; Singh, S.K.; Singh, V.K.; Verma, H.; Mishra, M.; Rashmi, K.; Kumar, A. Plant Growth-promoting Bacteria: Application in Bioremediation of Salinity and Heavy Metal–Contaminated Soils. In Microbe Mediated Remediation of Environmental Contaminants; Kumar, A., Singh, V.K., Singh, P., Mishra, V.K., Eds.; Woodhead Publishing: Sawston, UK, 2021; pp. 73–78. [Google Scholar]
- Tombuloglu, H.; Slimani, Y.; Tombuloglu, G.; Alshammari, T.; Almessiere, M.; Korkmaz, A.D.; Baykal, A.; Samia, A.C.S. Engineered Magnetic Nanoparticles Enhance Chlorophyll Content and Growth of Barley Through the Induction of Photosystem Genes. Environ. Sci. Pollut. Res. 2020, 27, 34311–34321. [Google Scholar] [CrossRef]
- Ali, E.; El-Shehawi, A.; Ibrahim, O.; Abdul-Hafeez, E.; Moussa, M.; Hassan, F. A Vital Role of Chitosan Nanoparticles in Improvisation the Drought Stress Tolerance in Catharanthus roseus (L.) Through Biochemical and Gene Expression Modulation. Plant Physiol. Biochem. 2021, 161, 166–175. [Google Scholar] [CrossRef]
- Singh, D.; Sillu, D.; Kumar, A.; Agnihotri, S. Dual Nanozyme Characteristics of Iron Oxide Nanoparticles Alleviate Salinity Stress and Promote the Growth of An Agroforestry Tree, Eucalyptus tereticornis Sm. Environ. Sci. Nano 2021, 8, 1308–1325. [Google Scholar] [CrossRef]
- Chaplygin, V.; Mandzhieva, S.; Minkina, T.; Barahov, A.; Nevidomskaya, D.; Kizilkaya, R.; GULSER, C.; Chernikova, N.; Mazarji, M.; Iljina, L.; et al. Accumulating Capacity of Herbaceous Plants of the Asteraceae and Poaceae Families Under Technogenic Soil Pollution with Zinc and Cadmium. Eeurasian J. Soil Sci. 2020, 9, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Ghazaryan, K.; Movsesyan, H.; Gevorgyan, A.; Minkina, T.; Sushkova, S.; Rajput, V.; Mandzhieva, S. Comparative Hydrochemical Assessment of Groundwater Quality from Different Aquifers for Irrigation Purposes Using IWQI: A Case-Study from Masis Province in Armenia. Groundw. Sustain. Dev. 2020, 11, 100459. [Google Scholar] [CrossRef]
- Ennaji, W.; Barakat, A.; El Baghdadi, M.; Rais, J. Heavy Metal Contamination in Agricultural Soil and Ecological Risk Assessment in The Northeast Area of Tadla Plain, Morocco. J. Sediment. Environ. 2020, 5, 307–320. [Google Scholar] [CrossRef]
- Zamora-Ledezma, C.; Negrete-Bolagay, D.; Figueroa, F.; Zamora-Ledezma, E.; Ni, M.; Alexis, F.; Guerrero, V.H. Heavy Metal Water Pollution: A Fresh Look About Hazards, Novel and Conventional Remediation Methods. Environ. Technol. Innov. 2021, 22, 101504. [Google Scholar] [CrossRef]
- Galati, S.; Gullì, M.; Giannelli, G.; Furini, A.; DalCorso, G.; Fragni, R.; Buschini, A.; Visioli, G. Heavy Metals Modulate DNA Compaction and Methylation at CpG Sites in the Metal Hyperaccumulator Arabidopsis halleri. Environ. Mol. Mutagen. 2021, 62, 133–142. [Google Scholar] [CrossRef]
- Kontaş, S.; Bostancı, D. Genotoxic Effects of Environmental Pollutant Heavy Metals on Alburnus chalcoides (Pisces: Cyprinidae) Inhabiting Lower Melet River (Ordu, Turkey). Bull. Environ. Contam. Toxicol. 2020, 104, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Sall, M.L.; Diaw, A.K.D.; Gningue-Sall, D.; Aaron, S.E.; Aaron, J.-J. Toxic Heavy Metals: Impact on the Environment and Human Health, and Treatment with Conducting Organic Polymers, A Review. Environ. Sci. Pollut. Res. 2020, 27, 29927–29942. [Google Scholar] [CrossRef]
- Raja, V.; Lakshmi, R.V.; Sekar, C.P.; Chidambaram, S.; Neelakantan, M.A. Health Risk Assessment of Heavy Metals in Groundwater of Industrial Township Virudhunagar, Tamil Nadu, India. Arch. Environ. Contam. Toxicol. 2021, 80, 144–163. [Google Scholar] [CrossRef]
- Zhao, Q.; Bai, J.; Gao, Y.; Zhang, G.; Lu, Q.; Jia, J. Heavy Metal Contamination in Soils from Freshwater Wetlands to Salt Marshes in the Yellow River Estuary, China. Sci. Total Environ. 2021, 774, 145072. [Google Scholar] [CrossRef]
- Alaboudi, K.A.; Ahmed, B.; Brodie, G. Phytoremediation of Pb and Cd Contaminated Soils by Using Sunflower (Helianthus annuus) Plant. Ann. Agric. Sci. 2018, 63, 123–127. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Abdullah Alsahli, A.; Jan, S.; Ahmad, P. Seed Priming with Titanium Dioxide Nanoparticles Enhances Seed Vigor, Leaf Water Status, and Antioxidant Enzyme Activities in Maize (Zea mays L.) under salinity stress. J. King Saud. Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- Shah, V.; Daverey, A. Effects of Sophorolipids Augmentation on the Plant Growth and Phytoremediation of Heavy Metal Contaminated Soil. J. Clean. Prod. 2021, 280, 124406. [Google Scholar] [CrossRef]
- Chen, S.; Chen, L.; Ma, Y.; Huang, Y. Can Phosphate Compounds be Used to Reduce the Plant Uptake of Pb and Resist the Pb Stress in Pb-Contaminated Soils? J. Environ. Sci. 2009, 21, 360–365. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Chen, S.; Ma, Y. Alleviation of Cadmium-Induced Root Growth Inhibition in Crop Seedlings by Nanoparticles. Ecotoxicol. Environ. Saf. 2012, 79, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.; Shahid, M.; Ahmed, T.; Tahir, M.; Naqqash, T.; Muhammad, S.; Song, F.; Abid, H.M.A.; Aslam, Z. Green Copper Nanoparticles drom A Native Klebsiella Pneumoniae Strain Alleviated Oxidative Stress Impairment of Wheat Plants by Reducing the Chromium Bioavailability and Increasing the Growth. Ecotoxicol. Environ. Saf. 2020, 192, 110303. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.; Ahmed, T.; Hussain, S.; Niazi, M.B.K.; Shahid, M.; Song, F. Biogenic Copper Nanoparticles Synthesized by Using a Copper-Resistant Strain Shigella Flexneri Snt22 Reduced the Translocation of Cadmium from Soil to Wheat Plants. J. Hazard. Mater. 2020, 398, 123175. [Google Scholar] [CrossRef]
- Ahmed, T.; Noman, M.; Manzoor, N.; Shahid, M.; Abdullah, M.; Ali, L.; Wang, G.; Hashem, A.; Al-Arjani, A.-B.F.; Alqarawi, A.A.; et al. Nanoparticle-Based Amelioration of Drought Stress and Cadmium Toxicity in Rice via Triggering the Stress Responsive Genetic Mechanisms and Nutrient Acquisition. Ecotoxicol. Environ. Saf. 2021, 209, 111829. [Google Scholar] [CrossRef]
