
Physiological and Biochemical Responses of Ageratum conyzoides, Oryza sativa f. spontanea (Weedy Rice) and Cyperus iria to Parthenium hysterophorus Methanol Extract

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Effect of P. hysterophorus Extract on Chlorophyll-a Content of the Test Plant Species
2.2. Effect of P. hysterophorus Extract on Chlorophyll-b Content of the Test Plant Species
2.3. Effect of P. hysterophorus Extract on Total Chlorophyll Content of the Test Plant Species
2.4. Effect of P. hysterophorus Extract on Carotenoid Content of the Test Plant Species
2.5. Effect of P. hysterophorus Extract on Photosynthesis Rate of the Test Plant Species
2.6. Effect of P. hysterophorus Extract on Stomatal Conductance of the Test Plant Species
2.7. Effect of P. hysterophorus Extract on Transpiration Rate of the Test Plant Species
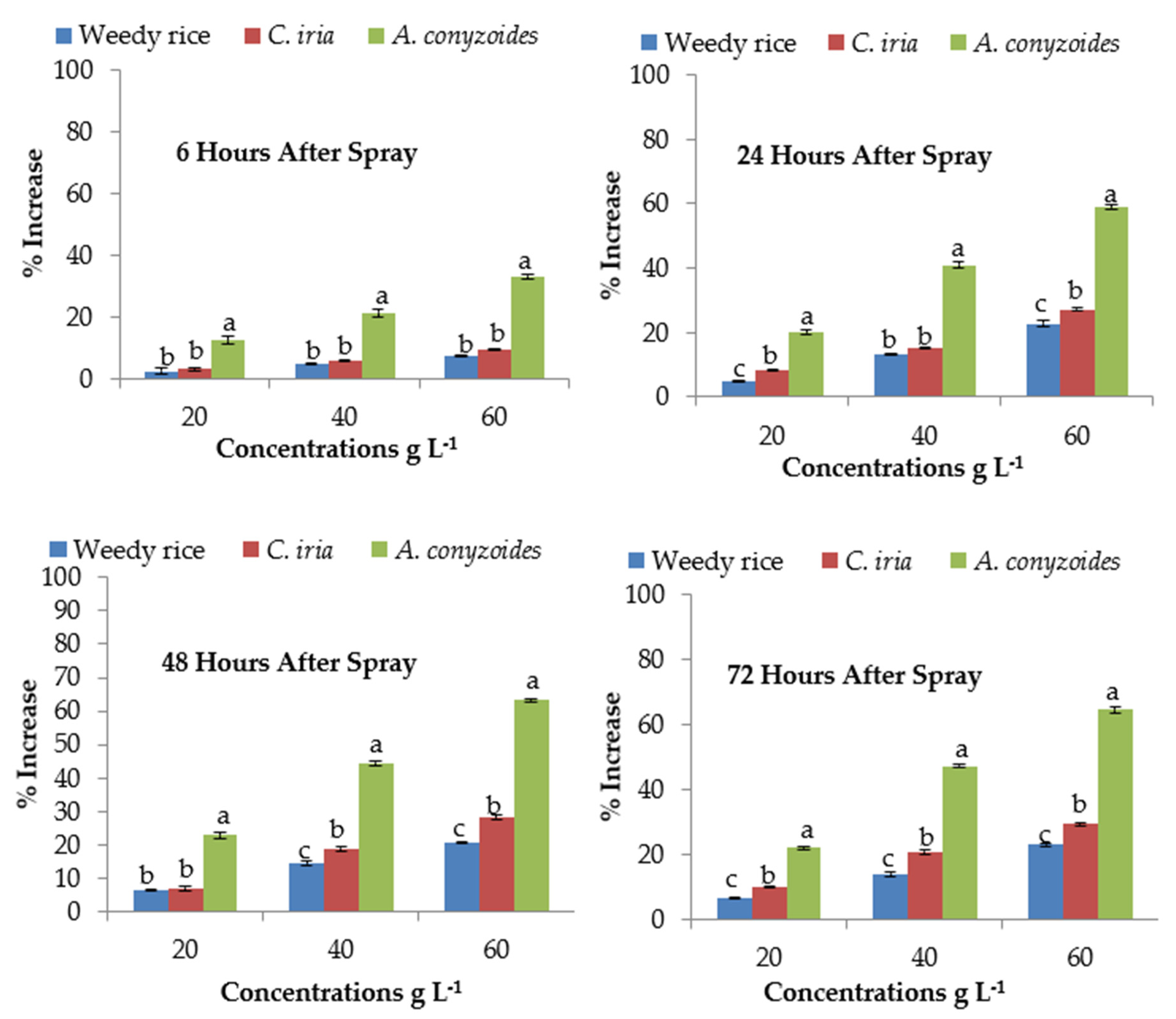
2.8. Effect of P. hysterophorus Extract on Malondialdehyde Content of the Test Plant Species
2.9. Effect of P. hysterophorus Extract on Proline Content of the Test Plant Species
2.10. Effect of P. hysterophorus Extract on Superoxide Dismutase of the Test Plant Species
2.11. Effect of P. hysterophorus Extract on Catalase Activity of the Test Plant Species
2.12. Effect of P. hysterophorus Extract on Peroxidase Activity of the Test Plant Species
3. Discussion
4. Materials and Methods
4.1. Experimental Site
4.2. Test Plants
4.3. Preparation of P. hysterophorus Extract for Foliar Spray
4.4. Experimental Layout
4.5. Data Collections
4.5.1. Photosynthesis Rate, Transpiration and Stomatal Conductance
4.5.2. Chlorophyll Pigments
4.5.3. Malondialdehyde (MDA) Content
4.5.4. Proline Content
4.5.5. Enzymes Extraction
Determination of SOD
Determination of CAT
Determination of POD
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Batish, D.R.; Kaur, S.; Singh, H.P.; Kohli, R.K. Nature of interference potential of leaf debris of Ageratum conyzoides. Plant Growth Regul. 2008, 57, 137–144. [Google Scholar] [CrossRef]
- Song, B.-K.; Chuah, T.-S.; Tam, S.M.; Olsen, K.M. Malaysian weedy rice shows its true stripes: WildOryzaand elite rice cultivars shape agricultural weed evolution in Southeast Asia. Mol. Ecol. 2014, 23, 5003–5017. [Google Scholar] [CrossRef] [PubMed]
- Van Chin, D. Biology and management of barnyardgrass, red sprangletop and weedy rice. Weed Biol. Manag. 2001, 1, 37–41. [Google Scholar] [CrossRef]
- Chauhan, B.S. Strategies to manage weedy rice in Asia. Crop. Prot. 2013, 48, 51–56. [Google Scholar] [CrossRef]
- Nadir, S.; Xiong, H.-B.; Zhu, Q.; Zhang, X.-L.; Xu, H.-Y.; Li, J.; Dongchen, W.; Henry, D.; Guo, X.-Q.; Khan, S.; et al. Weedy rice in sustainable rice production. A review. Agron. Sustain. Dev. 2017, 37, 46. [Google Scholar] [CrossRef]
- Ismail, B.S.; Siddique, M.A.B. The Inhibitory Effect of Grasshopper’s Cyperus (Cyperus iria L.) on the Seedling Growth of Five Malaysian Rice Varieties. Trop. Life Sci. Res. 2011, 22, 81–89. [Google Scholar]
