
Differences in Leaf Morphological Parameters of Pear (Pyrus communis L.) Based on Their Susceptibility to European Pear Rust Caused by Gymnosporangium sabinae (Dicks.) Oerst.


Abstract
:1. Introduction
2. Results
2.1. Evaluation of G. sabinae Severity
2.2. Evaluation of Morphological Parameters
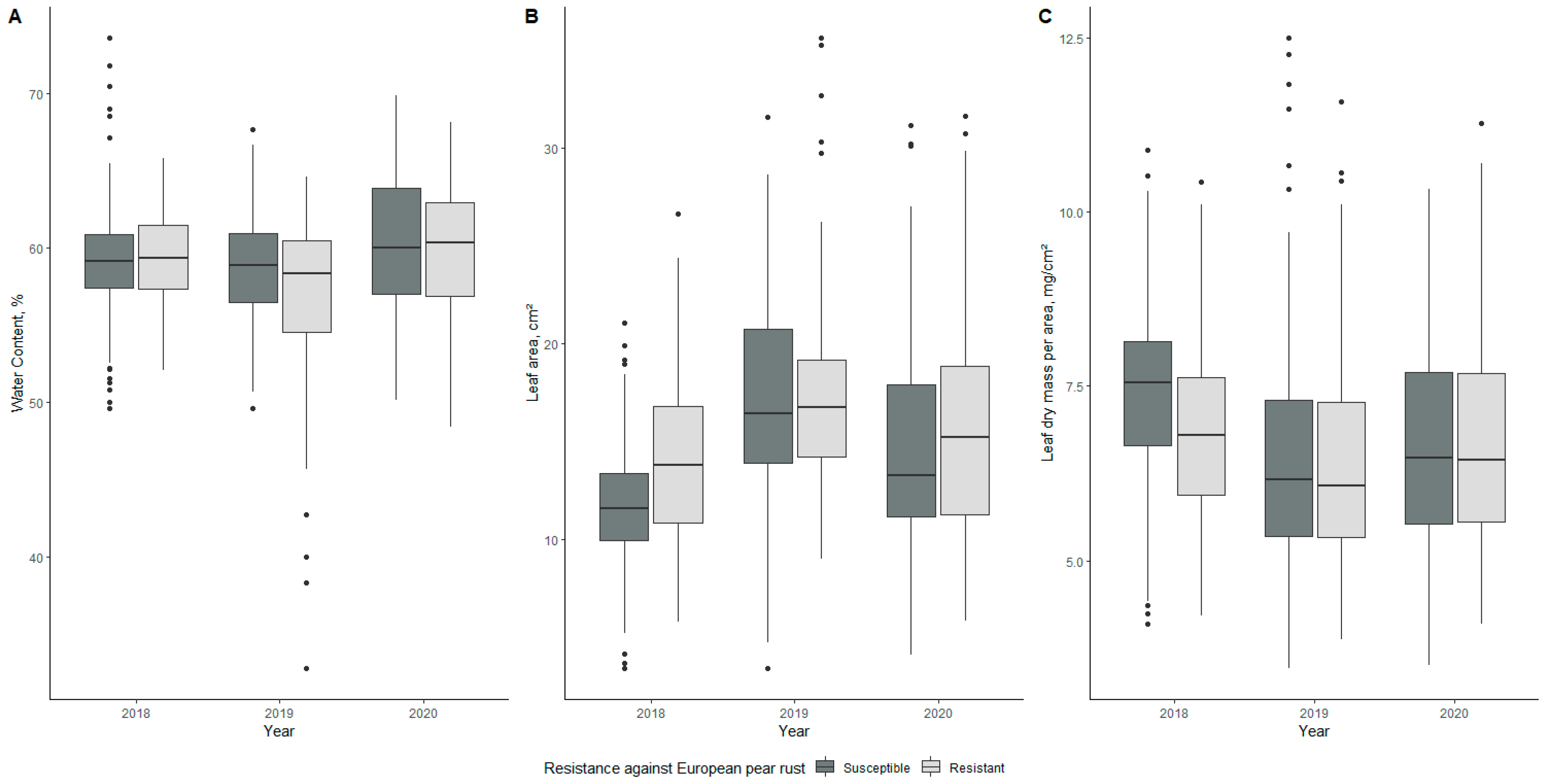
2.2.1. Leaf Water Content
2.2.2. Leaf Area
2.2.3. Leaf Mass per Area
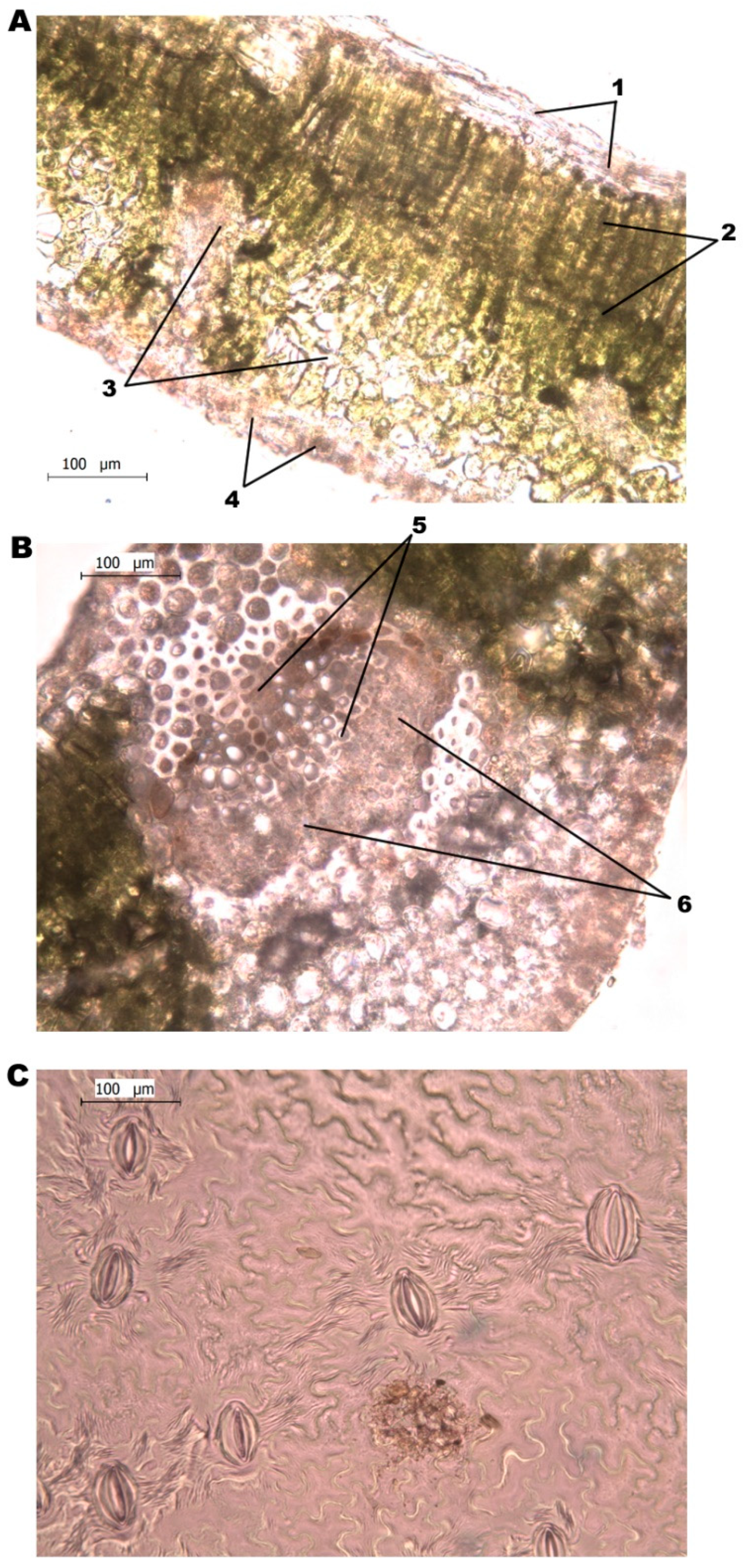
2.2.4. Leaf Epidermis