- Manzoor, N.; Ahmed, T.; Noman, M.; Shahid, M.; Nazir, M.M.; Ali, L.; Alnusaire, T.S.; Li, B.; Schulin, R.; Wang, G. Iron Oxide Nanoparticles Ameliorated the Cadmium and Salinity Stresses in Wheat Plants, Facilitating Photosynthetic Pigments and Restricting Cadmium Uptake. Sci. Total Environ. 2021, 769, 145221. [Google Scholar] [CrossRef]
- Zhou, P.; Adeel, M.; Shakoor, N.; Guo, M.; Hao, Y.; Azeem, I.; Li, M.; Liu, M.; Rui, Y. Application of Nanoparticles Alleviates Heavy Metals Stress and Promotes Plant Growth: An Overview. Nanomaterials 2020, 11, 26. [Google Scholar] [CrossRef]
- Lian, J.; Zhao, L.; Wu, J.; Xiong, H.; Bao, Y.; Zeb, A.; Tang, J.; Liu, W. Foliar Spray of TiO2 Nanoparticles Prevails Over Root Application in Reducing Cd Accumulation and Mitigating Cd-Induced Phytotoxicity in Maize (Zea mays L.). Chemosphere 2020, 239, 124794. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, N.; Liang, X.; Bai, X.; Zheng, L.; Zhao, J.; Li, Y.-F.; Zhang, Z.; Gao, Y. Silica Nanoparticles Alleviate Mercury Toxicity via Immobilization and Inactivation of Hg(Ii) in Soybean (Glycine max). Environ. Sci. Nano 2020, 7, 1807–1817. [Google Scholar] [CrossRef]
- Jiang, M.; Dai, S.; Wang, B.; Xie, Z.; Li, J.; Wang, L.; Li, S.; Tan, Y.; Tian, B.; Shu, Q.; et al. Gold Nanoparticles Synthesized Using Melatonin Suppress Cadmium Uptake and Alleviate Its Toxicity in Rice. Environ. Sci. Nano 2021, 8, 1042–1056. [Google Scholar] [CrossRef]
- Khan, Z.S.; Rizwan, M.; Hafeez, M.; Ali, S.; Adrees, M.; Qayyum, M.F.; Khalid, S.; Rehman, M.Z.U.; Sarwar, M.A. Effects of silicon nanoparticles on growth and physiology of wheat in cadmium contaminated soil under different soil moisture levels. Environ. Sci. Pollut. Res. 2020, 27, 4958–4968. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Alyemeni, M.N.; Al-Huqail, A.A.; Alqahtani, M.A.; Wijaya, L.; Ashraf, M.; Kaya, C.; Bajguz, A. Zinc Oxide Nanoparticles Application Alleviates Arsenic (As) Toxicity in Soybean Plants by Restricting the Uptake of as and Modulating Key Biochemical Attributes, Antioxidant Enzymes, Ascorbate-Glutathione Cycle and Glyoxalase System. Plants 2020, 9, 825. [Google Scholar] [CrossRef]
- Katiyar, P.; Yadu, B.; Korram, J.; Satnami, M.L.; Kumar, M.; Keshavkant, S. Titanium Nanoparticles Attenuates Arsenic Toxicity by Up-Regulating Expressions of Defensive Genes in Vigna radiata L. J. Environ. Sci. 2020, 92, 18–27. [Google Scholar] [CrossRef]
- Hussain, B.; Lin, Q.; Hamid, Y.; Sanaullah, M.; Di, L.; Hashmi, M.L.U.R.; Khan, M.B.; He, Z.; Yang, X. Foliage Application of Selenium and Silicon Nanoparticles Alleviates Cd And Pb Toxicity in Rice (Oryza sativa L.). Sci. Total Environ. 2020, 712, 136497. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon Nanoparticles (SiNP) Alleviate Chromium (VI) Phytotoxicity in Pisum sativum (L.) Seedlings. Plant Physiol. Biochem. 2015, 96, 189–198. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.; Sahi, S. Zinc Oxide Nanoparticles (ZnO NPS) Alleviate Heavy Metal-Induced Toxicity in Leucaena Leucocephala Seedlings: A Physiochemical Analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef]
- Konate, A.; He, X.; Zhang, Z.; Ma, Y.; Zhang, P.; Alugongo, G.M.; Rui, Y. Magnetic (Fe3O4) Nanoparticles Reduce Heavy Metals Uptake and Mitigate Their Toxicity in Wheat Seedling. Sustainability 2017, 9, 790. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Liu, T.; Li, F.; Yi, J.; Liu, C.; Yu, H.-Y. Silica Nanoparticles Alleviate Cadmium Toxicity in Rice Cells: Mechanisms and Size Effects. Environ. Pollut. 2017, 228, 363–369. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zia Ur Rehman, M.Z.U.; Adrees, M.; Arshad, M.; Qayyum, M.F.; Ali, L.; Hussain, A.; Chatha, S.A.S.; Imran, M. Alleviation of Cadmium Accumulation in Maize (Zea mays L.) by Foliar Spray of Zinc Oxide Nanoparticles and Biochar to Contaminated Soil. Environ. Pollut. 2019, 248, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Rehman, M.Z.U.; Waris, A.A. Zinc and Iron Oxide Nanoparticles Improved the Plant Growth and Reduced the Oxidative Stress and Cadmium Concentration in Wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wu, F.; Zhou, S.; Yang, J.; Tang, X.; Ye, W. Zinc Oxide Nanoparticles Alleviate the Arsenic Toxicity and Decrease the Accumulation of Arsenic in Rice (Oryza sativa L.). BMC Plant Biol. 2021, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Shelia, V.; Ludwig, F.; de Bruyn, L.L.; Rahman, M.H.U.; Hoogenboom, G. Bringing Farmers’ Perceptions into Science and Policy: Understanding Salinity Tolerance of Rice in Southwestern Bangladesh Under Climate Change. Land Use Policy 2021, 101, 105159. [Google Scholar] [CrossRef]
- Ahmad, I.; Akhtar, M.S. Use of Nanoparticles in Alleviating Salt Stress. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Akhtar, M.S., Ed.; Springer: Singapore, 2019; Volume 1, pp. 199–215. [Google Scholar]
- Khadka, K.; Earl, H.J.; Raizada, M.N.; Navabi, A. A Physio-Morphological Trait-Based Approach for Breeding Drought Tolerant Wheat. Front. Plant Sci. 2020, 11, 715. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- El-Saber, M.M.; A Mahdi, A.; Hassan, A.H.; Farroh, K.Y.; Osman, A. Effects of Magnetite Nanoparticles on Physiological Processes to Alleviate Salinity Induced Oxidative Damage in Wheat. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef] [PubMed]
- Farhangi-Abriz, S.; Torabian, S. Nano-Silicon Alters Antioxidant Activities of Soybean Seedlings Under Salt Toxicity. Protoplasma 2018, 255, 953–962. [Google Scholar] [CrossRef]
- Mushtaq, A.; Rizwan, S.; Jamil, N.; Ishtiaq, T.; Irfan, S.; Ismail, T.; Malghani, M.N.; Shahwani, M.N. Influence of Silicon Sources and Controlled Release Fertilizer on the Growth of Wheat Cultivars of Balochistan Under Salt Stress. Pak. J. Bot. 2019, 51, 1561–1567. [Google Scholar] [CrossRef]
