
Anti-Osteoporotic Effects of Commiphora Myrrha and Its Poly-Saccharide via Osteoclastogenesis Inhibition


Abstract
:1. Introduction
2. Results
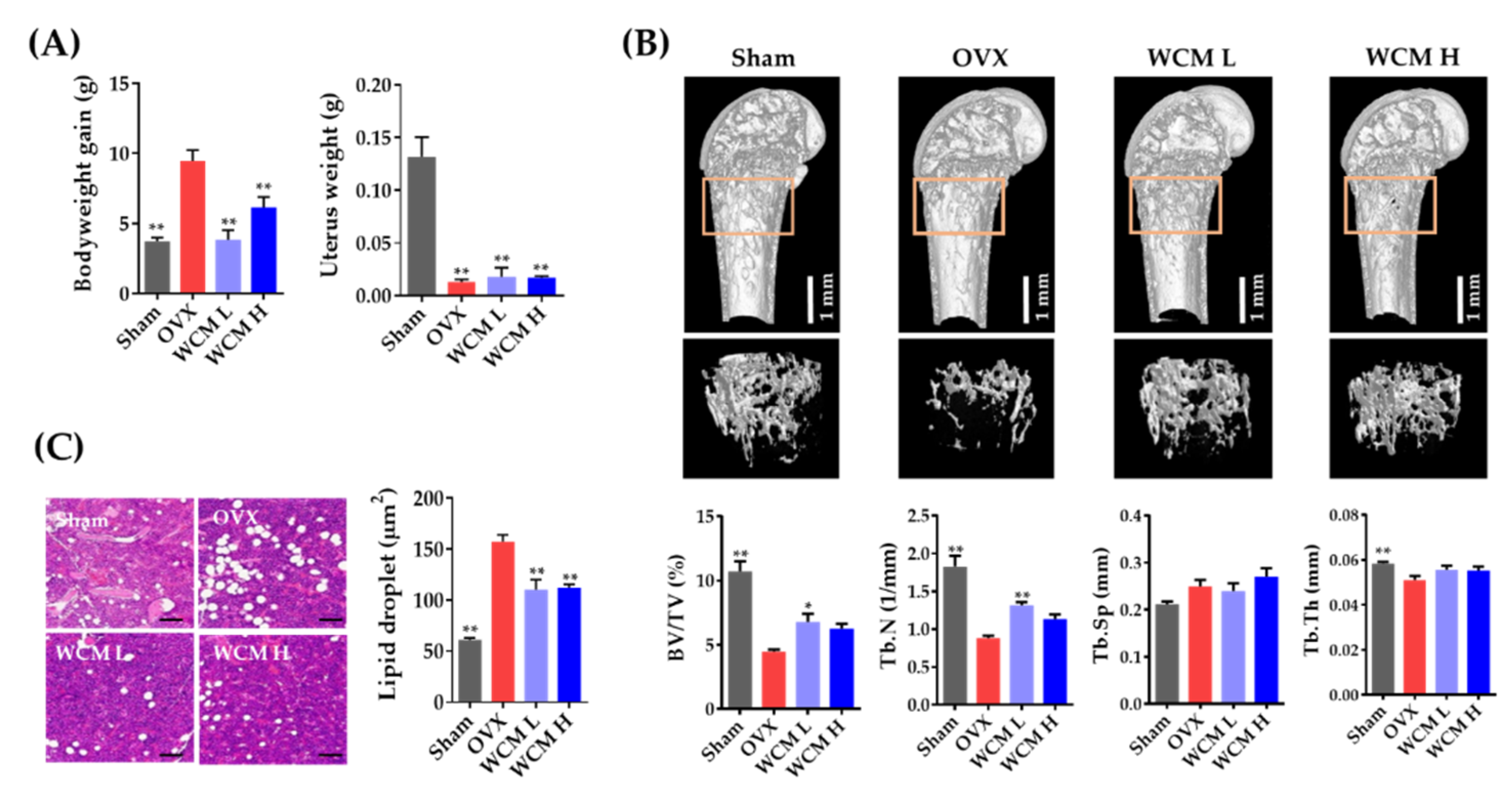
2.1. WCM Protected Estrogen Deprivation-Induced Bone Loss in Mice
2.2. Phytochemical Profile of WCM and WCM-PE
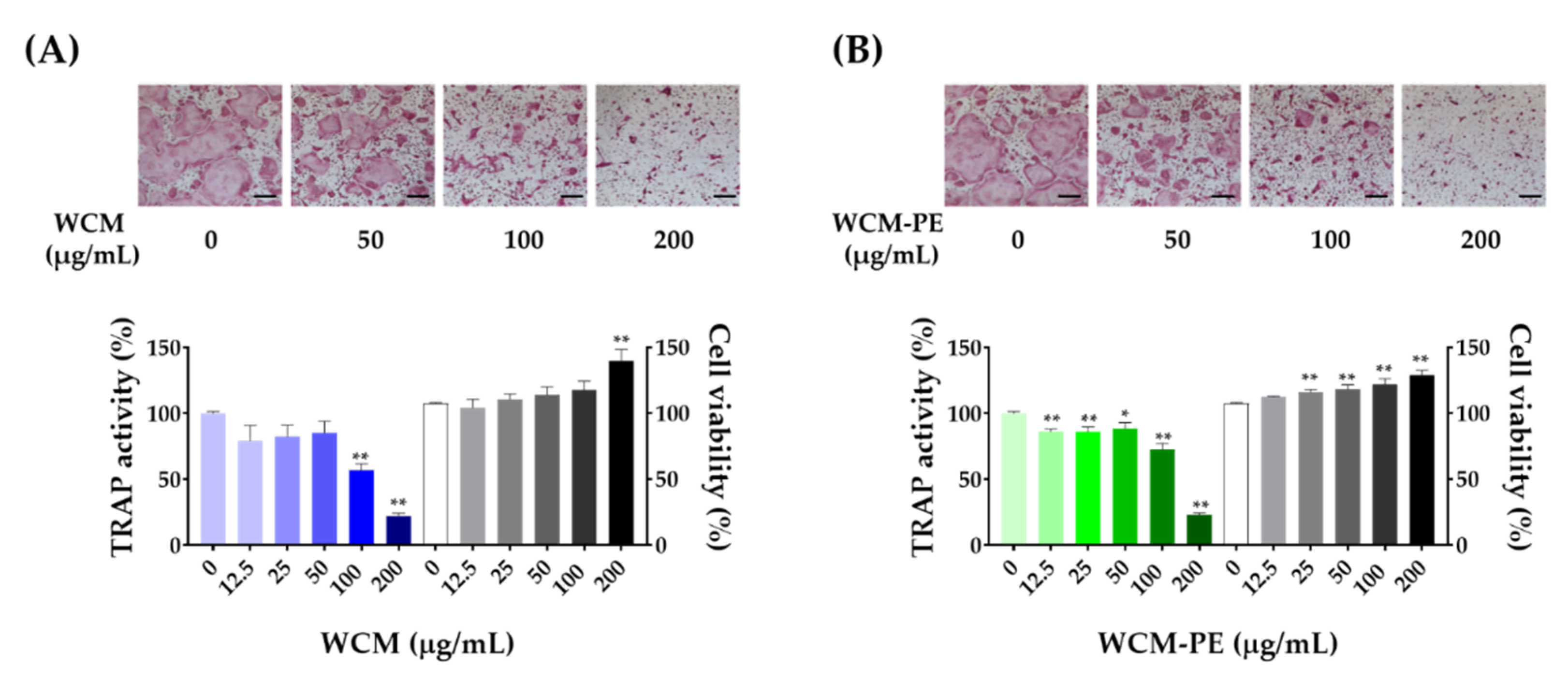
2.3. WCM and WCM-PE Inhibited Osteoclastogenesis
2.4. WCM and WCM-PE Suppressed Key Transcription Factors and Osteoclast-Specific Genes
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of WCM and WCM-PE
4.3. Chemical Profiling of WCM and WCM-PE
4.4. Animal Study
4.5. Osteoclast Differentiation Assay
4.6. Real-Time PCR and Western Blotting
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, M.X.; Yu, Q. Primary Osteoporosis in Postmenopausal Women. Chronic Dis. Transl. Med. 2015, 1, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast Differentiation and Activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Pereira, M.; Petretto, E.; Gordon, S.; Bassett, J.H.D.; Williams, G.R.; Behmoaras, J. Common Signalling Pathways in Macrophage and Osteoclast Multinucleation. J. Cell Sci. 2018, 131, 216267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Baron, R.; Ferrari, S.; Russell, R.G. Denosumab and Bisphosphonates: Different Mechanisms of Action and Effects. Bone 2011, 48, 677–692. [Google Scholar] [CrossRef]
- Reid, I.R. Efficacy, Effectiveness and Side Effects of Medications Used to Prevent Fractures. J. Int. Med. 2015, 277, 690–706. [Google Scholar] [CrossRef]
- Wang, X.; Yamauchi, K.; Mitsunaga, T. A Review on Osteoclast Diseases and Osteoclastogenesis Inhibitors Recently Developed from Natural Resources. Fitoterapia 2020, 142, 104482. [Google Scholar] [CrossRef] [PubMed]
- Banu, J.; Varela, E.; Fernandes, G. Alternative Therapies for The Prevention and Treatment of Osteoporosis. Nutr. Rev. 2012, 70, 22–40. [Google Scholar] [CrossRef]
- Wang, T.; Liu, Q.; Tjhioe, W.; Zhao, J.; Lu, A.; Zhang, G.; Tan, R.X.; Zhou, M.; Xu, J.; Feng, H.T. Therapeutic Potential and Outlook of Alternative Medicine for Osteoporosis. Curr. Drug Targets 2017, 18, 1051–1068. [Google Scholar] [CrossRef]