- Srinivas, C.; Lakshmi, C. Parthenium: A wide angle view. Indian J. Dermatol. Venereol. Leprol. 2007, 73, 296–306. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef] [PubMed]
- Adkins, S.W.; Sowerby, M.S. Allelopathic potential of the weed, Parthenium hysterophorus L. in Australia. Plant Prot. Q. 1996, 11, 20–23. [Google Scholar]
- Motmainna, M.; Juraimi, A.S.; Uddin, M.K.; Asib, N.B.; Islam, A.K.M.M.; Hasan, M. Allelopathic potential of Malaysian invasive weed species on Weedy rice (Oryza sativa f. spontanea Roshev). Allelopathy J. 2021, 53, 53–68. [Google Scholar] [CrossRef]
- Kaya, A.; Yigit, E. The physiological and biochemical effects of salicylic acid on sunflowers (Helianthus annuus) exposed to flurochloridone. Ecotoxicol. Environ. Saf. 2014, 106, 232–238. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.H.; Houborg, R.; McCabe, M.F. Response of Chlorophyll, Carotenoid and SPAD-502 Measurement to Salinity and Nutrient Stress in Wheat (Triticum aestivum L.). Agronomy 2017, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.-F.; Teixeira, M.; Valentine, J.S. Superoxide Dismutases and Superoxide Reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef]
- Li, D.-H.; Yuan, Y. Hydrogen peroxide enhances antioxidative defense in the leaves of water caltrop (Trapa bicornis) seedlings treated with lead. Biologia 2016, 71, 100–108. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondial-dehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Long. 2014, 1–31. [Google Scholar] [CrossRef]
- Catalá, A.; Díaz, M. Editorial: Impact of Lipid Peroxidation on the Physiology and Pathophysiology of Cell Membranes. Front. Physiol. 2016, 7, 423. [Google Scholar] [CrossRef] [Green Version]
- Khayatnezhad, M.; Gholamin, R.; Jamaati-e-Somarin, S.; Zabihi-e-Mahmoodabad, R. The leaf chlorophyll content and stress resistance relationship considering in corn cultivars (Zea mays). Adv. Environ. Biol. 2011, 5, 118–122. [Google Scholar]
- Ding, H.; Cheng, Z.; Liu, M.; Hayat, S.; Feng, H. Garlic exerts allelopathic effects on pepper physiology in a hydroponic co-culture system. Biol. Open 2016, 5, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Yılmaz, E.; Tuna, A.L.; Bürün, B. Tolerance strategies developed by plants to the effects of salt stress. CBU J. Sci. 2011, 7, 47–66. [Google Scholar]
- Erdal, S. Androsterone-induced molecular and physiological changes in maize seedlings in response to chilling stress. Plant Physiol. Biochem. 2012, 57, 1–7. [Google Scholar] [CrossRef]
- Mishra, S.; Srivastava, S.; Tripathi, R.; Govindarajan, R.; Kuriakose, S.; Prasad, M. Phytochelatin synthesis and response of antioxidants during cadmium stress in Bacopa monnieri L. Plant Physiol. Biochem. 2006, 44, 25–37. [Google Scholar] [CrossRef]
- El-Shora, H.M.; Abd El-Gawad, A.M. Physiological and biochemical responses of Cucurbita pepo L. mediated by Portulaca oleracea L. allelopathy. Fresenius Environ. Bull. 2015, 24, 386–393. [Google Scholar]