- Farouk, S.; Al-Amri, S.M. Exogenous Zinc Forms Counteract NaCl-Induced Damage by Regulating the Antioxidant System, Osmotic Adjustment Substances, and Ions in Canola (Brassica napus L. cv. Pactol) Plants. J. Soil Sci. Plant Nutr. 2019, 19, 887–899. [Google Scholar] [CrossRef]
- Pérez-Labrada, F.; López-Vargas, E.R.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Responses of Tomato Plants under Saline Stress to Foliar Application of Copper Nanoparticles. Plants 2019, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.K.S.H.; Qayyum, M.F.; Abdel-Hadi, A.; Rehman, R.A.; Ali, S.; Rizwan, M. Interactive Effect of Salinity and Silver Nanoparticles on Photosynthetic and Biochemical Parameters of Wheat. Arch. Agron. Soil Sci. 2017, 63, 1736–1747. [Google Scholar] [CrossRef]
- Abdoli, S.; Ghassemi-Golezani, K.; Alizadeh-Salteh, S. Responses of Ajowan (Trachyspermum ammi L.) to Exogenous Salicylic Acid and Iron Oxide Nanoparticles under Salt Stress. Environ. Sci. Pollut. Res. 2020, 27, 36939–36953. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.W.M.; Abdeldaym, E.A.; Abdelaziz, S.M.; El-Sawy, M.B.I.; Mottaleb, S.A. Synergetic Effects of Zinc, Boron, Silicon, and Zeolite Nanoparticles on Confer Tolerance in Potato Plants Subjected to Salinity. Agronomy 2019, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Mozafari, A.-A.; Ghadakchi Asl, A.G.; Ghaderi, N. Grape Response to Salinity Stress and Role of Iron Nanoparticle and Potassium Silicate to Mitigate Salt Induced Damage Under in vitro Conditions. Physiol. Mol. Biol. Plants 2018, 24, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, A.A.; Dedejani, S.; Ghaderi, N. Positive Responses of Strawberry (Fragaria × ananassa Duch.) Explants to Salicylic and Iron Nanoparticle Application under Salinity Conditions. Plant Cell Tissue Organ Cult. 2018, 134, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Kafi, M.; Nabati, J.; Saadatian, B.; Oskoueian, A.; Shabahang, J. Potato Response to Silicone Compounds (Micro and Nanoparticles) and Potassium as Affected by Salinity Stress. Ital. J. Agron. 2019, 14, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, L.M.; Dutt, M.; Shalan, A.M.; El-Kady, M.E.; El-Boray, M.S.; Shabana, Y.; Grosser, J.W. Silicon Nanoparticles Mitigate Oxidative Stress of in vitro-Derived Banana (Musa acuminata ‘Grand Nain’) under Simulated Water Deficit or Salinity Stress. S. Afr. J. Bot. 2020, 132, 155–163. [Google Scholar] [CrossRef]
- Baz, H.; Creech, M.; Chen, J.; Gong, H.; Bradford, K.; Huo, H. Water-Soluble Carbon Nanoparticles Improve Seed Germination and Post-Germination Growth of Lettuce under Salinity Stress. Agronomy 2020, 10, 1192. [Google Scholar] [CrossRef]
- Noohpisheh, Z.; Amiri, H.; Mohammadi, A.; Farhadi, S. Effect of the Foliar Application of Zinc Oxide Nanoparticles on Some Biochemical and Physiological Parameters of Trigonella foenum-graecum under Salinity Stress. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2021, 155, 267–280. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Chen, C.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P.; Yu, F. Zinc Oxide Nanoparticles (Zno-NPs) Induce Salt Tolerance by Improving the Antioxidant System and Photosynthetic Machinery in Tomato. Plant Physiol. Biochem. 2021, 161, 122–130. [Google Scholar] [CrossRef]
- Kumari, A.; Kaur, R.; Kaur, R. An Insight into Drought Stress and Signal Transduction of Abscisic Acid. Plant Sci. Today 2018, 5, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-K.; Li, L.-L.; Cao, Z.-H.; Zhao, Q.; Li, M.; Zhang, L.-Y.; Hao, Y.-J. Molecular Cloning and Functional Characterization of a Novel Apple Mdcipk6l Gene Reveals Its Involvement in Multiple Abiotic Stress Tolerance in Transgenic Plants. Plant Mol. Biol. 2012, 79, 123–135. [Google Scholar] [CrossRef]
- Chandra, P.; Wunnava, A.; Verma, P.; Chandra, A.; Sharma, R.K. Strategies to Mitigate the Adverse Effect of Drought Stress on Crop Plants—Influences of Soil Bacteria: A Review. Pedosphere 2021, 31, 496–509. [Google Scholar] [CrossRef]
- Sanjari, S.; Shobbar, Z.-S.; Ghanati, F.; Afshari-Behbahanizadeh, S.; Farajpour, M.; Jokar, M.; Khazaei, A.; Shahbazi, M. Molecular, Chemical, and Physiological Analyses of Sorghum Leaf Wax under Post-Flowering Drought Stress. Plant Physiol. Biochem. 2021, 159, 383–391. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Schmidhalter, U. Drought and Salinity: A Comparison of Their Effects on Mineral Nutrition of Plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Ashkavand, P.; Tabari, M.; Zarafshar, M.; Tomášková, I.; Struve, D. Effect of SiO2 Nanoparticles on Drought Resistance in Hawthorn Seedlings. For. Res. Pap. 2015, 76, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Aghdam, M.T.B.; Mohammadi, H.; Ghorbanpour, M. Effects of Nanoparticulate Anatase Titanium Dioxide on Physiological and Biochemical Performance of Linum usitatissimum (Linaceae) under Well-Watered and Drought Stress Conditions. Braz. J. Bot. 2016, 39, 139–146. [Google Scholar] [CrossRef]
- Kamalizadeh, M.; Bihamta, M.; Zarei, A. Drought Stress and TiO2 Nanoparticles Affect the Composition of Different Active Compounds in the Moldavian Dragonhead Plant. Acta Physiol. Plant 2019, 41, 21. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Andrews, J.; Fugice, J.; Singh, U.; Bindraban, P.S.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Facile Coating of Urea with Low-Dose ZnO Nanoparticles Promotes Wheat Performance and Enhances Zn Uptake Under Drought Stress. Front. Plant Sci. 2020, 11, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and Silica Nanostructure-Based Recovery of Strawberry Plants Subjected to Drought Stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Nguyen, D.V.; Nguyen, H.M.; Le, N.T.; Nguyen, K.H.; Nguyen, H.T.; Le, H.M.; Nguyen, A.T.; Dinh, N.T.T.; Hoang, S.A.; Ha, C.V. Copper Nanoparticle Application Enhances Plant Growth and Grain Yield in Maize Under Drought Stress Conditions. J. Plant Growth Regul. 2021, 1–12. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.F.; Abd El-Mageed, T.A.; Abd El-Mageed, S.A.; Rady, M.M.; Ali, E.F. Foliar Application of Zinc Oxide Nanoparticles Promotes Drought Stress Tolerance in Eggplant (Solanum melongena L.). Plants 2021, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Behboudi, F.; Tahmasebi-Sarvestani, Z.; Kassaee, M.Z.; Modarres-Sanavy, S.A.M.; Sorooshzadeh, A.; Mokhtassi-Bidgoli, A. Evaluation of Chitosan Nanoparticles Effects with Two Application Methods on Wheat under Drought Stress. J. Plant Nutr. 2019, 42, 1439–1451. [Google Scholar] [CrossRef]