- He, J.; Li, X.; Wang, Z.; Bennett, S.; Chen, K.; Xiao, Z.; Zhan, J.; Chen, S.; Hou, Y.; Chen, J.; et al. Therapeutic Anabolic and Anticatabolic Benefits of Natural Chinese Medicines for the Treatment of Osteoporosis. Front. Pharmacol. 2019, 10, 1344. [Google Scholar] [CrossRef]
- Tit, D.M.; Bungau, S.; Iovan, C.; Nistor Cseppento, D.C.; Endres, L.; Sava, C.; Sabau, A.M.; Furau, G.; Furau, C. Effects of the Hormone Replacement Therapy and of Soy Isoflavones on Bone Resorption in Postmenopause. J. Clin. Med. 2018, 7, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haffor, A.S. Effect of Myrrh (Commiphora molmol) on Leukocyte Levels Before and During Healing from Gastric Ulcer or Skin Injury. J. Immunotoxicol. 2010, 7, 68–75. [Google Scholar] [CrossRef]
- Shen, T.; Li, G.-H.; Wang, X.-N.; Lou, H.-X. The Genus Commiphora: A Review of Its Traditional Uses, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2012, 142, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Hanus, L.O.; Rezanka, T.; Dembitsky, V.M.; Moussaieff, A. Myrrh-Commiphora Chemistry. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2005, 149, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, M.A.; Hammouda, A.A. Analgesic, Anti-Inflammatory and Anti-Hyperlipidemic Activities of Commiphora Molmol Extract (Myrrh). J. Intercult. Ethnopharmacol. 2014, 3, 56–62. [Google Scholar] [CrossRef]
- Fatani, A.J.; Alrojayee, F.S.; Parmar, M.Y.; Abuohashish, H.M.; Ahmed, M.M.; Al-Rejaie, S.S. Myrrh Attenuates Oxidative and Inflammatory Processes in Acetic Acid-Induced Ulcerative Colitis. Exp. Ther. Med. 2016, 12, 730–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orabi, S.H.; Al-Sabbagh, E.S.; Khalifa, H.K.; Mohamed, M.A.E.-G.; Elhamouly, M.; Gad-Allah, S.M.; Abdel-Daim, M.M.; Eldaim, M.A.A. Commiphora myrrha Resin Alcoholic Extract Ameliorates High Fat Diet Induced Obesity via Regulation of UCP1 and Adiponectin Proteins Expression in Rats. Nutrients 2020, 12, 803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Ashry, E.S.; Rashed, N.; Salama, O.M.; Saleh, A. Components, Therapeutic Value and Uses of Myrrh. Pharmazie 2003, 58, 163–168. [Google Scholar]
