
Rapid Detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the Loop-Mediated Isothermal Amplification (LAMP) Method

 , ,
, ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Nucleic Acid Extraction from Frass and Insects
2.2. LAMP Assay Conditions
2.3. Diagnostic Sensitivity, Specificity, and Accuracy of the LAMP Assay
2.4. Blind Panel Validation of the Assay
2.5. Repeatability and Reproducibility of the Diagnostic Methods
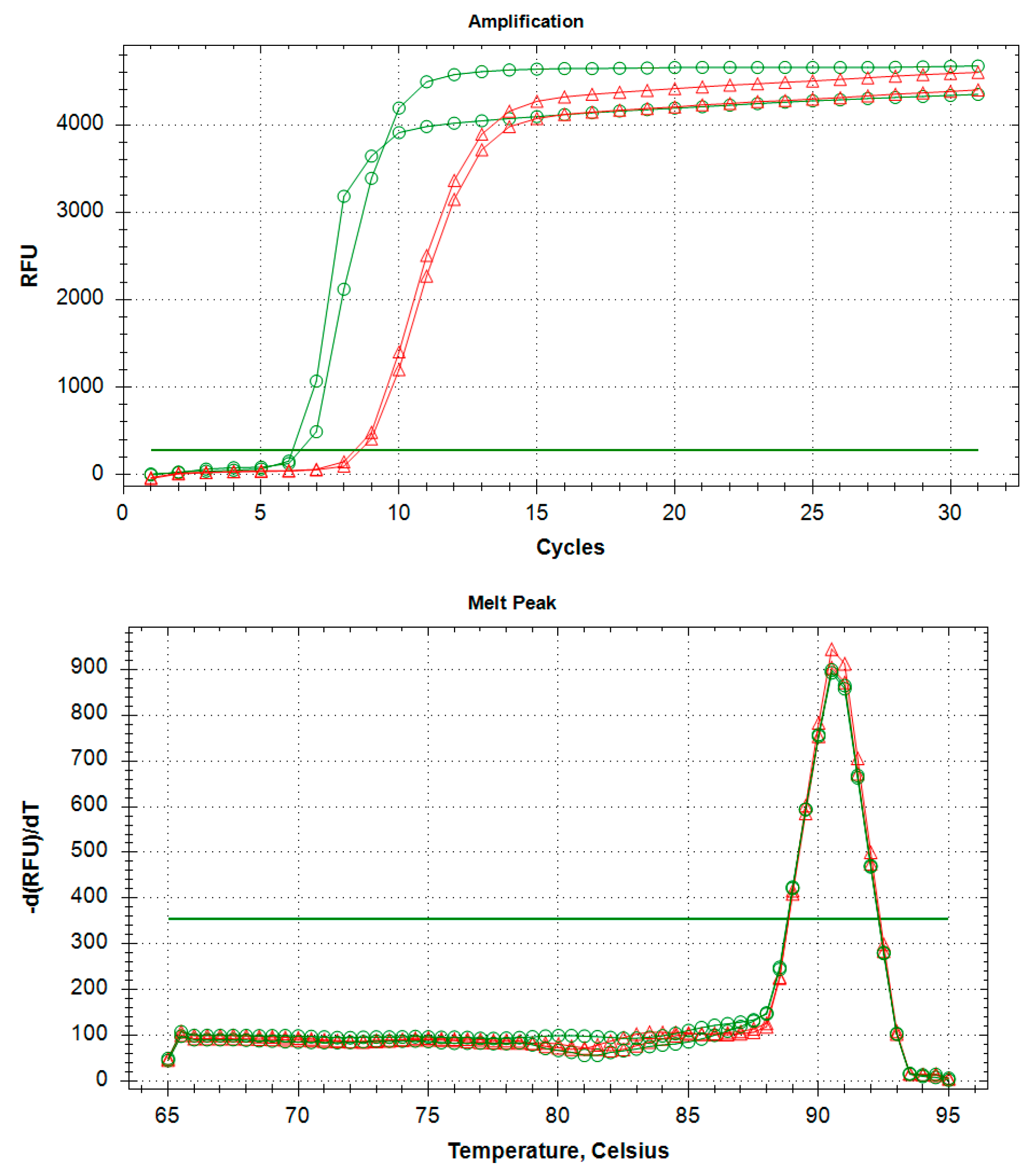
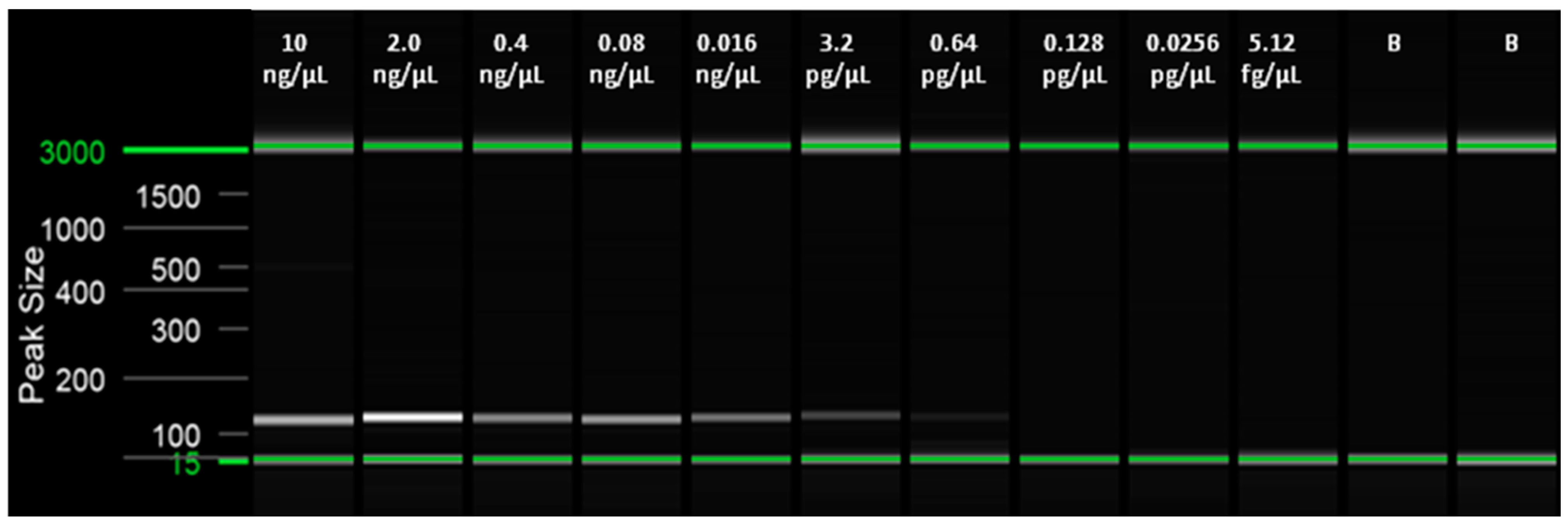
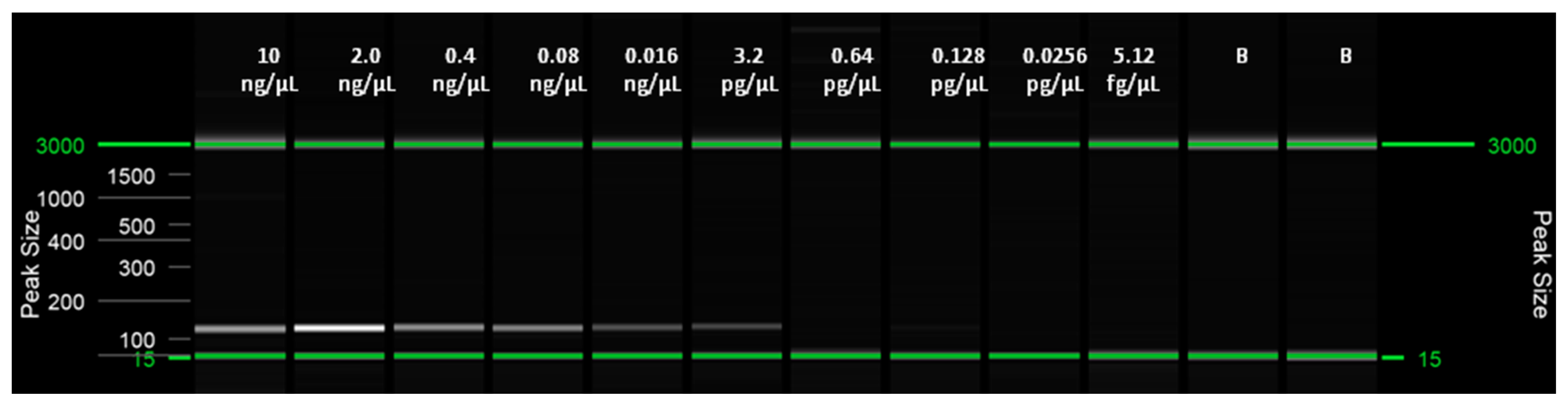
