
So Much for Glucosinolates: A Generalist Does Survive and Develop on Brassicas, but at What Cost?

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Diet and Food Plant Composition
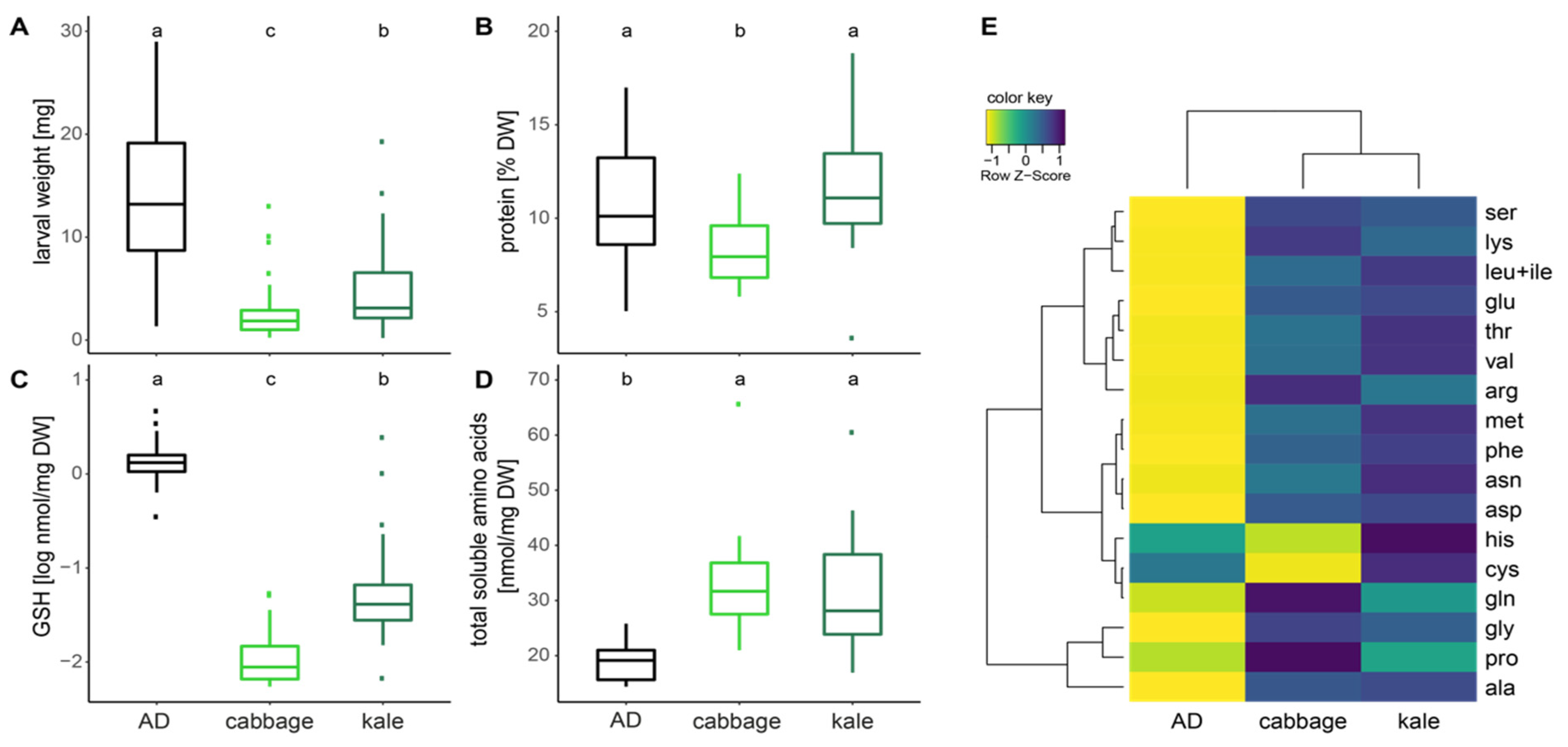
2.2. Caterpillar Development and Body Chemistry
2.3. Adult Body Weight and Composition
2.4. Conjugate Excretion in Faeces
3. Discussion
4. Materials and Methods
4.1. Plants
4.2. Plant Chemistry and Diet Composition
4.3. Insects
4.4. Experimental Protocol
4.5. Caterpillar and Adult Body Chemistry
4.6. Artificial Diet Feeding and Excretion Assays
4.7. Non-Targeted UHPLC-MS Analyses of Larval Frass
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Burow, M.; Losansky, A.; Müller, R.; Plock, A.; Kliebenstein, D.J.; Wittstock, U. The Genetic Basis of Constitutive and Herbivore-Induced ESP-Independent Nitrile Formation in Arabidopsis. Plant Physiol. (Bethesda) 2009, 149, 561–574. [Google Scholar] [CrossRef] [Green Version]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Ann. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [Green Version]
- Matile, P. The mustard oil bomb. Compartmentation of the myrosinase system. Biochem. Physiol. Pflanz. 1980, 175, 722–731. [Google Scholar] [CrossRef]
- Wittstock, U.; Burow, M. Tipping the Scales—Specifier Proteins in Glucosinolate Hydrolysis. IUBMB Life 2007, 59, 744–751. [Google Scholar] [CrossRef]
- Hopkins, R.J.; Dam, N.M.V.; Loon, J.J.A.V. Role of Glucosinolates in Insect-Plant Relationships and Multitrophic Interactions. Ann. Rev. Entomol. 2009, 54, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Eigenbrode, S.D.; Stringam, G.R.; Thiagarajah, M.R. Feeding and Growth of Plutella xylostella and Spodoptera eridania on Brassica juncea with Varying Glucosinolate Concentrations and Myrosinase Activities. J. Chem. Ecol. 2000, 26, 2401–2419. [Google Scholar] [CrossRef]
- Müller, R.; de Vos, M.; Sun, J.Y.; Sønderby, I.E.; Halkier, B.A.; Wittstock, U.; Jander, G. Differential Effects of Indole and Aliphatic Glucosinolates on Lepidopteran Herbivores. J. Chem. Ecol. 2010, 36, 905–913. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Interaction of glucosinolate content of Arabidopsis thaliana mutant lines and feeding and oviposition by generalist and specialist lepidopterans. Phytochemistry (Oxford) 2013, 86, 36–43. [Google Scholar] [CrossRef]