- Turner, A.S. Animal Models of Osteoporosis--Necessity and Limitations. Eur. Cells Mater. 2001, 1, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Saleh, N.; Nassef, N.A.; Shawky, M.K.; Elshishiny, M.I.; Saleh, H.A. Novel Approach for Pathogenesis of Osteoporosis in Ovariectomized Rats as a Model of Postmenopausal Osteoporosis. Exp. Gerontol. 2020, 137, 110935. [Google Scholar] [CrossRef] [PubMed]
- Shim, K.S.; Hwang, Y.H.; Jang, S.A.; Kim, T.; Ha, H. Water Extract of Lysimachia christinae Inhibits Trabecular Bone Loss and Fat Accumulation in Ovariectomized Mice. Nutrients 2020, 12, 1927. [Google Scholar] [CrossRef] [PubMed]
- Corina, M.; Vulpoi, C.; Brănişteanu, D. Relationship between Bone Mineral Density, Weight, and Estrogen Levels in Pre and Postmenopausal Women. Rev. Med. Chir. Soc. Med. Nat. Iasi 2012, 116, 946–950. [Google Scholar]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Müller, R. Guidelines for Assessment of Bone Microstructure in Rodents Using Micro-Computed Tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Bi, Y.N.; Zhang, P.Y.; Yuan, X.M.; Liu, Y.; Zhang, Y.; Huang, J.Y.; Zhou, K. Optimization of the Time Window of Interest in Ovariectomized Imprinting Control Region Mice for Antiosteoporosis Research. BioMed Res. Int. 2017, 2017, 8417814. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Takeshita, S. The Role of Osteoclast Differentiation and Function in Skeletal Homeostasis. J. Biol. Chem. 2016, 159, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Arai, A.; Mizoguchi, T.; Harada, S.; Kobayashi, Y.; Nakamichi, Y.; Yasuda, H.; Penninger, J.M.; Yamada, K.; Udagawa, N.; Takahashi, N. Fos Plays an Essential Role in the Upregulation of RANK Expression in Osteoclast Precursors within the Bone Microenvironment. J. Cell. Sci. 2012, 125, 2910–2917. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H.; Kim, S.; Matsuo, K.; Suzuki, H.; Suzuki, T.; Sato, K.; Yokochi, T.; Oda, H.; Nakamura, K.; Ida, N.; et al. RANKL Maintains Bone Homeostasis through c-Fos-Dependent Induction of Interferon-beta. Nature 2002, 416, 744–749. [Google Scholar] [CrossRef]