2.2.5. Mesophyll Tissue Thickness
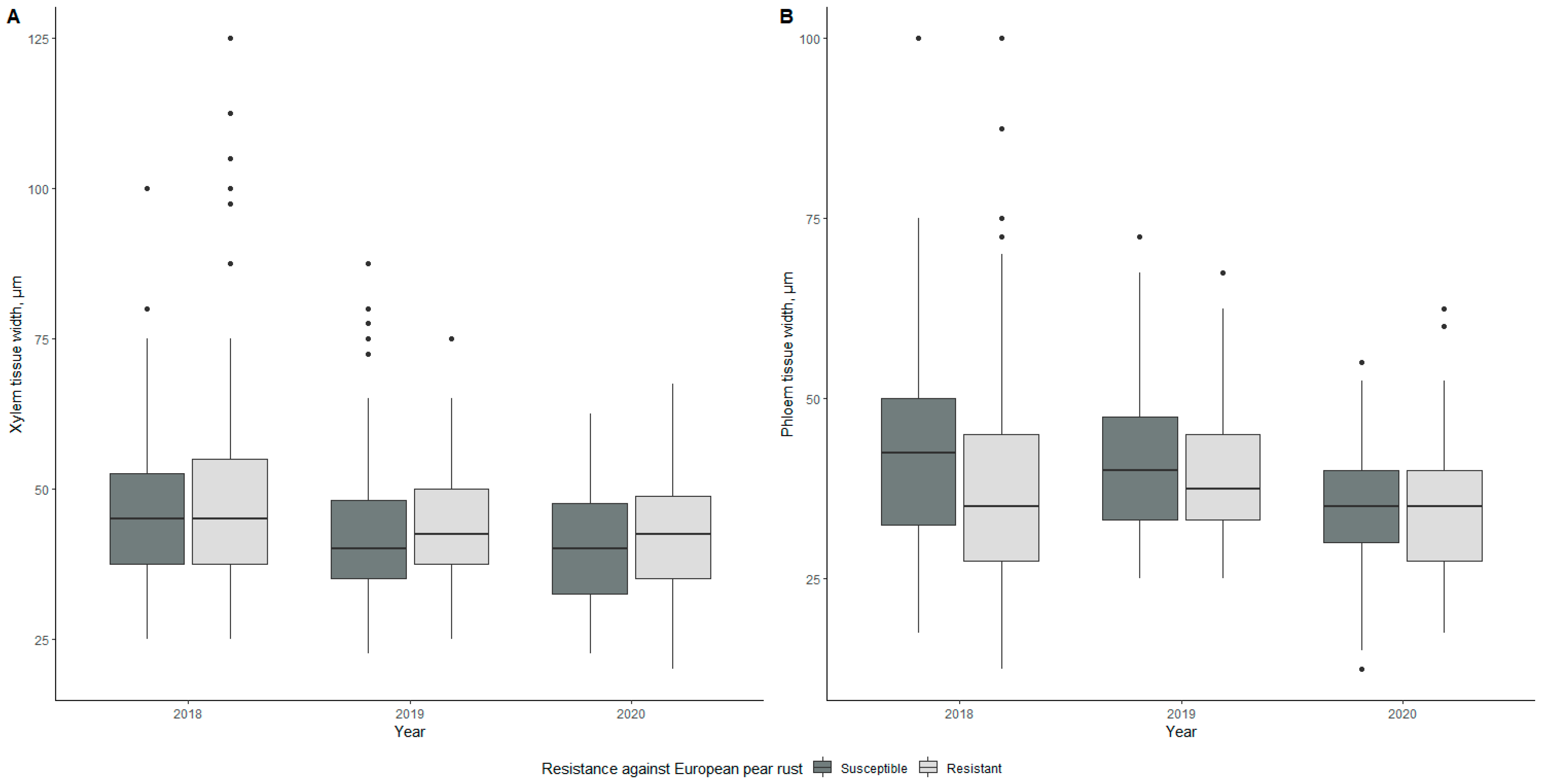
2.2.6. Vascular Tissue—Xylem and Phloem
2.2.7. Stomatal Density
2.2.8. Correlation Analysis
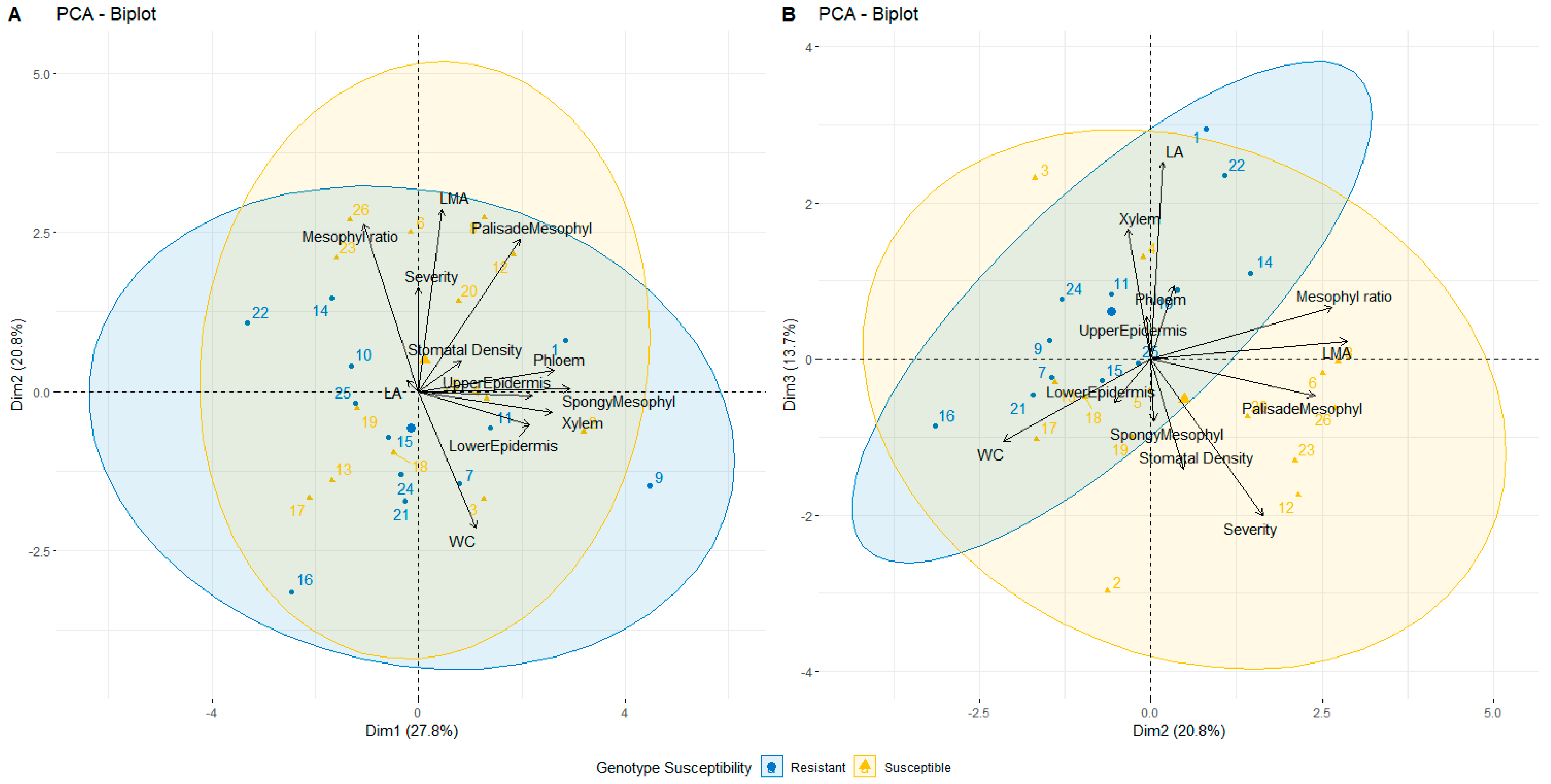
2.2.9. PCA Analysis
3. Discussion
3.1. Evaluation of G. sabinae Disease Severity
3.2. Leaf Morphology
4. Materials and Methods
4.1. Site of Experiment and Plant Material
4.2. Evaluation of G. sabinae Infection Severity
4.3. Measurement of Morphological Parameters