- Lawlor, D.W. Limitation to Photosynthesis in Water-stressed Leaves: Stomata vs. Metabolism and the Role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding Oxidative Stress and Antioxidant Functions to Enhance Photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casson, S.; Gray, J.E. Influence of environmental factors on stomatal development. New Phytol. 2008, 178, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Blum, U.; Gerig, T.M. Relationships between Phenolic Acid Concentrations, Transpiration, Water Utilization, Leaf Area Expansion, and Uptake of Phenolic Acids: Nutrient Culture Studies. J. Chem. Ecol. 2005, 31, 1907–1932. [Google Scholar] [CrossRef]
- Li, Z.-H.; Wang, Q.; Ruan, X.; Pan, C.-D.; Jiang, D.-A. Phenolics and Plant Allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [Green Version]
- Rakhra, G.; Sharma, A.D.; Singh, J. Anti-oxidative potential of boiling soluble antioxidant enzymes inAmelioration of drought-induced oxidative stress in tolerant and sensitive cultivars of Triticum aestivum. J. Crop. Sci. Biotechnol. 2015, 18, 103–122. [Google Scholar] [CrossRef]
- Quan, L.-J.; Zhang, B.; Shi, W.-W.; Li, H.-Y. Hydrogen Peroxide in Plants: A Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Saidi, I.; Ayouni, M.; Dhieb, A.; Chtourou, Y.; Chaïbi, W.; Djebali, W. Oxidative damages induced by short-term exposure to cadmium in bean plants: Protective role of salicylic acid. S. Afr. J. Bot. 2013, 85, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.; Panda, S.K. Role of salicylic acid in regulating cadmium induced oxidative stress in Oryza sativa L. roots. Bulg. J. Plant Physiol. 2004, 30, 95–110. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, A.; Hayat, S.; Ahmad, A.; Tahir, I. Efficacy of brassinosteroid analogues in the mitigation of toxic effects of salt stress in Brassica juncea plants. J. Environ. Biol. 2017, 38, 27–36. [Google Scholar] [CrossRef]
- Abbas, W.; Ashraf, M.; Akram, N.A. Alleviation of salt-induced adverse effects in eggplant (Solanum melongena L.) by glycinebetaine and sugarbeet extracts. Sci. Hortic. 2010, 125, 188–195. [Google Scholar] [CrossRef]
- El-Khatib, A.A.; Barakat, N.A.; Nazeir, H. Growth And Physiological Response Of Some Cultivated Species under Allelopathic Stress of Calotropis Procera (Aiton) WT. Appl. Sci. Rep. 2016, 14, 237–246. [Google Scholar] [CrossRef]
- Motmainna, M.; Juraimi, A.S.; Uddin, M.K.; Asib, N.B.; Islam, A.K.M.M.; Hasan, M. Bioherbicidal Properties of Parthenium hysterophorus, Cleome rutidosperma and Borreria alata Extracts on Selected Crop and Weed Species. Agronomy 2021, 11, 643. [Google Scholar] [CrossRef]
- El-Mergawi, R.A.; Al-Humaid, A.I. Searching for natural herbicides in methanol extracts of eight plant species. Bull. Natl. Res. Cent. 2019, 43, 1–6. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Amin, T.M. Response to Salt Stress and Strategies to Improve Salt Tolerance in Chinese Kale (Brassica Oleracea var. Albo-Glabra). Ph.D. Thesis, University Putra Malaysia, Seri Kembangan, Malaysia, 2012. [Google Scholar]