- Elsheery, N.I.; Helaly, M.N.; El-Hoseiny, H.M.; Alam-Eldein, S.M. Zinc Oxide and Silicone Nanoparticles to Improve the Resistance Mechanism and Annual Productivity of Salt-Stressed Mango Trees. Agronomy 2020, 10, 558. [Google Scholar] [CrossRef]
- Adrees, M.; Khan, Z.S.; Hafeez, M.; Rizwan, M.; Hussain, K.; Asrar, M.; Alyemeni, M.N.; Wijaya, L.; Ali, S. Foliar Exposure of Zinc Oxide Nanoparticles Improved the Growth of Wheat (Triticum aestivum L.) and Decreased Cadmium Concentration in Grains under Simultaneous Cd and Water Deficient Stress. Ecotoxicol. Environ. Saf. 2021, 208, 111627. [Google Scholar] [CrossRef]
- Mosa, K.A.; El-Naggar, M.; Ramamoorthy, K.; Alawadhi, H.; Elnaggar, A.; Wartanian, S.; Ibrahim, E.; Hani, H. Copper Nanoparticles Induced Genotoxicty, Oxidative Stress, and Changes in Superoxide Dismutase (SOD) Gene Expression in Cucumber (Cucumis sativus) Plants. Front. Plant Sci. 2018, 9, 872. [Google Scholar] [CrossRef] [Green Version]
- Burman, U.; Saini, M.; Kumar, P.-. Effect of Zinc Oxide Nanoparticles on Growth and Antioxidant System of Chickpea Seedlings. Toxicol. Environ. Chem. 2013, 95, 605–612. [Google Scholar] [CrossRef]
- Singh, N.B.; Amist, N.; Yadav, K.; Singh, D.; Pandey, J.K.; Singh, S.C. Zinc Oxide Nanoparticles as Fertilizer for the Germination, Growth and Metabolism of Vegetable Crops. J. Nanoeng. Nanomanuf. 2013, 3, 353–364. [Google Scholar] [CrossRef]
- Hernández-Hernández, H.; Quiterio-Gutiérrez, T.; Cadenas-Pliego, G.; Ortega-Ortiz, H.; Hernández-Fuentes, A.D.; Cabrera De La Fuente, M.C.; Valdés-Reyna, J.; Juárez-Maldonado, A. Impact of Selenium and Copper Nanoparticles on Yield, Antioxidant System, and Fruit Quality of Tomato Plants. Plants 2019, 8, 355. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, R.C.; Kumaraswamy, R.V.; Kumari, S.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-chitosan Nanoparticle Boost Defense Responses and Plant Growth in Maize (Zea mays L.). Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, P.; Priyanka, N.; Manikandan, K.; Ganeshbabu, I.; Indiraarulselvi, P.; Geetha, N.; Muralikrishna, K.; Bhattacharya, R.C.; Tiwari, M.; Sharma, N.; et al. Enhanced Plant Growth Promoting Role of Phycomolecules Coated Zinc Oxide Nanoparticles with P Supplementation in Cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 110, 118–127. [Google Scholar] [CrossRef]
- Priyanka, N.; Venkatachalam, P. Biofabricated Zinc Oxide Nanoparticles Coated with Phycomolecules as Novel Micronutrient Catalysts for Stimulating Plant Growth of Cotton. Adv. Nat. Sci. Nanosci. Nanotechnol. 2016, 7, 045018. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanpour, M.; Hatami, M.; Hatami, M. Activating Antioxidant Enzymes, Hyoscyamine and Scopolamine Biosynthesis of Hyoscyamus niger l. Plants with Nano-sized Titanium Dioxide and Bulk Application. Acta Agric. Slov. 2015, 105, 23–32. [Google Scholar] [CrossRef]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc Oxide Nanoparticle-Mediated Changes in Photosynthetic Efficiency and Antioxidant System of Tomato Plants. Photosynthesis 2018, 56, 678–686. [Google Scholar] [CrossRef]
- de Sousa, A.; Saleh, A.M.; Habeeb, T.H.; Hassan, Y.M.; Zrieq, R.; Wadaan, M.A.; Hozzein, W.N.; Selim, S.; Matos, M.; AbdElgawad, H. Silicon Dioxide Nanoparticles Ameliorate the Phytotoxic Hazards of Aluminum in Maize Grown on Acidic Soil. Sci. Total Environ. 2019, 693, 133636. [Google Scholar] [CrossRef] [PubMed]
- Moradbeygi, H.; Jamei, R.; Heidari, R.; Darvishzadeh, R. Investigating the Enzymatic and Non-Enzymatic Antioxidant Defense by Applying Iron Oxide Nanoparticles in Dracocephalum moldavica L. Plant under Salinity Stress. Sci. Hortic. 2020, 272, 109537. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. Changes in Soil Properties and Salt Tolerance of Safflower in Response to Biochar-Based Metal Oxide Nanocomposites of Magnesium and Manganese. Ecotoxicol. Environ. Saf. 2021, 211, 111904. [Google Scholar] [CrossRef]
- Rivero-Montejo, S.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as Novel Elicitors to Improve Bioactive Compounds in Plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Wang, J.; Mei, J.; Ren, G. Plant microRNAs: Biogenesis, Homeostasis, and Degradation. Front. Plant Sci. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazier, T.P.; Burklew, C.E.; Zhang, B. Titanium Dioxide Nanoparticles Affect the Growth and Microrna Expression of Tobacco (Nicotiana tabacum). Funct. Integr. Genom. 2014, 14, 75–83. [Google Scholar] [CrossRef]
- Kaveh, R.; Li, Y.-S.; Ranjbar, S.; Tehrani, R.; Brueck, C.; Van Aken, B. Changes in Arabidopsis thaliana Gene Expression in Response to Silver Nanoparticles and Silver Ions. Environ. Sci. Technol. 2013, 47, 10637–10644. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Fan, X.; Li, X.; Zhang, Z.; Sun, L.; Fu, Z.; Lavoie, M.; Pan, X.; Qian, H. Distinct Physiological and Molecular Responses in Arabidopsis thaliana Exposed to Aluminum Oxide Nanoparticles and Ionic Aluminum. Environ. Pollut. 2017, 228, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Sanzari, I.; Leone, A.; Ambrosone, A. Nanotechnology in Plant Science: To Make a Long Story Short. Front. Bioeng. Biotechnol. 2019, 7, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Hu, P.; Li, F.; Wu, H.; Shen, Y.; White, J.C.; Tian, X.; Li, Z.; Giraldo, J.P. Emerging Investigator Series: Molecular Mechanisms of Plant Salinity Stress Tolerance Improvement by Seed Priming with Cerium Oxide Nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Večeřová, K.; Večeřa, Z.; Dočekal, B.; Oravec, M.; Pompeiano, A.; Tříska, J.; Urban, O. Changes of Primary and Secondary Metabolites In Barley Plants Exposed to CdO Nanoparticles. Environ. Pollut. 2016, 218, 207–218. [Google Scholar] [CrossRef]