- Bejai, S.; Fridborg, I.; Ekbom, B. Varied response of Spodoptera littoralis against Arabidopsis thaliana with metabolically engineered glucosinolate profiles. Plant Physiol. Biochem. 2012, 50, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Zalucki, M.P.; Zalucki, J.M.; Perkins, L.E.; Schramm, K.; Vassão, D.G.; Gershenzon, J.; Heckel, D.G. A Generalist Herbivore Copes with Specialized Plant Defence: The Effects of Induction and Feeding by Helicoverpa armigera (Lepidoptera: Noctuidae) Larvae on Intact Arabidopsis thaliana (Brassicales) Plants. J. Chem. Ecol. 2017, 43, 608–616. [Google Scholar] [CrossRef]
- Zalucki, J.M.; Heckel, D.G.; Wang, P.; Kuwar, S.; Vassão, D.G.; Perkins, L.E.; Zalucki, M.P. A generalist feeding on Brassica; it does not get any better with selection. Plants 2021, 10, 954. [Google Scholar] [CrossRef]
- Jeschke, V.; Gershenzon, J.; Vassão, D.G. A mode of action of glucosinolate-derived isothiocyanates: Detoxification depletes glutathione and cysteine levels with ramifications on protein metabolism in Spodoptera littoralis. Insect Biochem. Mol. Biol. 2016, 71, 37–48. [Google Scholar] [CrossRef]
- Schramm, K.; Vassão, D.G.; Reichelt, M.; Gershenzon, J.; Wittstock, U. Metabolism of glucosinolate-derived isothiocyanates to glutathione conjugates in generalist lepidopteran herbivores. Insect Biochem. Mol. Biol. 2012, 42, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Katsikis, C.I.; Wang, P.; Zalucki, M.P. Life history traits of a key agricultural pest, Helicoverpa armigera (Lepidoptera: Noctuidae): Are laboratory settings appropriate? Aust. Entomol. 2020, 59, 189–201. [Google Scholar] [CrossRef]
- Gols, R.; Bukovinszky, T.; Van Dam, N.M.; Dicke, M.; Bullock, J.M.; Harvey, J.A. Performance of generalist and specialist herbivores and their endoparasitoids differs on cultivated and wild Brassica populations. J. Chem. Ecol. 2008, 34, 132–143. [Google Scholar] [CrossRef]
- Falk, K.L.; Tokuhisa, J.G.; Gershenzon, J. The Effect of Sulfur Nutrition on Plant Glucosinolate Content: Physiology and Molecular Mechanisms. Plant Biol. 2007, 9, 573–581. [Google Scholar] [CrossRef]
- Hwang, I.M.; Park, B.; Dang, Y.M.; Kim, S.-Y.; Seo, H.Y. Simultaneous direct determination of 15 glucosinolates in eight Brassica species by UHPLC-Q-Orbitrap-MS. Food Chem. 2019, 282, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.R.; Jo, J.S.; Lee, J.G. Comparison of glucosinolate profiles in different tissues of nine brassica crops. Molecules 2015, 20, 15827–15841. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Neyazaki, M.; Shindo, K.; Ogawa, T.; Momose, M. Quantitative profiling of glucosinolates by LC–MS analysis reveals several cultivars of cabbage and kale as promising sources of sulforaphane. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 903, 171–176. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P.; Obregón, S.; Padilla, G.; de Haro, A. Seasonal variation in glucosinolate content in Brassica oleracea crops grown in northwestern Spain. Phytochemistry (Oxford) 2008, 69, 403–410. [Google Scholar] [CrossRef]