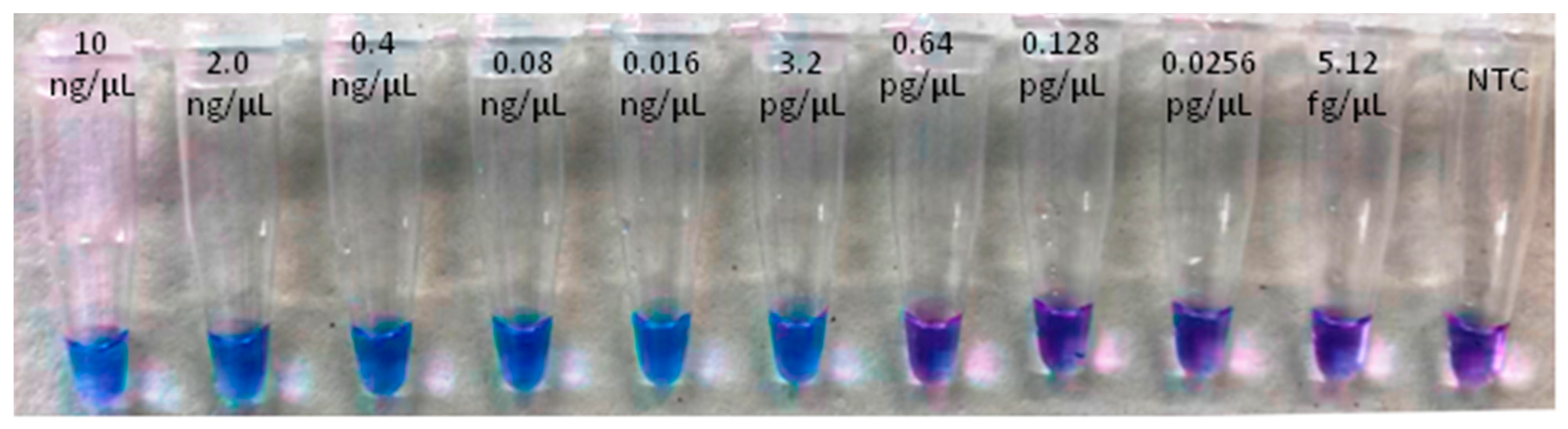
2.6. Limit of Detection (LoD) of the LAMP Assay and Comparison with qPCR (Probe) and Conventional PCR (End-Point) Assays
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. DNA Extraction
4.3. Design of the LAMP and Conventional PCR End-Point Primers
4.4. LAMP (Real Time and Visual) and End-Point PCR Assay Optimization
4.5. Performance Characteristics of the LAMP Assay
4.6. Blind Panel Validation of the Assays
4.7. Repeatability and Reproducibility
4.8. Limit of Detection (LOD)
4.9. Comparison between Conventional PCR and qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tisserat, N.; Cranshaw, W.; Leatherman, D.; Utley, C.; Alexander, K. Black Walnut Mortality in Colorado Caused by the Walnut Twig Beetle and Thousand Cankers Disease. Plant Heal. Prog. 2009, 10, 10. [Google Scholar] [CrossRef]
- Seybold, S.J.; Dallara, P.L.; Hishinuma, S.M.; Flint, M.L. Detecting and Identifying the Walnut Twig Beetle: Monitoring Guidelines for the Invasive Vector of Thousand Cankers Disease of Walnut. 2013. Available online: http://ipm.ucanr.edu/PDF/PESTNOTES/WTB_trapping.pdf (accessed on 15 January 2021).