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Helfer, S. Overview of the Rust Fungi (Uredinales) Occurring on Rosaceae in Europe. Nova Hedwig. 2005, 81, 325–370. [Google Scholar] [CrossRef]
- Lāce, B. Gymnosporangium Species—An Important Issue of Plant Protection. Proc. Latv. Acad. Sci. Sect. B 2017, 71, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, X.; Wang, D.L. First Report of Rust Disease Caused by Puccinia Elymi on Leymus Chinensis in China. Plant Pathol. 2008, 57, 376. [Google Scholar] [CrossRef]
- Lowe, R.G.T.; Howlett, B.J. Indifferent, Affectionate, or Deceitful: Lifestyles and Secretomes of Fungi. PLoS Pathog. 2012, 8, e1002515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aime, M.C. Toward Resolving Family-Level Relationships in Rust Fungi (Uredinales). Mycoscience 2006, 47, 112–122. [Google Scholar] [CrossRef]
- Hau, B.; de Vallavieille-Pope, C. Wind-Dispersed Diseases. In The Epidemiology of Plant Diseases; Cooke, B., Jones, D., Kaye, B., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 387–416. [Google Scholar] [CrossRef]
- Lāce, B.; Bankina, B. Evaluation of European pear rust severity depending on agro-ecological factors. In Proceedings of the Annual 19th International Scientific Conference Research for Rural Development 2013, Jelgava, Latvia, 15–17 May 2013; pp. 6–12. Available online: http://www2.llu.lv/research_conf/Proceedings/19th_volume1.pdf (accessed on 25 March 2021).