- Choi, S.-H.; Park, S.; Lim, Y.P.; Kim, S.-J.; Park, J.-T.; An, G. Metabolite profiles of glucosinolates in cabbage varieties (Brassica oleracea var. capitata) by season, color, and tissue position. Horticult. Environ. Biotechnol. 2014, 55, 237–247. [Google Scholar] [CrossRef]
- Wittstock, U.; Kliebenstein, D.J.; Lambrix, V.; Reichelt, M.; Gershenzon, J. Chapter five Glucosinolate hydrolysis and its impact on generalist and specialist insect herbivores. Recent Adv. Phytochem. 2003, 37, 101–125. [Google Scholar]
- Wang, P.; Vassao, D.G.; Raguschke, B.; Furlong, M.J.; Zalucki, M.P. Balancing nutrients in a toxic environment: The challenge of eating. Insect Sci. 2021, 1–17. [Google Scholar] [CrossRef]
- Williamson, J.M.; Boettcher, B.; Meister, A. Intracellular Cysteine Delivery System That Protects against Toxicity by Promoting Glutathione Synthesis. Proc. Natl. Acad. Sci. USA 1982, 79, 6246–6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbehenn, R.V.; Kochmanski, J.; Menachem, B.; Poirier, L.M. Allocation of Cysteine for Glutathione Production in Caterpillars with Different Antioxidant Defense Strategies: A Comparison of Lymantria Dispar and Malacosoma Disstria. Arch. Insect Biochem. Physiol. 2013, 84, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Gols, R.; Harvey, J.A.; Reichelt, M.; Gershenzon, J.; Pandit, S.S.; Vassão, D.G. Detoxification of plant defensive glucosinolates by an herbivorous caterpillar is beneficial to its endoparasitic wasp. Mol. Ecol. 2020, 29, 4014–4031. [Google Scholar] [CrossRef]
- Jagdale, S.; Tellis, M.; Barvkar, V.T.; Joshi, R.S. Glucosinolate induces transcriptomic and metabolic reprogramming in Helicoverpa armigera. 3 Biotech 2021, 11, 26. [Google Scholar] [CrossRef]
- Li, S.; Yu, X.; Feng, Q. Fat Body Biology in the Last Decade. Ann. Rev. Entomol. 2019, 64, 315–333. [Google Scholar] [CrossRef]
- Jeschke, V.; Kearney, E.E.; Schramm, K.; Kunert, G.; Shekhov, A.; Gershenzon, J.; Vassão, D.G. How glucosinolates affect generalist lepidopteran larvae: Growth, development and glucosinolate metabolism. Front. Plant Sci. 2017, 8, 1995. [Google Scholar] [CrossRef] [Green Version]