- Kolařik, M.; Freeland, E.; Utley, C.; Tisserat, N. Geosmithia morbida sp. Nov. A new phytopathogenic species living in simbiosis with the walnut twig beetle (P. juglandis) on Juglans in USA. Mycologia 2011, 103, 325–332. [Google Scholar] [CrossRef]
- Utley, C.; Nguyen, T.; Roubtsova, T.; Coggeshall, M.; Ford, T.M.; Grauke, L.J.; Graves, A.D.; Leslie, C.A.; McKenna, J.; Woeste, K.; et al. Susceptibility of Walnut and Hickory Species to Geosmithia morbida. Plant Dis. 2013, 97, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Montecchio, L.; Faccoli, M. First Record of Thousand Cankers Disease Geosmithia morbida and Walnut Twig Beetle Pityophthorus juglandis on Juglans nigra in Europe. Plant Dis. 2014, 98, 696. [Google Scholar] [CrossRef] [PubMed]
- Moricca, S.; Bracalini, M.; Benigno, A.; Ginetti, B.; Pelleri, F.; Panzavolta, T. Thousand Cankers Disease Caused by Geosmithia morbida and Its Insect Vector Pityophthorus juglandis First Reported on Juglans nigra in Tuscany, Central Italy. Plant Dis. 2019, 103, 369. [Google Scholar] [CrossRef]
- Montecchio, L.; Fanchin, G.; Simonato, M.; Faccoli, M. First Record of Thousand Cankers Disease Fungal Pathogen Geosmithia morbida and Walnut Twig Beetle Pityophthorus juglandis on Juglans regia in Europe. Plant Dis. 2014, 98, 1445. [Google Scholar] [CrossRef] [PubMed]
- Yaghmour, M.A.; Nguyen, T.L.; Roubtsova, T.V.; Hasey, J.; Fichtner, E.J.; Debuse, C.; Seybold, S.J.; Bostock, R.M. First Report of Geosmithia morbida on English Walnut and its Paradox Rootstock in California. Plant Dis. 2014, 98, 1441. [Google Scholar] [CrossRef] [PubMed]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps for Europe: 0–13 000 B.P.; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Pelleri, F.; Castro, G.; Marchi, M.; Fernandez-Moya, J.; Chiararbaglio, P.M.; Giorcelli, A.; Bergante, S.; Gennaro, M.; Manetti, M.C.; Plutino, M.; et al. The walnut plantations (Juglans spp.) in Italy and Spain: Main factors affecting growth. Ann. Silvic. Res. 2020, 44, 14–23. [Google Scholar]
- Chahal, K.; Gazis, R.; Klingeman, W.; Hadziabdic, D.; Lambdin, P.; Grant, J.; Windham, M. Assessment of Alternative Candidate Subcortical Insect Vectors From Walnut Crowns in Habitats Quarantined for Thousand Cankers Disease. Environ. Èntomol. 2019, 48, 882–893. [Google Scholar] [CrossRef] [PubMed]
- Seybold, S.J.; Klingeman, W.; Hishinuma, S.M.; Coleman, T.W.; Graves, A.D. Status and Impact of Walnut Twig Beetle in Urban Forest, Orchard, and Native Forest Ecosystems. J. For. 2019, 117, 152–163. [Google Scholar] [CrossRef]
- Regulation (EU) 2019/2072 of 28 November 2019, Establishing Uniform Conditions for the Implementation of Regulation (EU) 2016/2031 of the European Parliament and the Council, as Regards Protective Measures against Pests of Plants, and Repealing Commission Regulation (EC) No 690/2008 and Amending Commission Implementing Regulation (EU) 2018/2019. 10.12.2019 Official Journal of the European Union L 319/1. Available online: http://data.europa.eu/eli/reg_impl/2019/2072/2020-10-06 (accessed on 18 January 2021).
- Regulation (EU) 2016/2031 of the European Parliament of the Council of 26 October 2016 on Protective Measures against Pests of Plants, Amending Regulations (EU) No 228/2013, (EU) No 652/2014 and (EU) No 1143/2014 of the European Parliament and of the Council and Repealing Council Directives 69/464/EEC, 74/647/EEC, 93/85/EEC, 98/57/EC, 2000/29/EC, 2006/91/EC and 2007/33/EC. OJ L 317 23.11.2016, 4. Available online: http://data.europa.eu/eli/reg/2016/2031/oj (accessed on 18 January 2021).