- Stewart, R.R.C.; Bewley, J.D. Lipid Peroxidation Associated with Accelerated Aging of Soybean Axes. Plant Physiol. 1980, 65, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Gupta, A.S.; Heinen, J.L.; Holaday, A.S.; Burke, J.J.; Allen, R.D. Increased resistance to oxidative stress in transgenic plants that overexpress chloroplastic Cu/Zn superoxide dismutase. Proc. Natl. Acad. Sci. USA 1993, 90, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B-and ozone-induced biochemical changes in antioxidant enzymes of Ara-bidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Plants | Dose (g L−1) | Chlorophyll-a (mg g−1 FW) | Chlorophyll-b (mg g−1 FW) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Hours after Spray | Hours after Spray | ||||||||
| 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | ||
| 0 | 3.03a ± 0.04 (0) | 3.05a ± 0.06 (0) | 2.91a ± 0.11 (0) | 3.06a ± 0.07 (0) | 1.14a ± 0.08 (0) | 1.16a ± 0.09 (0) | 1.11a ± 0.12 (0) | 1.31a ± 0.13 (0) | |
| 20 | 2.65a ± 0.11 (12.50) | 2.13b ± 0.07 (30.01) | 2.04b ± 0.04 (29.78) | 2.11b ± 0.10 (31.10) | 1.01a ± 0.03 (13.53) | 0.86a ± 0.22 (25.55) | 0.82a ± 0.07 (26.54) | 0.96a ± 0.07 (26.45) | |
| A. conyzoides | 40 | 2.42a ± 0.03 (20.20) | 1.63b ± 0.04 (46.49) | 1.57b ± 0.03 (45.90) | 1.64b ± 0.10 (46.54) | 0.95a ± 0.07 (18.29) | 0.67b ± 0.14 (42.20) | 0.63b ± 0.07 (43.27) | 0.74b ± 0.06 (43.27) |
| 60 | 2.06a ± 0.07 (31.98) | 0.78b ± 0.10 (74.32) | 0.77b ± 0.03 (73.53) | 0.79b ± 0.04 (74.09) | 0.82a ± 0.13 (29.56) | 0.38b ± 0.03 (67.50) | 0.34b ± 0.05 (68.95) | 0.39b ± 0.06 (70.44) | |
| 0 | 3.89a ± 0.08 (0) | 3.86a ± 0.10 (0) | 3.90a ± 0.16 (0) | 3.95a ± 0.12 (0) | 2.37a ± 0.17 (0) | 2.23a ± 0.17 (0) | 2.12a ± 0.22 (0) | 2.15a ± 0.20 (0) | |
| 20 | 3.80a ± 0.08 (2.33) | 3.45b ± 0.07 (10.61) | 3.37c ± 0.10 (13.59) | 3.18c ± 0.03 (19.34) | 2.32a ± 0.02 (2.01) | 2.01b ± 0.08 (9.74) | 1.94b ± 0.20 (8.30) | 1.92b ± 0.13 (10.75) | |
| Weedy rice | 40 | 3.59a ± 0.04 (7.70) | 3.06b ± 0.10 (20.86) | 3.11b ± 0.09 (20.37) | 3.10b ± 0.10 (21.41) | 2.20a ± 0.16 (6.97) | 1.85b ± 0.09 (16.87) | 1.75b ± 0.11 (17.51) | 1.75b ± 0.11 (17.93) |
| 60 | 3.39a ± 0.03 (12.80) | 2.56b ± 0.06 (33.75) | 2.59b ± 0.05 (33.73) | 2.52b ± 0.05 (36.21) | 2.11a ± 0.09 (10.96) | 1.51b ± 0.06 (32.08) | 1.43b ± 0.09 (32.23) | 1.47b ± 0.07 (31.68) | |
| 0 | 3.90ab ± 0.07 (0) | 3.78b ± 0.05 (0) | 3.86ab ± 0.09 (0) | 3.99a ± 0.05 (0) | 3.39a ± 0.09 (0) | 3.07bc ± 0.12 (0) | 3.29ab ± 0.10 (0) | 2.83c ± 0.13 (0) | |
| 20 | 3.75a ± 0.04 (3.63) | 3.05b ± 0.11 (19.30) | 3.10b ± 0.07 (19.65) | 3.19b ± 0.07 (20.12) | 3.28a ± 0.06 (3.36) | 2.53b ± 0.29 (17.60) | 2.68b ± 0.28 (18.53) | 2.30b ± 0.06 (18.71) | |
| C. iria | 40 | 3.54a ± 0.06 (9.02) | 2.79b ± 0.07 (26.09) | 2.85b ± 0.07 (26.05) | 2.84b ± 0.05 (28.76) | 3.13a ± 0.06 (7.68) | 2.39b ± 0.07 (22.24) | 2.51bc ± 0.16 (23.75) | 2.15c ± 0.10 (22.61) |