- Raigond, P.; Raigond, B.; Kaundal, B.; Singh, B.; Joshi, A.; Dutt, S. Effect of Zinc Nanoparticles on Antioxidative System of Potato Plants. J. Environ. Biol. 2017, 38, 435–439. [Google Scholar] [CrossRef]
- Karimi, N.; Behbahani, M.; Dini, G.; Razmjou, A. Enhancing the Secondary Metabolite and Anticancer Activity of Echinacea Purpurea Callus Extracts by Treatment with Biosynthesized ZnO Nanoparticles. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 045009. [Google Scholar] [CrossRef]
- Modarresi, M.; Chahardoli, A.; Karimi, N.; Chahardoli, S. Variations of Glaucine, Quercetin and Kaempferol Contents in Nigella arvensis Against Al2O3, NiO, and TiO2 Nanoparticles. Heliyon 2020, 6, e04265. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Song, F.; Guo, J.; Zhu, X.; Liu, S.; Liu, F.; Li, X. Nano-ZnO-Induced Drought Tolerance is Associated with Melatonin Synthesis and Metabolism in Maize. Int. J. Mol. Sci. 2020, 21, 782. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Hosseini, M.S.; Daneshvar Hakimi Meybodi, N.; Peijnenburg, W. Mitigation of the Effect of Drought on Growth and Yield of Pomegranates by Foliar Spraying of Different Sizes of Selenium Nanoparticles. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 1–18. [Google Scholar] [CrossRef]
- Ji, H.; Pardo, J.M.; Batelli, G.; Van Oosten, M.J.; Bressan, R.A.; Li, X. The Salt Overly Sensitive (SOS) Pathway: Established and Emerging Roles. Mol. Plant 2013, 6, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.; El-Sharkawy, I.; Sherif, S. Salt Stress Signals on Demand: Cellular Events in the Right Context. Int. J. Mol. Sci. 2020, 21, 3918. [Google Scholar] [CrossRef]
- Ishitani, M.; Liu, J.; Halfter, U.; Kim, C.-S.; Shi, W.; Zhu, J.-K. SOS3 Function in Plant Salt Tolerance Requires N-Myristoylation and Calcium Binding. Plant Cell 2000, 12, 1667–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Ballesta, M.C.; Zapata, L.; Chalbi, N.; Carvajal, M. Multiwalled carbon nanotubes enter broccoli cells enhancing growth and water uptake of plants exposed to salinity. J. Nanobiotechnol. 2016, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleck, A.T.; Schulze, S.; Hinrichs, M.; Specht, A.; Waßmann, F.; Schreiber, L.; Schenk, M.K. Silicon Promotes Exodermal Casparian Band Formation in Si-Accumulating and Si-Excluding Species by Forming Phenol Complexes. PLoS ONE 2015, 10, e0138555. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.; Flowers, T.J.; Gong, H. Silicon Decreases Chloride Transport in Rice (Oryza sativa L.) in Saline Conditions. J. Plant Physiol. 2013, 170, 847–853. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Gong, H.-J.; Yin, J.-L. Role of Silicon in Mediating Salt Tolerance in Plants: A Review. Plants 2019, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Dar, N.A.; Amin, I.; Wani, W.; Wani, S.A.; Shikari, A.B.; Wani, S.H.; Masoodi, K.Z. Abscisic Acid: A Key Regulator of Abiotic Stress Tolerance in Plants. Plant Gene 2017, 11, 106–111. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Hussain, H.I.; Yi, Z.; Rookes, J.E.; Kong, L.; Cahill, D.M. Delivery of Abscisic Acid to Plants Using Glutathione Responsive Mesoporous Silica Nanoparticles. J. Nanosci. Nanotechnol. 2018, 18, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Raza, M.A.; Awan, S.A.; Shah, G.A.; Rizwan, M.; Ali, B.; Tariq, R.; Hassan, M.J.; Alyemeni, M.N.; Brestic, M.; et al. Amelioration of Salt Induced Toxicity in Pearl Millet by Seed Priming with Silver Nanoparticles (AgNPs): The Oxidative Damage, Antioxidant Enzymes and Ions Uptake are Major Determinants of Salt Tolerant Capacity. Plant Physiol. Biochem. 2020, 156, 221–232. [Google Scholar] [CrossRef]
- Jahani, M.; Khavari-Nejad, R.A.; Mahmoodzadeh, H.; Saadatmand, S. Effects of Cobalt Oxide Nanoparticles (CO3O4 NPs) on Ion Leakage, Total Phenol, Antioxidant Enzymes Activities and Cobalt Accumulation in Brassica napus L. Not. Bot. Hortic. Agrobot. Cluj-Napoca 2020, 48, 1260–1275. [Google Scholar] [CrossRef]
- Yan, L.; Li, P.; Zhao, X.; Ji, R.; Zhao, L. Physiological and Metabolic Responses of Maize (Zea mays) Plants to Fe3O4 Nanoparticles. Sci. Total Environ. 2020, 718, 137400. [Google Scholar] [CrossRef]
- Sofy, A.; Sofy, M.; Hmed, A.; Dawoud, R.; Alnaggar, A.; Soliman, A.; El-Dougdoug, N. Ameliorating the Adverse Effects of Tomato mosaic tobamovirus Infecting Tomato Plants in Egypt by Boosting Immunity in Tomato Plants Using Zinc Oxide Nanoparticles. Molecules 2021, 26, 1337. [Google Scholar] [CrossRef] [PubMed]
- Chahardoli, A.; Karimi, N.; Ma, X.; Qalekhani, F. Effects of Engineered Aluminum and Nickel Oxide Nanoparticles on the Growth and Antioxidant Defense Systems of Nigella arvensis L. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pelegrino, M.T.; Pieretti, J.C.; Lange, C.N.; Kohatsu, M.Y.; Freire, B.M.; Batista, B.L.; Fincheira, P.; Tortella, G.R.; Rubilar, O.; Seabra, A.B.; et al. Foliar Spray Application of CuO Nanoparticles (NPs) and S -Nitrosoglutathione Enhances Productivity, Physiological and Biochemical Parameters of Lettuce Plants. J. Chem. Technol. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Sohail; Kamran, K.; Kemmerling, B.; Shutaywi, M.; Mashwani, Z.U.R. Nano Zinc Elicited Biochemical Characterization, Nutritional Assessment, Antioxidant Enzymes and Fatty Acid Profiling of Rapeseed. PLoS ONE 2020, 15, e0241568. [Google Scholar] [CrossRef]
- Pavani, K.V.; Beulah, M.; Poojitha, G.U.S.; Kantabathini, V.P.; Mallula, B.; Udayar, S.P.G. The Effect of Zinc Oxide Nanoparticles (ZnO NPs) on Vigna mungo L. Seedling Growth and Antioxidant Activity. Nanosci. Nanotechnol. Asia 2020, 10, 117–122. [Google Scholar] [CrossRef]