- Wilstermann, A.; Hoppe, B.; Schrader, G.; DelBianco, A.; Vos, S. Pest survey card on Geosmithia morbida and its vector Pityophthorus juglandis. EFSA Support. Publ. 2020, 17, 1894E. [Google Scholar] [CrossRef]
- Rugman-Jones, P.F.; Hoddle, C.D.; Hoddle, M.S.; Stouthamer, R. The Lesser of Two Weevils: Molecular-Genetics of Pest Palm Weevil Populations Confirm Rhynchophorus vulneratus (Panzer 1798) as a Valid Species Distinct from R. ferrugineus (Olivier 1790), and Reveal the Global Extent of Both. PLoS ONE 2013, 8, e78379. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Lewis, M.; Hoddle, M.S. Palmageddon: The wasting of ornamental palms by invasive palm weevils, Rhynchophorus spp. J. Pest Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R.A. Common pathways by which non-native forest insects move internationally and domestically. J. Pest Sci. 2019, 92, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef]
- Wang, D.; Huo, G.; Ren, D.; Li, Y. Development and evaluation of a loop-mediated isothermal amplification (lamp) method for detecting Listeria monocytogenes in raw milk. J. Food Saf. 2010, 30, 251–261. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, G.; Lu, C.; Deng, R.; Zhi, A.; Guo, J.; Zhao, D.; Xu, Z. Rapid Detection of Listeria monocytogenes in Raw Milk with Loop-Mediated Isothermal Amplification and Chemosensor. J. Food Sci. 2011, 76, M611–M615. [Google Scholar] [CrossRef]
- Mori, Y.; Kanda, H.; Notomi, T. Loop-mediated isothermal amplification (LAMP): Recent progress in research and development. J. Infect. Chemother. 2013, 19, 404–411. [Google Scholar] [CrossRef]
- Blaser, S.; Diem, H.; von Felten, A.; Gueuing, M.; Andreou, M.; Boonham, N.; Tomlinson, J.; Müller, P.; Utzinger, J.; Frey, J.E.; et al. From laboratory to point of entry: Development and implementation of a LAMP-based genetic identification system to prevent introduction of quarantine insect species. Pest Manag. Sci. 2018, 74, 1504–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchi, N.; Ioos, R.; Santini, A. Fast and reliable molecular methods to detect fungal pathogens in woody plants. Appl. Microbiol. Biotechnol. 2020, 104, 2453–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehlíková, D.; Luchi, N.; Aglietti, C.; Pepori, A.L.; Diez, J.J.; Santini, A. Real-time loop-mediated isothermal amplification assay for rapid detection of Fusarium circinatum. BioTechniques 2020, 69, 11–17. [Google Scholar] [CrossRef]
- Rizzo, D.; Luchi, N.; Da Lio, D.; Bartolini, L.; Nugnes, F.; Cappellini, G.; Bruscoli, T.; Salemi, C.; Griffo, R.V.; Garonna, A.P.; et al. Development of a loop-mediated isothermal amplification (LAMP) assay for the identification of the invasive wood borer Aromia bungii (Coleoptera: Cerambycidae) from frass. 3 Biotech 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-H.; Wang, H.-Y.; Chen, Y.-F.; Ko, C.-C. Loop-mediated isothermal amplification for rapid identification of biotypes B and Q of the globally invasive pest Bemisia tabaci, and studying population dynamics. Pest Manag. Sci. 2012, 68, 1206–1213. [Google Scholar] [CrossRef]
- Fekrat, L.; Aghl, M.Z.; Tahan, V. Application of the LAMP Assay as a Diagnostic Technique for Rapid Identification of Thrips tabaci (Thysanoptera: Thripidae). J. Econ. Èntomol. 2015, 108, 1337–1343. [Google Scholar] [CrossRef]
- Rizzo, D.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Bruscoli, T.; Bracalini, M.; Benigno, A.; Salemi, C.; Del Nista, D.; Aronadio, A.; et al. A duplex real-time PCR with probe for simultaneous detection of Geosmithia morbida and its vector Pityophthorus juglandis. PLoS ONE 2020, 15, e0241109. [Google Scholar] [CrossRef]