| 60 | 3.27a ± 0.05 (16.15) | 2.42c ± 0.03 (36.03) | 2.45bc ± 0.06 (36.35) | 2.54b ± 0.06 (36.31) | 2.99a ± 0.01 (11.72) | 2.00c ± 0.07 (34.88) | 2.24b ± 0.06 (31.86) | 1.93c ± 0.05 (32.00) | |
| Test Plants | Dose (g L−1) | Total Chlorophyll (mg g−1 FW) | Carotenoids (mg g−1 FW) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Hours after Spray | Hours after Spray | ||||||||
| 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | ||
| 0 | 4.17ab ± 0.05 (0) | 4.20ab ± 0.04 (0) | 4.02b ± 0.09 (0) | 4.37a ± 0.08 (0) | 1.02b ± 0.03 (0) | 1.03a ± 0.03 (0) | 1.01a ± 0.02 (0) | 1.14a ± 0.03 (0) | |
| 20 | 3.66a ± 0.15 (12.79) | 2.99b ± 0.18 (22.78) | 2.86b ± 0.08 (28.88) | 3.07b ± 0.08 (29.70) | 0.86a ± 0.03 (16.82) | 0.70c ± 0.02 (31.94) | 0.68c ± 0.03 (32.30) | 0.77b ± 0.01 (32.08) | |
| A. conyzoides | 40 | 3.37a ± 0.10 (19.67) | 2.30b ± 0.11 (45.31) | 2.20b ± 0.07 (45.17) | 2.38b ± 0.10 (45.56) | 0.81a ± 0.02 (21.01) | 0.55c ± 0.01 (46.67) | 0.54c ± 0.01 (46.71) | 0.60b ± 0.01 (47.04) |
| 60 | 2.88a ± 0.16 (31.31) | 1.16b ± 0.11 (72.45) | 1.11b ± 0.07 (72.26) | 1.18b ± 0.06 (73.00) | 0.69a ± 0.03 (32.89) | 0.30b ± 0.01 (70.94) | 0.30b ± 0.01 (70.46) | 0.33b ± 0.02 (71.29) | |
| 0 | 6.26a ± 0.10 (0) | 6.09a ± 0.10 (0) | 6.02a ± 0.18 (0) | 6.10a ± 0.10 (0) | 1.84a ± 0.03 (0) | 1.47a ± 0.70 (0) | 1.78a ± 0.07 (0) | 1.80a ± 0.06 (0) | |
| 20 | 6.12a ± 0.07 (2.21) | 5.46b ± 0.06 (10.29) | 5.31bc ± 0.12 (11.73) | 5.10c ± 0.13 (16.32) | 1.79a ± 0.06 (2.65) | 1.30c ± 0.02 (11.67) | 1.53b ± 0.03 (13.64) | 1.49b ± 0.07 (17.05) | |
| Weedy rice | 40 | 5.79a ± 0.14 (7.43) | 4.91b ± 0.05 (19.40) | 4.85b ± 0.15 (19.36) | 4.87b ± 0.05 (20.18) | 1.71a ± 0.02 (7.13) | 1.15c ± 0.03 (21.89) | 1.40b ± 0.09 (21.22) | 1.40b ± 0.01 (22.16) |
| 60 | 5.50a ± 0.09 (12.10) | 4.07b ± 0.05 (33.14) | 4.02b ± 0.04 (32.20) | 3.99b ± 0.10 (34.62) | 1.64a ± 0.02 (10.82) | 1.03c ± 0.01 (30.08) | 1.24b ± 0.04 (30.01) | 1.21b ± 0.02 (32.45) | |
| 0 | 7.29a ± 0.06 (0) | 6.85b ± 0.10 (0) | 7.15a ± 0.04 (0) | 6.82b ± 0.12 (0) | 1.68b ± 0.03 (0) | 1.73b ± 0.03 (0) | 1.81a ± 0.03 (0) | 1.80a ± 0.04 (0) | |
| 20 | 7.03a ± 0.07 (3.51) | 5.58b ± 0.24 (18.54) | 5.78b ± 0.22 (19.14) | 5.49b ± 0.11 (19.53) | 1.61a ± 0.03 (3.84) | 1.48a ± 0.06 (14.33) | 1.48a ± 0.10 (18.43) | 1.48a ± 0.06 (18.06) | |
| C. iria | 40 | 6.67a ± 0.1 (8.40) | 5.18bc ± 0.06 (24.37) | 5.36b ± 0.11 (24.99) | 5.00c ± 0.14 (26.21) | 1.54a ± 0.02 (8.05) | 1.26b ± 0.02 (27.36) | 1.32b ± 0.03 (27.43) | 1.29b ± 0.15 (28.32) |
| 60 | 6.26a ± 0.03 (14.09) | 4.42c ± 0.06 (35.51) | 4.70b ± 0.05 (34.28) | 4.47c ± 0.06 (34.52) | 1.49a ± 0.01 (11.41) | 1.14b ± 0.03 (34.19) | 1.20b ± 0.03 (34.07) | 1.17b ± 0.04 (35.32) | |
| Test Plants | Dose (g L −1) | Photosynthesis Rate (µmol m−2 s−1) | Stomatal Conductance (mol m−2 s−1) | Transpiration Rate (mmol m−2 s−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hours after Spray | Hours after Spray | Hours after Spray | |||||||||||
| 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | ||