- Matsuo, K.; Galson, D.L.; Zhao, C.; Peng, L.; Laplace, C.; Wang, K.Z.Q.; Bachler, M.A.; Amano, H.; Aburatani, H.; Ishikawa, H.; et al. Nuclear Factor of Activated T-cells (NFAT) Rescues Osteoclastogenesis in Precursors Lacking c-Fos. J. Biol. Chem. 2004, 279, 26475–26480. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Lee, S.H.; Ha Kim, J.; Choi, Y.; Kim, N. NFATc1 Induces Osteoclast Fusion via Up-Regulation of Atp6v0d2 and the Dendritic Cell-Specific Transmembrane Protein (DC-STAMP). Mol. Endocrinol. 2008, 22, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Kodama, J.; Kaito, T. Osteoclast Multinucleation: Review of Current Literature. Int. J. Mol. Sci. 2020, 21, 5685. [Google Scholar] [CrossRef]
- Mohammed, A.S.A.; Naveed, M.; Jost, N. Polysaccharides; Classification, Chemical Properties, and Future Perspective Applications in Fields of Pharmacology and Biological Medicine (A Review of Current Applications and Upcoming Potentialities). J. Polym. Environ. 2021. [Google Scholar] [CrossRef]
- Li, Q.; Niu, Y.; Xing, P.; Wang, C. Bioactive Polysaccharides from Natural Resources Including Chinese Medicinal Herbs on Tissue Repair. Chin. Med. 2018, 13, 7. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Wei, B.; Peng, Z.; Chen, X.; Fu, Q.; Wang, C.; Zhen, J.; Sun, J. A Polysaccharide from the Dried Rhizome of Drynaria fortunei (Kunze) J. Sm. Prevents Ovariectomized (OVX)-Induced Osteoporosis in Rats. J. Cell. Mol. Med. 2020, 24, 3692–3700. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Jang, S.A.; Lee, A.; Cho, C.W.; Song, Y.R.; Hong, H.D.; Ha, H.; Kim, T. Polysaccharides Isolated from Lotus Leaves (LLEP) Exert Anti-Osteoporotic Effects by Inhibiting Osteoclastogenesis. Int. J. Biol. Macromol. 2020, 161, 449–456. [Google Scholar] [CrossRef]