- Marchiol, L.; Iafisco, M.; Fellet, G.; Adamiano, A. Nanotechnology Support the Next Agricultural Revolution: Perspectives to Enhancement of Nutrient Use Efficiency. In Advances in Agronomy; Academic Press Sparks: Amsterdam, The Netherlands, 2020; Volume 161, pp. 27–116. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Nanoparticles | Plants | Germination and Morphological Responses | Physiological Responses | References |
|---|---|---|---|---|
| Si (10 μM) | Pisum sativum L. | Presence of Si NPs improved the growth in presence of Cr | Si NPs minimized the Cr storage, enhanced the synthesis of defense enzymes and augmented nutrient uptake | [69] |
| ZnO (25 mg/L) | Leucaena leucocephala | Application of NPs induced seedling growth | ZnO NPs amendment improved pigments and soluble proteins, reduced peroxidation; there was rise in the antioxidant defense enzymes | [70] |
| Fe3O4 | Triticum aestivum L. | Fe3O4 NP treatment minimized the inhibitory action of HMs | Fe3O4 NPs supplementation improved the level of superoxide dismutase and peroxidase | [71] |
| Si (19, 48, and 202 nm) | Oryza sativa L. | Si NPs enhanced the number of cultured cells and decreased proportionally with the rise in NP size; the treatment maintained the cellular integrity in the presence of metals | Si NPs amendment caused altered expression of genes responsible for reduced metal uptake | [72] |
| ZnO (0, 50, 75, and 100 mg/L) | Zea mays L. | Treatment caused rise in plant length, leaf number, and biomass | ZnO NPs application enhanced chlorophyll content, gas exchange characteristics, and antioxidant enzymes; addition led to reduced content of Cd in root and shoot | [73] |
| ZnO (0, 25, 50, 75, and 100 mg/L) and Fe NPs (0, 5, 10, 15, and 20 mg/L) | T. aestivum L. | Treatment induced plant growth, dry weight, and grains under Cd stress | Addition of NPs decreased the loss of electrolyte and activity of superoxide dismutase and peroxidase along with diminished Cd accumulation | [74] |
| Si | Glycine max L. | Si NPs minimized the growth inhibitory action of Hg | Incorporation of Si NPs improved the chlorophyll content and reduced the Hg content in root and shoot | [63] |
| Mel-Au (200 μM) | O. sativa L. | — | Application of Mel-Au NPs caused reduction of Cd level in root and shoot, improved chlorophyll content and raised the activity of antioxidant enzymes | [64] |
| Fe (25 and 50 mg/L) | O. sativa L. | Treatment of Fe NPs improved plant length and dry weight | Fe NPs application caused rise in the level of proline, glutathione and phyto-chelatins; Fe NPs addition led to improved defense enzymes and glyoxalase machinery | [19] |
| ZnO (10–100 mg/L) | O. sativa L. | Amendment of ZnO increased the growth of seedlings | Treatment facilitated reduced accumulation of arsenic in root and shoot together with rise in phytochelatin level | [75] |
| Cu (25, 50, and 100 mg kg−1 of soil) | T. aestivum L. | Rise in plant height and shoot dry weight | Increase in N and P content; reduced Cd transport, rise in the level of vital ions and antioxidant pool | [58] |
| Cu (0, 25, 50, and 100 mg kg−1 of soil) | T. aestivum L. | Improved biomass and growth | Reduced Cr availability; increase in nutrient uptake; rise in antioxidant content | [57] |
| Fe2O3 (0, 25, 50, and 100 mg kg−1 soil) | O. sativa L. | Improved fresh and dry biomass; increased height | Augmented detoxifying enzymes, photosynthetic potential, and nutrient uptake attributes; reduced formation of ROS, lowered expression of genes supporting the transport of Cd; restricted Cd mobilization in upper plant parts | [59] |
| Fe2O3 (25, 50, and 100 mg kg−1 soil) | T. aestivum L. | Rise in plant fresh and dry biomass; increase in plant length | Reduced Cd transport; enhanced N, P, and K content; increased antioxidants and pigment content | [60] |
| TiO2 (0, 100, and 250 mg/L soil) | Z. mays | Foliar application improved shoot and root dry weight | Reduced accumulation of Cd; increased activities of antioxidant enzymes | [61] |
| SiO2 (30 and 50 nm) | G. max | Improved seedling fresh weight | Improved chlorophyll content; lowered accumulation of Hg in root | [63] |
| Au (200 μM) | O. sativa L. | — | Reduced level of Cd in root and leaves by 33 and 46.2%, respectively; improvement in antioxidant defense enzyme; restricted expression of genes associated with metal transport | [64] |
| Si (0, 25, 50, and 100 mg/kg soil) | T. aestivum L. | Improved plant height | Improved chlorophyll; photosynthesis; diminished Cd content in tissues; | [64] |
| ZnO (0, 50, and 100 mg L−1) | G. max | Improved root and shoot growth | Reduced arsenic concentration in root and shoot; improved photosynthesis, water loss, photochemical yield; raised antioxidative defense enzymes | [66] |
| Ti (0.1 to 0.25%) | Vigna radiata L. | Augmented radicle length and biomass | Decline in the level of ROS and lipid peroxidation; upregulation of genes related with antioxidative enzymes | [67] |
| Se and Si (5, 10, and 20 mg L−1) | O. sativa L. | — | Lowered accrual of Cd and Pb; improved yield | [68] |
| Nanoparticles | Plants | Germination and Morphological Responses | Physiological Responses | References |
|---|---|---|---|---|
| Ag (0, 2, 5, and 10 mM) | Triticum aestivum L. | Seed priming with Ag NPs significantly augmented the fresh and dry biomass of salinity stressed wheat plants at all doses compared to the control. | Ag NPs increased the activities of vital antioxidative enzymes whilst declined the contents of stress indicators, i.e., MDA and H2O2 in wheat leaves as compared to salt stressed plants. | [85] |
| Zn-, B-, Si-, and Zeolite NPs | Solanum tuberosum L., Diamont cultivar | Application of individual and binary treatment of NPs improved plant height, shoot dry weight, number of stems per plant, and tuber yield as compared to the control. | NP treatment increased leaf relative water content, leaf photosynthetic rate, leaf stomatal conductance, and chlorophyll content in comparison to the control; improved nutrients contents, leaf proline content, and leaf gibberellic acid level; and enhanced the contents of protein adn carbohydrates, and antioxidative enzymes’ activities. | [87] |