- Schlaeppi, K.; Bodenhausen, N.; Buchala, A.; Mauch, F.; Reymond, P. The glutathione-deficient mutant pad2-1 accumulates lower amounts of glucosinolates and is more susceptible to the insect herbivore Spodoptera littoralis. Plant J. Cell Mol. Biol. 2008, 55, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Benrey, B.; Denno, R.F. The slow-growth-high-mortality hypothesis: A test using the cabbage butterfly. Ecology (Durham) 1997, 78, 987–999. [Google Scholar] [CrossRef]
- Clancy, K.M.; Price, P.W. Rapid herbivore growth enhances enemy attack: Sublethal plant defenses remain a paradox. Ecology (Durham) 1987, 68, 733–737. [Google Scholar] [CrossRef]
- Proffit, M.; Khallaf, M.A.; Carrasco, D.; Larsson, M.C.; Anderson, P.; Dam, N. ‘Do you remember the first time?’ Host plant preference in a moth is modulated by experiences during larval feeding and adult mating. Ecol. Lett. 2015, 18, 365–374. [Google Scholar] [CrossRef]
- Thöming, G.; Larsson, M.C.; Hansson, B.S.; Anderson, P. Comparison of plant preference hierarchies of male and female moths and the impact of larval rearing hosts. Ecology (Durham) 2013, 94, 1744–1752. [Google Scholar] [CrossRef]
- Docimo, T.; Reichelt, M.; Schneider, B.; Kai, M.; Kunert, G.; Gershenzon, J.; D’Auria, J.C. The first step in the biosynthesis of cocaine in Erythroxylum coca: The characterization of arginine and ornithine decarboxylases. Plant Mol. Biol. 2012, 78, 599–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celorio-Mancera, M.D.L.P.; Heckel, D.G.; Vogel, H. Transcriptional analysis of physiological pathways in a generalist herbivore: Responses to different host plants and plant structures by the cotton bollworm, Helicoverpa armigera. Entomol. Exp. Appl. 2012, 144, 123–133. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Linear and Nonlinear Mixed Effects Models. R Package Version 2007, 3, 1–89. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeschke, V.; Zalucki, J.M.; Raguschke, B.; Gershenzon, J.; Heckel, D.G.; Zalucki, M.P.; Vassão, D.G. So Much for Glucosinolates: A Generalist Does Survive and Develop on Brassicas, but at What Cost? Plants 2021, 10, 962. https://doi.org/10.3390/plants10050962
Jeschke V, Zalucki JM, Raguschke B, Gershenzon J, Heckel DG, Zalucki MP, Vassão DG. So Much for Glucosinolates: A Generalist Does Survive and Develop on Brassicas, but at What Cost? Plants. 2021; 10(5):962. https://doi.org/10.3390/plants10050962
Chicago/Turabian StyleJeschke, Verena, Jacinta M. Zalucki, Bettina Raguschke, Jonathan Gershenzon, David G. Heckel, Myron P. Zalucki, and Daniel G. Vassão. 2021. "So Much for Glucosinolates: A Generalist Does Survive and Develop on Brassicas, but at What Cost?" Plants 10, no. 5: 962. https://doi.org/10.3390/plants10050962