- Ye, Z.; Li, T.; Qing, D.; Sun, Y.; Chen, H.; Yu, Q.; Yan, C. Structural Elucidation and Osteogenic Activity of a Novel Heteropolysaccharide from Alhagi pseudalhagi. Int. J. Biol. Macromol. 2021, 171, 185–197. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Jang, S.A.; Kim, T.; Ha, H. Forsythia suspensa Protects against Bone Loss in Ovariectomized Mice. Nutrients 2019, 11, 1831. [Google Scholar] [CrossRef] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New Method for Quantitative Determination of Uronic Acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Karkhanis, Y.D.; Zeltner, J.Y.; Jackson, J.J.; Carlo, D.J. A New and Improved Microassay to Determine 2-Keto-3-Deoxyoctonate in Lipopolysaccharide of Gram-Negative Bacteria. Anal. Biochem. 1978, 85, 595–601. [Google Scholar] [CrossRef]
- Xu, G.; Amicucci, M.J.; Cheng, Z.; Galermo, A.G.; Lebrilla, C.B. Revisiting Monosaccharide Analysis–Quantitation of a Comprehensive Set of Monosaccharides Using Dynamic Multiple Reaction Monitoring. Analyst 2018, 143, 200–207. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Ha, H.; Kim, R.; Cho, C.W.; Song, Y.R.; Hong, H.D.; Kim, T. Anti-Osteoporotic Effects of Polysaccharides Isolated from Persimmon Leaves via Osteoclastogenesis Inhibition. Nutrients 2018, 10, 901. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WCM-PE | |
|---|---|
| Chemical composition (%) | |
| Neutral sugar | 67.78 ± 1.79 |
| Uronic acid | 34.58 ± 0.36 |
| 2-keto-3-deoxy-mannooctanoic acid | 0.20 ± 0.02 |
| Protein | 2.65 ± 0.13 |
| Component sugar (mol%) | |
| Arabinose | 29.84 |
| Fucose | 0.78 |
| Galactose | 65.56 |
| Rhamnose | 0.96 |
| Glucoronic acid | 2.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, Y.-H.; Lee, A.; Kim, T.; Jang, S.-A.; Ha, H. Anti-Osteoporotic Effects of Commiphora Myrrha and Its Poly-Saccharide via Osteoclastogenesis Inhibition. Plants 2021, 10, 945. https://doi.org/10.3390/plants10050945
Hwang Y-H, Lee A, Kim T, Jang S-A, Ha H. Anti-Osteoporotic Effects of Commiphora Myrrha and Its Poly-Saccharide via Osteoclastogenesis Inhibition. Plants. 2021; 10(5):945. https://doi.org/10.3390/plants10050945
Chicago/Turabian StyleHwang, Youn-Hwan, Ami Lee, Taesoo Kim, Seon-A Jang, and Hyunil Ha. 2021. "Anti-Osteoporotic Effects of Commiphora Myrrha and Its Poly-Saccharide via Osteoclastogenesis Inhibition" Plants 10, no. 5: 945. https://doi.org/10.3390/plants10050945