| 0 | 16.06a ± 0.06 (0) | 14.34c ± 0.03 (0) | 13.70d ± 0.01 (0) | 14.60b ± 0.05 (0) | 0.41a ± 0.01 (0) | 0.35c ± 0.02 (0) | 0.39b ± 0.01 (0) | 0.40b ± 0.03 (0) | 7.22a ± 0.04 (0) | 6.00b ± 0.01 (0) | 5.03c ± 0.01 (0) | 5.00c ± 0.06 (0) | |
| 20 | 15.00a ± 0.39 (6.56) | 10.35b ± 1.25 (27.86) | 10.15b ± 0.03 (25.89) | 10.40b ± 0.32 (28.79) | 0.39a ± 0.01 (4.40) | 0.29b ± 0.06 (17.94) | 0.30b ± 0.02 (23.61) | 0.30b ± 0.05 (23.59) | 6.80a ± 0.11 (5.84) | 4.19b ± 0.33 (30.20) | 3.80b ± 0.12 (24.33) | 3.73b ± 0.23 (25.28) | |
| A. conyzoides | 40 | 14.32a ± 0.44 (10.83) | 8.10b ± 0.63 (43.54) | 6.76c ± 0.01 (60.61) | 7.29bc ± 0.14 (50.09) | 0.38a ± 0.03 (7.50) | 0.21b ± 0.01 (39.15) | 0.20b ± 0.05 (48.42) | 0.21b ± 0.02 (47.54) | 6.55a ± 0.01 (9.27) | 2.75b ± 0.54 (54.08) | 2.26b ± 0.11 (55.00) | 2.24b ± 0.17 (55.09) |
| 60 | 12.57a ± 0.06 (21.73) | 3.93b ± 0.19 (72.61) | 3.34c ± 0.44 (75.62) | 3.09c ± 0.12 (78.85) | 0.32a ± 0.02 (20.38) | 0.14bc ± 0.01 (59.54) | 0.15b ± 0.07 (62.36) | 0.12c ± 0.05 (69.37) | 5.73a ± 0.14 (20.66) | 1.27b ± 0.07 (78.81) | 1.09bc ± 0.06 (78.22) | 1.05c ± 0.01 (78.94) | |
| 0 | 48.71a ± 0.29 (0) | 45.53b ± 0.29 (0) | 45.95b ± 0.68 (0) | 45.45b ± 0.04 (0) | 0.53a ± 0.01 (0) | 0.58ab ± 0.02 (0) | 0.57b ± 0.03 (0) | 0.61c ± 0.04 (0) | 14.80a ± 0.02 (0) | 13.47bc ± 0.01 (0) | 13.17c ± 0.05 (0) | 14.08b ± 0.58 (0) | |
| 20 | 47.88a ± 0.12 (1.70) | 43.23b ± 0.52 (5.04) | 43.33b ± 0.42 (5.70) | 42.67b ± 0.06 (6.11) | 0.52b ± 0.01 (1.57) | 0.56b ± 0.02 (4.72) | 0.54ab ± 1 (4.92) | 0.57a ± 0.02 (6.36) | 14.65a ± 0.16 (1.02) | 12.35b ± 0.02 (8.37) | 12.10b ± 0.10 (8.14) | 12.78b ± 0.91 (9.22) | |
| Weedy rice | 40 | 46.78a ± 0.20 (3.93) | 40.95b ± 0.13 (10.05) | 39.07c ± 0.11 (14.96) | 38.10d ± 0.05 (16.16) | 0.51b ± 0.01 (3.29) | 0.54a ± 0.01 (7.03) | 0.51b ± 0.02 (9.97) | 0.55a ± 0.05 (9.03) | 14.23a ± 0.01 (3.84) | 11.36b ± 0.02 (15.79) | 11.20b ± 0.09 (15.00) | 11.42b ± 1.26 (18.90) |
| 60 | 44.89a ± 0.23 (7.82) | 32.84b ± 0.10 (27.87) | 33.08b ± 0.69 (28.00) | 32.56b ± 0.50 (28.37) | 0.49a ± 0.04 (6.96) | 0.45b ± 0.01 (22.20) | 0.44b ± 0.05 (23.28) | 0.50a ± 0.01 (18.07) | 13.50a ± 0.50 (6.96) | 9.63b ± 0.48 (28.53) | 19.14b ± 0.25 (30.63) | 9.67b ± 0.59 (31.32) | |
| 0 | 46.84a ± 0.54 (0) | 42.35b ± 0.10 (0) | 42.23b ± 1.40 (0) | 43.14b ± 0.08 (0) | 0.41a ± 0.04 (0) | 0.43a ± 0.04 (0) | 0.38a ± 0.01 (0) | 0.40a ± 0.02 (0) | 13.09a ± 0.14 (0) | 11.22b ± 0.07 (0) | 11.22b ± 0.07 (0) | 11.29b ± 0.01 (0) | |
| 20 | 45.87a ± 0.10 (2.05) | 38.72b ± 0.34 (8.57) | 37.47c ± 0.30 (11.27) | 38.39b ± 0.56 (11.01) | 0.40a ± 0.03 (1.77) | 0.41a ± 0.05 (4.87) | 0.36a ± 0.02 (6.21) | 0.37a ± 0.01 (8.51) | 12.78a ± 0.14 (2.36) | 10.01b ± 0.74 (10.74) | 9.81b ± 0.62 (12.58) | 9.74b ± 0.62 (13.73) | |
| C. iria | 40 | 44.24a ± 0.45 (5.54) | 36.25b ± 0.70 (14.41) | 34.75c ± 0.19 (17.72) | 35.79b ± 0.44 (17.04) | 0.39a ± 0.04 (3.50) | 0.39a ± 0.01 (9.58) | 0.33a ± 0.02 (13.38) | 0.34a ± 0.03 (15.00) | 12.23a ± 0.88 (96.55) | 9.21b ± 0.50 (17.87) | 8.96b ± 0.46 (20.08) | 8.99b ± 1.10 (20.37) |