- Kogovšek, P.; Hodgetts, J.; A Hall, J.; Prezelj, N.; Nikolic, P.; Mehle, N.; Lenarčič, R.; Rotter, A.; Dickinson, M.; Boonham, N.; et al. LAMP assay and rapid sample preparation method for on-site detection of flavescence dorée phytoplasma in grapevine. Plant Pathol. 2015, 64, 286–296. [Google Scholar] [CrossRef]
- Bracalini, M.; Cerboneschi, M.; Croci, F.; Panzavolta, T.; Tiberi, R.; Biancalani, C.; Macconni, S.; Tegli, S. Alien pest molecular diagnostics: Can DNA traces be exploited to assess the damage caused by the western conifer seed bug on stone pine fructification? Bull. Insectology 2015, 68, 51–60. [Google Scholar]
- Poland, M.T.; Rassati, D. Improved biosecurity surveillance of non-native forest insects: A review of current methods. J. Pest Sci. 2019, 92, 37–49. [Google Scholar] [CrossRef]
- Rizzo, D.; Taddei, A.; Da Lio, D.; Nugnes, F.; Barra, E.; Stefani, L.; Bartolini, L.; Griffo, R.; Spigno, P.; Cozzolino, L.; et al. Identification of the Red-Necked Longhorn Beetle Aromia bungii (Faldermann, 1835) (Coleoptera: Cerambycidae) with Real-Time PCR on Frass. Sustainability 2020, 12, 6041. [Google Scholar] [CrossRef]
- Rizzo, D.; Taddei, A.; Da Lio, D.; Bruscoli, T.; Cappellini, G.; Bartolini, L.; Salemi, C.; Luchi, N.; Pennacchio, F.; Rossi, E. Molecular Identification of Anoplophora glabripennis (Coleoptera: Cerambycidae) From Frass by Loop-Mediated Isothermal Amplification. J. Econ. Èntomol. 2020, 113, 2911–2919. [Google Scholar] [CrossRef] [PubMed]
- Ioos, R.; Fourrier, C.; Iancu, G.; Gordon, T.R. Sensitive Detection of Fusarium circinatum in Pine Seed by Combining an Enrichment Procedure with a Real-Time Polymerase Chain Reaction Using Dual-Labeled Probe Chemistry. Phytopathology 2009, 99, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, J.; Dickinson, M.; Boonham, N. Detection of Botrytis cinerea by loop-mediated isothermal amplification. Lett. Appl. Microbiol. 2010, 51, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.H. Insect Vector Relationships with Procaryotic Plant Pathogens. Annu. Rev. Phytopathol. 1982, 20, 397–417. [Google Scholar] [CrossRef]
- Panzavolta, T.; Panichi, A.; Bracalini, M.; Croci, F.; Ginetti, B.; Ragazzi, A.; Tiberi, R.; Moricca, S. Dispersal and Propagule Pressure of Botryosphaeriaceae Species in a Declining Oak Stand is Affected by Insect Vectors. Forest 2017, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Teter, S.; Steffen, L. Real-Time qPCR: Guidelines for a Comparison of Reagent Performance; Application Note 2020, #AN299; Promega Corporation: Milan, Italy, 2020. [Google Scholar]
- Moricca, S.; Bracalini, M.; Benigno, A.; Ghelardini, L.; Furtado, E.L.; Marino, C.L.; Panzavolta, T. Observations on the non-native thousand cankers disease of walnut in Europe’s southernmost outbreak. Glob. Ecol. Conserv. 2020, 23, e01159. [Google Scholar] [CrossRef]
- Moricca, S.; Bracalini, M.; Benigno, A.; Ghelardini, L.; Furtado, E.L.; Marino, C.L.; Panzavolta, T. Thousand cankers disease in Juglans: Optimizing sampling and identification procedures for the vector Pityophthorus juglandis, and the causal agent Geosmithia morbida. MethodsX 2020, 7, 101174. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, J.; Potvin, A.; Pelletier, G.; Stewart, D.; Feau, N.; Alayon, D.I.O.; Dale, A.L.; Coelho, A.; Uzunovic, A.; Bilodeau, G.J.; et al. Molecular Detection of 10 of the Most Unwanted Alien Forest Pathogens in Canada Using Real-Time PCR. PLoS ONE 2015, 10, e0134265. [Google Scholar] [CrossRef]