| Fe (0, 0.08, and 0.8 ppm), and potassium silicate (0, 1, and 2 mM) | Vitis vinifera | — | Application of NPs significantly increased the total protein content, activities of antioxidative enzymes (POD, CAT, and SOD), and hydrogen peroxide, while reduced proline content. | [88] |
| Fe (0.0, 0.08, and 0.8 ppm) | Fragaria ananassa | Application of Fe NPs (at higher concentrations) increased root dry weight and dry weight of the explants. | Fe NPs improved the contents of photosynthetic pigments and total soluble carbohydrate, membrane stability index, and relative water content of salinity-stressed plants. | [89] |
| N–Na2SiO3 (400 ppm) | S. tuberosum L. | Foliar spraying of N–Na2SiO3 restored the tuber number per plant and tuber yield along with improved water use efficiency and tuber dry matter percentage under salinity stress. | Application of N–Na2SiO3 exerted positive impacts on the quantum yield of PS II, carotenoids content, and DPPH radical scavenging activity in salinity stressed plants. | [90] |
| SiO2 (0, 50, 100, and 150 mg/L) | Musa acuminata | All doses of SiO2 NPs improved the number of shoots and shoot length of banana. | Application of SiO2 NPs increased chlorophyll content, lowered electrolyte leakage, reduced MDA content, and altered the content of phenolic compounds | [91] |
| CNPs (0.3% and 90–110 nm) | Lactuca sativa | The salinity-induced deleterious effects on germination and associated parameters were alleviated by the exposure of C NPs, e.g., treatment of C NPs for 2 h significantly improved the germination rate in some varieties. | — | [92] |
| ZnO (0, 1000, and 3000 ppm) | Trigonella foenum-graecum | — | Interaction of NaCl and ZnO was recorded to reverse the salinity induced consequences (L-proline, protein, MDA, aldehydes, sugars, H2O2, and antioxidative enzymes) in both cultivars, but the results were more apparent in case cv. Ardestanian than cv. Mashhadian. | [93] |
| ZnO (10, 50, and 100 mg/L) | Lycopersicon esculentum | Foliar spraying of ZnO NPs increased shoot length and root length, biomass, and leaf area. | Increased chlorophyll content and photosynthetic attributes, protein content, and activities of antioxidative enzymes (POX, SOD, and CAT) in salinity-stressed tomato plants. | [94] |
| TiO2, (40, 60, and 80 ppm) | Zea mays L. | Seed priming with TiO2 positively impacted the germination (germination percentage, germination energy, and seedling vigor index) and seedling growth (lengths of root and shoot, fresh, and dry weight) and reduced the mean emergence time. | Results showed the enhancement in potassium ion concentration, relative water content, contents of total phenolic and proline contents; increased SOD, CAT, and PAL activities; and decreased sodium ion concentration, membrane electrolyte leakage, and MDA content. | [53] |
| Nanoparticles | Plants | Germination and Morphological Responses | Physiological Responses | References |
|---|---|---|---|---|
| Silica (0, 10, 50, and 100 mg/L) | S NPs increased plant biomass and xylem water potential in drought-stressed seedlings. | S NPs improved the photosynthetic rate and stomatal conductance considerably; no effects were recorded on MDA content, relative water content, and electrolyte leakage index; carbohydrate and proline content under drought stress declined. | [101] | |
| TiO2 (0, 10, 100, and 500 mg/L, and 10–25 nm) | Linum usitatissimum L. | Number of capsules per plant increased in plants under the application of TiO2 as compared to control and enhanced the seed weight. | Nano-TiO2 treatment of drought-stressed plants surged carotenoids content and ameliorated cell membrane damage, and seed oil and protein contents (at 100 mg/L) compared to normal. | [102] |
| Chitosan (0, 30, 60, and 90 ppm) | Triticum aestivum L. | Foliar spraying of C NPs (particularly at 90 ppm) at tillering, stem elongation, and heading stages caused increase in leaf area, crop yield, and biomass as compared to the control. | Application of C NPs increased relative water content, chlorophyll content, photosynthetic rate, and CAT and SOD activities in comparison to the control. | [108] |
| TiO2 (0, 5, 10, 20, 30, 50, 100, and 150 ppm) | Dracocephalum moldavica L. | TiO2 treatment had no significant impact on the plant dry weight | TiO2 application (30–50 ppm) increased certain beneficial phenolic substances (rosmarinic acid and chlorogenic acid) in stressed plants. | [103] |
| Si (0, 25, 50, and 100 mg/kg) | T. aestivum L. | Treatment of Si NPs to drought-stressed plant grown in Cd contaminated soil showed maximum values of shoot, root, and grain dry biomass, i.e., 70%, 54%, and 75%, respectively (at 100 mg/kg). | Application of Si NPs significantly improved the contents of chlorophyll and ameliorated oxidative stress by lowering the content of MDA, H2O2, and electrolyte leakage. | [65] |
| Fe (25, 50, and 100 mg/kg) | T. aestivum L. | Application of Fe NPs increased plant height, spike length, and dry weight under drought stress over control. | Chlorophyll a content increased up to 66% in wheat plants compared to the respective controls and eradicated the oxidative stress by switching antioxidative defense system. | [30] |
| ZnO (1.0%) | T. aestivum L. | Application of ZnO NPs strongly alleviated the delay in panicle initiation time and significantly increased grain yield as compared to the control under drought stress. | ZnO NPs enhanced Zn uptake but could not mitigate the negative impacts of drought stress on N and P uptake in wheat plants. | [104] |
| ZnO (50, 100, and 150 mg/L) and Si (150 and 300 mg/L) | Mangifera indica L. | Increased leaf area improved the total yield and fruit physiochemical characteristics as compared to the control under both NPs treatments. | Application of both NPs enhanced leaf NPK content; total carbohydrates, total sugars, and proline content; and SOD, POX, and CAT activities (at 100 mg/L nZnO and 150 mg/L nSi) over the control. | [109] |