| 60 | 41.90a ± 0.54 (10.54) | 28.23b ± 1.12 (33.33) | 26.90b ± 0.10 (36.31) | 27.47b ± 0.40 (36.33) | 0.38a ± 0.10 (6.34) | 0.32a ± 0.02 (25.45) | 0.28a ± 0.03 (27.70) | 0.29a ± 0.01 (28.94) | 11.62A ± 0.01 (11.25) | 7.32b ± 0.22 (34.69) | 7.04b ± 0.03 (37.18) | 6.95b ± 0.48 (38.48) | |
| Test Plants | Dose (g L −1) | Malondialdehyde Content (µmol g−1 FW) | Proline Content (µmol g−1 FW) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Hours after Spray | Hours after Spray | ||||||||
| 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | ||
| 0 | 1.90a ± 0.06 | 1.79a ± 0.14 | 1.71a ± 0.03 | 1.79a ± 0.14 | 5.06a ± 0.28 | 4.93a ± 0.14 | 5.01a ± 0.15 | 4.99a ± 0.08 | |
| 20 | 2.32a ± 0.09 | 2.58a ± 0.18 | 2.45a ± 0.06 | 2.62a ± 0.21 | 6.60b ± 0.23 | 7.03ab ± 0.14 | 7.40a ± 0.32 | 7.12a ± 0.10 | |
| A. conyzoides | 40 | 2.71b ± 0.08 | 3.21a ± 0.22 | 3.14a ± 0.03 | 3.29a ± 0.24 | 7.83b ± 0.53 | 9.12a ± 0.29 | 9.41a ± 0.22 | 9.37a ± 0.06 |
| 60 | 3.49b ± 0.06 | 4.52a ± 0.29 | 4.37a ± 0.05 | 4.67a ± 0.33 | 9.57b ± 0.69 | 12.14a ± 0.42 | 12.51a ± 0.46 | 12.58a ± 0.08 | |
| 0 | 2.80a ± 0.11 | 2.70ab ± 0.06 | 2.58b ± 0.09 | 2.56b ± 0.11 | 4.13a ± 0.24 | 3.75b ± 0.20 | 4.01ab ± 0.07 | 3.85ab ± 0.14 | |
| 20 | 2.90a ± 0.12 | 3.12a ± 0.11 | 3.00a ± 0.09 | 2.98a ± 0.12 | 4.31a ± 0.28 | 4.29a ± 0.24 | 4.67a ± 0.14 | 4.51a ± 0.28 | |
| Weedy rice | 40 | 2.99b ± 0.08 | 3.39a ± 0.15 | 3.25ab ± 0.12 | 3.21ab ± 0.19 | 4.60b ± 0.21 | 4.73ab ± 0.28 | 5.14a ± 0.17 | 5.01ab ± 0.22 |
| 60 | 3.28b ± 0.11 | 3.69a ± 0.06 | 3.59a ± 0.08 | 3.66a ± 0.12 | 4.99c ± 0.32 | 5.82b ± 0.38 | 6.53a ± 0.36 | 6.29ab ± 0.18 | |
| 0 | 2.74a ± 0.08 | 2.61ab0.15 | 2.45b ± 0.10 | 2.43b ± 0.04 | 3.76a ± 0.24 | 3.69a ± 0.13 | 3.98a ± 0.06 | 3.73a ± 0.10 | |
| 20 | 2.93a ± 0.02 | 3.12a ± 0.17 | 2.91a ± 0.13 | 2.90a ± 0.03 | 4.06b ± 0.34 | 4.35ab ± 0.25 | 4.78a ± 0.21 | 4.45ab ± 0.22 | |
| C. iria | 40 | 3.06b ± 0.06 | 3.46a ± 0.20 | 3.21ab ± 0.11 | 3.22ab ± 0.07 | 4.33b ± 0.27 | 4.92a ± 0.16 | 5.29a ± 0.22 | 5.01a ± 0.20 |
| 60 | 3.44b ± 0.10 | 3.82a ± 0.24 | 3.72ab ± 0.16 | 3.74ab ± 0.13 | 4.75c ± 0.26 | 6.04b ± 0.34 | 6.91a ± 0.15 | 6.49ab ± 0.21 | |
| Test Plants | Dose (gL −1) | Superoxide Dismutase (Unit g−1 FW) | Catalase (µmol g−1 FW) | Peroxidase (µmol g−1 FW) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hours after Spray | Hours after Spray | Hours after Spray | |||||||||||
| 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | 6 | 24 | 48 | 72 | ||
| 0 | 3.40a ± 0.07 | 3.35a ± 0.10 | 3.31a ± 0.04 | 3.29a ± 0.29 | 5.34a ± 0.20 | 5.15a ± 0.08 | 5.09a ± 0.11 | 5.04a ± 0.18 | 11.00c ± 1.93 | 12.77a ± 1.18 | 10.15a ± 0.92 | 12.72a ± 0.63 | |
| 20 | 4.11a ± 0.12 | 4.28a ± 0.15 | 4.25a ± 0.08 | 4.39a ± 0.38 | 6.00a ± 0.19 | 6.18a ± 0.03 | 6.26a ± 0.13 | 6.15a ± 0.20 | 12.38c ± 2.20 | 16.66ab ± 1.48 | 13.79bc ± 1.11 | 16.94a ± 0.72 | |
| A. conyzoides | 40 | 4.85a ± 0.11 | 5.10a ± 0.12 | 5.07a ± 0.01 | 5.13a ± 0.46 | 6.48b ± 0.28 | 7.27a ± 0.19 | 7.35a ± 0.15 | 7.41a ± 0.29 | 14.15b ± 2.61 | 21.82a ± 1.93 | 17.40b ± 1.66 | 21.96a ± 0.87 |