- Oren, E.; Klingeman, W.; Gazis, R.; Moulton, J.; Lambdin, P.; Coggeshall, M.; Hulcr, J.; Seybold, S.J.; Hadziabdic, D. A novel molecular toolkit for rapid detection of the pathogen and primary vector of thousand cankers disease. PLoS ONE 2018, 13, e0185087. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.; Juzwik, J.; Miller, F.; Roberts, L.; Ginzel, M.D. Detection of Geosmithia morbida on Numerous Insect Species in Four Eastern States. Plant Heal. Prog. 2019, 20, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Balachowsky, A. Coléoptères scolytides. Faune de France, 50; Librairie de la Faculté des Sciences: Paris, France, 1949; p. 320. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Abdulmawjood, A.; Grabowski, N.; Fohler, S.; Kittler, S.; Nagengast, H.; Klein, G. Development of Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid and Sensitive Identification of Ostrich Meat. PLoS ONE 2014, 9, e100717. [Google Scholar] [CrossRef]
- Goto, M.; Honda, E.; Ogura, A.; Nomoto, A.; Hanaki, K.-I. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. BioTechniques 2009, 46, 167–172. [Google Scholar] [CrossRef]
- Aglietti, C.; Luchi, N.; Pepori, A.L.; Bartolini, P.; Pecori, F.; Raio, A.; Capretti, P.; Santini, A. Real-time loop-mediated isothermal amplification: An early-warning tool for quarantine plant pathogen detection. AMB Express 2019, 9. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). PM 7/98 (4) Specific requirements for laboratories preparing accreditation for a plant pest diagnostic activity. EPPO Bull. 2019, 49, 530–563. [Google Scholar] [CrossRef] [Green Version]
- Dhami, M.K.; Dsouza, M.; Waite, D.W.; Anderson, D.; Li, D. Real time PCR Assay for the Identification of the Brown Marmorated Stink Bug (Halyomorpha halys). Front. Mol. Biosci. 2016, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Koohkanzade, M.; Zakiaghl, M.; Dhami, M.K.; Fekrat, L.; Namaghi, H.S. Rapid identification of Bactrocera zonata (Dip.: Tephritidae) using TaqMan real-time PCR assay. PLoS ONE 2018, 13, e0205136. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Real Time LAMP Protocol | ||
|---|---|---|---|
| Repeatability SD | Reproducibility SD | ||
| Assay 1 | Assay 2 | ||
| 1 | 0.01 | 0.00 | 0.04 |
| 2 | 0.05 | 0.03 | 0.08 |
| 3 | 0.15 | 0.01 | 0.01 |
| 4 | 0.03 | 0.14 | 0.06 |
| 5 | 0.04 | 0.09 | 0.01 |
| 6 | 0.09 | 0.06 | 0.07 |
| 7 | 0.03 | 0.01 | 0.03 |
| 8 | 0.04 | 0.06 | 0.08 |
| Species | Classification | Matrix | Collection Date | Supplier | Source Plant/Device | Hosts |
|---|---|---|---|---|---|---|
| Pityophthorus juglandis | Coleoptera, Curculionidae, Scolytinae | frass | 2018 | PPS-T | J. nigra | Juglans spp. Pterocarya spp. |
| adult | 2018 | UF | J. nigra | |||
| Pityophthorus pubescens (Marsham) | adult | 2018 | UF | trap | conifers | |
| Ips sexdentatus (Börner) | adult | 2018 | UF | trap | conifers | |
| Ips typographus (Linnaeus) | adult | 2014 | PPS-T | trap | conifers | |
| Orthotomicus erosus (Wollaston) | adult | 2018 | UF | trap | conifers | |
| Hylurgus ligniperda (Fabricius) | adult | 2018 | UF | trap | conifers | |
| Tomicus destruens (Wollaston) | adult | 2018 | UF | trap | conifers | |
| Xyleborinus saxesenii (Ratzeburg) | adult | 2018 | UF | trap | several host genera (including Juglans) | |
| Anisandrus dispar (Fabricius) | adult | 2020 | CREA-AE | Malus sp. | several host genera (including Juglans) | |
| Xyleborus monographus (Fabricius) | adult | 2020 | CREA-AE | trap | polyphagous (including Juglans) | |
| Xylosandrus compactus (Eichhoff) | frass | 2018 | PPS-T | Laurus nobilis | several host genera | |
| adult | 2018 | PPS-T | Laurus nobilis | |||
| Xylosandrus crassiusculus (Motschulsky) | adult | 2018 | UP | Malus sp. | several host genera | |
| adult | 2019 | UP | Malus sp. | |||
| Xylosandrus germanus (Blandford) | adult | 2019 | UF | trap | several host genera (including Juglans) | |
| Lepturges confluens (Haldeman) | Coleoptera, Cerambicidae | adult | 2020 | PPS-T | Juglans sp. | several host genera (including Juglans) |