| ZnO (0, 25, 50, and 100 mg/L) | T. aestivum L. | Application of ZnO NPs improved T. aestivum L. growth and crop yield. | Foliar spray enhanced chlorophyll content, and the activities of SOD and POXs. | [110] |
| SiO2 NPs, Se NPs, and SiO2/Se NPs (50 and 100 mg/L) | Fragaria × ananassa Duch. | Spraying of different NPs improved the growth and yield parameters of drought-stressed strawberry plants. | Treatment of Se/SiO2 at 100 mg/L showed maximal benefits to plants by preserving more photosynthetic pigments comparatively and increased relative water content, membrane stability index, and water use efficiency. Spraying of Se/SiO2 also increased drought tolerance via increasing the activities of CAT, APX, GPX, and SOD and decreased lipid peroxidation and H2O2 content. | [105] |
| Zero-valent copper NPs (3.333, 4.444, and 5.556 mg/L) | Zea mays | Cu NPs improved the biomass of drought-stressed plants, and increased total seed number and grain yield of maize plants. | Treatment of Cu NPs increased the contents of anthocyanin, chlorophyll, and carotenoid and improved drought stress tolerance by decreasing the oxidative stress via the enhancement of ROS scavenging antioxidative enzymes. | [106] |
| ZnO NPs (50 and 100 ppm) | Solanum melongena L. | Foliar spray of ZnO NPs improved the plant growth and productivity. | Application of ZnO NPs improved macro- and micronutrients’ uptake and increased relative water content in plants under drought stress. | [107] |
| Nanoparticles | Plant Name | Biochemical Changes/Significant Findings | References |
|---|---|---|---|
| Ag | Pennisetum glaucum | Seed priming with Ag NPs increased the activity of antioxidant enzymes, proline content, total phenolics and flavonoid contents in pearl millet. Ion homeostasis is maintained by decreasing the sodium (Na+) and Na+/K+ ratio, while potassium (K+) increased by NPs leading to increased salt tolerance. | [147] |
| Si | Coriandrum sativum | Si NPs are found to have a beneficial effect on coriander under Pb stress. Foliar application of Si NPs alleviated the adverse impacts of Pb by modulating the vitamin C, flavonoids, antioxidant enzyme activities (CAT, POD, and SOD), and malondialdehyde (MDA) and by minimizing the oxidative stress. | [18] |
| Co3O4 | Brassica napus | Low concentrations of Co3O4 NP treatment induced a beneficial effect on growth parameters by modulating APX, SOD, CAT, GR, phenylalanine ammonia lyase (PAL), tyrosine ammonia lyase (TAL), polyphenol oxidase (PPO), Guaiacol peroxidase (GPX), and glutathione S-transferase (GST) activity. | [148] |
| Fe3O4 | Zea mays | A beneficial role of NPs on root development and membrane integrity is reported. Root length of maize plant significantly increased after NP treatment, with decreased malondialdehyde (MDA) level. NPs was also found to inhibit the pathways related to antioxidant defense. | [149] |
| ZnO | Solanum lycopersicum | Application of ZnO NPs led to increased tolerance in tomato plant towards Tomato mosaic virus (ToMV). Treatment of NPs led to increased growth parameters; photosynthesis; increases in the activity of enzymatic antioxidant defense systems (CAT, SOD, POX, APX, GR, and lipoxygenase (LOX) activity); and the accumulation of phenolic compounds, ascorbic acid content, and proline production. | [150] |
| Al2O3 and NiO | Nigella arvensis | Activity of antioxidant enzymes such as APX, CAT, SOD, and POD are found to increase. Total antioxidant capacity, reducing power, iridoids content, saponin content, and phenolic content are also found to be higher in plants treated with Al2O3 NPs (100–2500 mg/L) and low concentrations (<100 mg/L) of NiO NP treatment. However, at higher treatment level (>100 mg/L) of NiO NPs, concentrations of these metabolites were reduced. | [151] |
| CuO and S-nitrosoglutathione (GSNO) | Lactuca sativa | Treatment resulted in positive effects on plant growth and threefold increases in lettuce dry weight. Total phenolic content (two-fold) and flavonoid content (four-fold) were also increased significantly. Increased accumulation of micronutrients such as K, Na, Ca, Mg, and S is observed in the leaves. | [152] |
| Zn | B. napus | Many parameters such as protein, proline, total soluble sugar (TSS), total flavonoid content (TFC), and total phenolic content (TPC) were changed after NP treatment. Activity of antioxidant enzymes such as SOD, POD, and CAT also increased. | [153] |
| ZnO | Vigna mungo | Treatment was found to induce the activities of antioxidant enzymes such as GPX, GR, and CAT and was increase in the ascorbic acid and hydrogen peroxide contents. | [154] |
| Combination of FeO and hydrogel | Oryza sativa | Treatment alleviated the Cd and drought stress in rice plants. Rice plants were found to have increased biomass, increased activity of antioxidant enzymes (SOD, POD, and CAT), and increased photosynthetic efficiency, together with a reduction in ROS. | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajput, V.D.; Minkina, T.; Kumari, A.; Harish; Singh, V.K.; Verma, K.K.; Mandzhieva, S.; Sushkova, S.; Srivastava, S.; Keswani, C. Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles. Plants 2021, 10, 1221. https://doi.org/10.3390/plants10061221
Rajput VD, Minkina T, Kumari A, Harish, Singh VK, Verma KK, Mandzhieva S, Sushkova S, Srivastava S, Keswani C. Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles. Plants. 2021; 10(6):1221. https://doi.org/10.3390/plants10061221
Chicago/Turabian StyleRajput, Vishnu D., Tatiana Minkina, Arpna Kumari, Harish, Vipin Kumar Singh, Krishan K. Verma, Saglara Mandzhieva, Svetlana Sushkova, Sudhakar Srivastava, and Chetan Keswani. 2021. "Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles" Plants 10, no. 6: 1221. https://doi.org/10.3390/plants10061221