| 60 | 5.69b ± 0.10 | 6.89a ± 0.27 | 6.87a ± 0.06 | 7.11a ± 0.63 | 7.12b ± 0.36 | 8.18a ± 0.06 | 8.31a ± 0.17 | 8.28a ± 0.25 | 11.23c ± 3.08 | 27.63a ± 2.37 | 22.47b ± 1.93 | 28.22a ± 1.45 | |
| 0 | 4.01a ± 0.06 | 3.98a ± 0.04 | 3.79a ± 0.15 | 3.83a ± 0.17 | 5.88a ± 0.43 | 5.52a ± 0.25 | 5.86a ± 0.27 | 5..44a ± 0.37 | 19.74a ± 1.46 | 21.15a ± 1.08 | 21.71a ± 1.93 | 21.79a ± 1.60 | |
| 20 | 4.14a ± 0.07 | 4.31a ± 0.03 | 4.05a ± 0.15 | 4.07a ± 0.22 | 6.02a ± 0.38 | 5.80a ± 0.23 | 6.23a ± 0.31 | 5.81a ± 0.36 | 20.22b ± 1.57 | 23.94a ± 1.26 | 25.07a ± 1.91 | 25.15a ± 1.65 | |
| Weedy rice | 40 | 4.27b ± 0.06 | 4.63a ± 0.07 | 4.42ab ± 0.12 | 4.45ab ± 0.17 | 6.16a ± 0.42 | 6.26a ± 0.28 | 6.70a ± 0.34 | 6.20a ± 0.34 | 20.86b ± 1.45 | 26.70a ± 1.34 | 28.45a ± 2.32 | 29.63a ± 1.82 |
| 60 | 4.60b ± 0.12 | 5.22a ± 0.06 | 5.08a ± 0.18 | 5.17a ± 0.18 | 6.31a ± 0.40 | 6.77a ± 0.33 | 7.08a ± 0.30 | 6.69a ± 0.48 | 21.99b ± 1.46 | 32.06a ± 1.47 | 35.05a ± 3.12 | 36.09a ± 2.72 | |
| 0 | 3.99a ± 0.06 | 3.67c ± 0.06 | 3.85ab ± 0.13 | 3.73bc ± 0.03 | 6.02a ± 0.08 | 5.64ab ± 0.34 | 5.47ab ± 0.28 | 5.46b ± 0.29 | 17.48b ± 1.46 | 21.43a ± 0.92 | 19.96a ± 0.54 | 21.40a ± 0.92 | |
| 20 | 4.17ab ± 0.11 | 4.01b ± 0.05 | 4.23a ± 0.11 | 4.08ab ± 0.01 | 6.21a ± 0.12 | 6.11a ± 0.35 | 5.86a ± 0.30 | 6.01a ± 0.32 | 18.16b ± 1.40 | 24.87a ± 0.89 | 24.56a ± 0.71 | 26.31a ± 1.45 | |
| C. iria | 40 | 4.31b ± 0.05 | 4.29b ± 0.06 | 4.53a ± 0.14 | 4.41ab ± 0.05 | 6.37a ± 0.05 | 6.49a ± 0.34 | 6.48a ± 0.28 | 6.60a ± 0.31 | 18.61c ± 1.46 | 28.25ab ± 0.93 | 27.21b ± 0.54 | 29.80a ± 1.21 |
| 60 | 4.77c ± 0.06 | 5.05b ± 0.08 | 5.26a ± 0.16 | 5.21ab ± 0.02 | 6.58a ± 0.06 | 7.17a ± 0.43 | 7.01a ± 0.39 | 7.06a ± 0.37 | 19.74c ± 1.40 | 35.72ab ± 1.58 | 33.66b ± 0.70 | 36.46a ± 1.36 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motmainna, M.; Juraimi, A.S.; Uddin, M.K.; Asib, N.B.; Islam, A.K.M.M.; Ahmad-Hamdani, M.S.; Berahim, Z.; Hasan, M. Physiological and Biochemical Responses of Ageratum conyzoides, Oryza sativa f. spontanea (Weedy Rice) and Cyperus iria to Parthenium hysterophorus Methanol Extract. Plants 2021, 10, 1205. https://doi.org/10.3390/plants10061205
Motmainna M, Juraimi AS, Uddin MK, Asib NB, Islam AKMM, Ahmad-Hamdani MS, Berahim Z, Hasan M. Physiological and Biochemical Responses of Ageratum conyzoides, Oryza sativa f. spontanea (Weedy Rice) and Cyperus iria to Parthenium hysterophorus Methanol Extract. Plants. 2021; 10(6):1205. https://doi.org/10.3390/plants10061205
Chicago/Turabian StyleMotmainna, Mst., Abdul Shukor Juraimi, Md. Kamal Uddin, Norhayu Binti Asib, A. K. M. Mominul Islam, Muhammad Saiful Ahmad-Hamdani, Zulkarami Berahim, and Mahmudul Hasan. 2021. "Physiological and Biochemical Responses of Ageratum conyzoides, Oryza sativa f. spontanea (Weedy Rice) and Cyperus iria to Parthenium hysterophorus Methanol Extract" Plants 10, no. 6: 1205. https://doi.org/10.3390/plants10061205