| Zeuzera pyrina (Linnaeus) | Lepidoptera, Cossidae | larva | 2017 | PPS-T | Olea europaea | several host genera (including Juglans) |
| Dilutions | Real Time LAMP | Visual LAMP | qPCR Probe P. juglandis (Rizzo et al., 2020a) | End-Point PCR (14F/125R) | |
|---|---|---|---|---|---|
| Tamp Means ±SD | Mean Melting Temperatures ±SD | Positive (+)/ -Negative (−) | Cq Means ± SD | Positive (+)/-Negative (−) | |
| 10 ng/µL | 6.76 ± 0.42 | 91.00 ± 0.00 | + | 18.31 ± 1.15 | + |
| 2.0 ng/µL | 7.76 ± 0.09 | 90.75 ± 0.35 | + | 20.69 ± 0.67 | + |
| 0.4 ng/µL | 8.25 ± 0.01 | 90.5 ± 0.00 | + | 23.05 ± 0.34 | + |
| 0.08 ng/µL | 9.60 ± 0.54 | 91.00 ± 0.00 | + | 24.57 ± 0.21 | + |
| 0.016 ng/µL | 10.17 ± 0.12 | 91.00 ± 0.00 | + | 26.85 ± 0.47 | + |
| 3.2 pg/µL | 12.92 ± 2.30 | 90.75 ± 0.35 | + | 28.35 ± 0.43 | + |
| 0.64 pg/µL | 13.96 ± 1.76 | 91.00 ± 0.00 | + | 30.16 ± 0.17 | +/− |
| 0.128 pg/µL | n/a | n/a | 32.30 ± 0.05 | n/a | |
| 0.0256 pg/µL | n/a | n/a | 33.53 ± 0.64 | n/a | |
| 5.12 fg/µL | n/a | n/a | n/a | n/a | |
| Dilutions | Real Time LAMP | Visual LAMP | qPCR Probe P. juglandis (Rizzo et al., 2020a) | End-Point PCR |
|---|---|---|---|---|
| Tamp Means ±SD | Positive (+)/-Negative (−) | Cq Means ±SD | Positive (+)/-Negative (−) | |
| 10 ng/µL | 6.70 ± 0.01 | + | 25.67 ± 1.64 | + |
| 2.0 ng/µL | 7.33 ± 0.02 | + | 28.69 ± 1.67 | + |
| 0.4 ng/µL | 8.11 ± 0.06 | + | 29.05 ± 0.34 | + |
| 0.08 ng/µL | 9.16 ± 0.08 | + | 31.64 ± 1.21 | + |
| 0.016 ng/µL | 9.65 ± 0.59 | + | 33.76 ± 1.47 | + |
| 3.2 pg/µL | 13.59 ± 1.38 | + | 35.64 ± 1.74 | + |
| 0.64 pg/µL | n/a | n/a | n/a | n/a |
| 0.128 pg/µL | n/a | n/a | n/a | n/a |
| 0.0256 pg/µL | n/a | n/a | n/a | n/a |
| 5.12 fg/µL | n/a | n/a | n/a | n/a |
| Primer Name | Length (nt) | Sequence 5′-3′ | Nucleotide Position | Product Size (bp) | Reference Sequence |
|---|---|---|---|---|---|
| Pjug_B3 | 18 | GTCGCAGATCGGTCTTAAG | 538-519 | 160 bp | KP201676 |
| Pjug_BIP(B1c + B2) | 39 | ACATGTTGGCGATCGGACCGAGAACTCGACAGCTAACAG | 446-465 | ||
| 509-489 | |||||
| Pjug_F3 | 19 | TCGATCTAAGGTCCACGG | 303-321 | ||
| Pjug_FIP(F1c + F2) | 39 | CGGTCGAACGCTCATAGGAGGTTAACGGACCCGTGAAAT | 429-449 | ||
| 350-331 | |||||
| Pjug_LoopB | 19 | TGCCGATTCTGACATCCG | 468-486 | ||
| Pjug_LoopF | 18 | AACCGTTCGTATACCGTCG | 401-382 |
| Primers | Sequence (5′-3′) | Length | Annealing | Type of Protocol | Reference |
|---|---|---|---|---|---|
| Pjug_14_F | GCATAGTAGGGACCTCACTTAGTG | 112 bp | 55 °C | End point | This study |
| Pjug_125_R | ATAAAGGCATGGGCTGTTACTACA | ||||
| Pjug_253_F | TCCCACGTCTTAATAATATAAG | 183 bp | 55 °C | qPCR Probe | Rizzo et al., 2020a |
| Pjug_435_R | CTCCTGCTATATGAAGACTA | ||||
| Pjug_281_P | Hex_ACTCTTACCACCATCATTAACATTCCT_BHQ1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzo, D.; Moricca, S.; Bracalini, M.; Benigno, A.; Bernardo, U.; Luchi, N.; Da Lio, D.; Nugnes, F.; Cappellini, G.; Salemi, C.; et al. Rapid Detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the Loop-Mediated Isothermal Amplification (LAMP) Method. Plants 2021, 10, 1048. https://doi.org/10.3390/plants10061048
Rizzo D, Moricca S, Bracalini M, Benigno A, Bernardo U, Luchi N, Da Lio D, Nugnes F, Cappellini G, Salemi C, et al. Rapid Detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the Loop-Mediated Isothermal Amplification (LAMP) Method. Plants. 2021; 10(6):1048. https://doi.org/10.3390/plants10061048
Chicago/Turabian StyleRizzo, Domenico, Salvatore Moricca, Matteo Bracalini, Alessandra Benigno, Umberto Bernardo, Nicola Luchi, Daniele Da Lio, Francesco Nugnes, Giovanni Cappellini, Chiara Salemi, and et al. 2021. "Rapid Detection of Pityophthorus juglandis (Blackman) (Coleoptera, Curculionidae) with the Loop-Mediated Isothermal Amplification (LAMP) Method" Plants 10, no. 6: 1048. https://doi.org/10.3390/plants10061048