- Juhasova, G.; Praslicka, J. Occurrence and Harmful Effects of Gymnosporangium Sabinae (Dicks.) Winter in Slovak Republic. Plant Prot. Sci. 2002, 38, 89–93. [Google Scholar] [CrossRef]
- Dervis, S.; Dixon, L.; Doǧanlar, M.; Rossman, A. Gall Production on Hawthorns Caused by Gymnosporangium spp. in Hatay Province, Turkey. Phytoparasitica 2010, 38, 391–400. [Google Scholar] [CrossRef]
- Bussotti, F.; Pollastrini, M. Evaluation of Leaf Features in Forest Trees: Methods, Techniques, Obtainable Information and Limits. Ecol. Indic. 2015, 52, 219–230. [Google Scholar] [CrossRef]
- Ritchie, S.W.; Nguyen, H.T.; Holaday, A.S. Leaf Water Content and Gas-Exchange Parameters of Two Wheat Genotypes Differing in Drought Resistance. Crop Sci. 1990, 30, 105–111. [Google Scholar] [CrossRef]
- Wang, M.; Sun, Y.; Sun, G.; Liu, X.; Zhai, L.; Shen, Q.; Guo, S. Water Balance Altered in Cucumber Plants Infected with Fusarium Oxysporum f. sp. cucumerinum. Sci. Rep. 2015, 5, 7722. [Google Scholar] [CrossRef] [Green Version]
- Castro-Díez, P.; Puyravaud, J.P.; Cornelissen, J.H.C. Leaf Structure and Anatomy as Related to Leaf Mass per Area Variation in Seedlings of a Wide Range of Woody Plant Species and Types. Oecologia 2000, 124, 476–486. [Google Scholar] [CrossRef]
- Cronin, J.P.; Welsh, M.E.; Dekkers, M.G.; Abercrombie, S.T.; Mitchell, C.E. Host Physiological Phenotype Explains Pathogen Reservoir Potential. Ecol. Lett. 2010, 13, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and Consequences of Variation in Leaf Mass per Area (LMA): A Meta-Analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H.; Potts, B.M.; Ratkowsky, D.A.; Pinkard, E.A.; Mohammed, C.L. Association of Eucalyptus Globulus Leaf Anatomy with Susceptibility to Teratosphaeria Leaf Disease. For. Pathol. 2018, 48. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The Formation and Function of Plant Cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafolla-Arellano, J.C.; Báez-Sañudo, R.; Tiznado-Hernández, M.E. The Cuticle as a Key Factor in the Quality of Horticultural Crops. Sci. Hortic. 2018, 232, 145–152. [Google Scholar] [CrossRef]
- Kachroo, A.; Robin, G.P. Systemic Signaling during Plant Defense. Curr. Opin. Plant Biol. 2013, 16, 527–533. [Google Scholar] [CrossRef]
- Semerdijeva, I.B.; Piperkova, N.G.; Zarkova, M.V.; Koleva-Valkova, L.H. Anatomical Changes in Peach Leaves Infected by Taphrina deformans (Berk.) Tul. Ecol. Balk. 2014, 5, 101–106. [Google Scholar]
- Varga, D.; Udvardy, L. Comparative Anatomical Study of Leaf Tissues of Scab Resistant and Susceptible Apple Cultivars. Int. J. Hortic. Sci. 2010, 16, 43–45. [Google Scholar] [CrossRef]
- Gonçalves-Zuliani, A.M.O.; Cardoso, K.A.K.; Belasque Junior, J.; Zanutto, C.A.; Hashiguti, H.T.; Bock, C.H.; Nakamura, C.V.; Nunes, W.M.D.C. Reaction of Detached Leaves from Different Varieties of Sweet Orange to Inoculation with Xanthomonas citri subsp. citri. Summa Phytopathol. 2016, 42, 125–133. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Ali, K.A.; Doss, S.G.; Das, N.K.; Aggarwal, R.K.; Bandopadhyay, T.K.; Sarkar, A.; Bajpai, A.K. Association of Leaf Micro-Morphological Characters with Powdery Mildew Resistance in Field-Grown Mulberry (Morus spp.) Germplasm. AoB Plants 2011, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; Chattopadhyay, S.; Saha, A.K.; Kumar, S.N. Association of Morphological and Biochemical Features of Mulberry Lines with Resistance to Bacterial Leaf Spot. Arch. Phytopathol. PFL 2014, 47, 2020–2032. [Google Scholar] [CrossRef]
- Melotto, M.; Zhang, L.; Oblessuc, P.R.; He, S.Y. Stomatal Defense a Decade Later. Plant Physiol. 2017, 174, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.L.; Li, B.H.; Zhang, Z.F.; Li, B.D.; Xu, X.M. Effect of Environmental Conditions on Germination and Survival of Teliospores and Basidiospores of the Pear Rust Fungus (Gymnosporangium asiaticum). Eur. J. Plant Pathol. 2006, 115, 341–350. [Google Scholar] [CrossRef]
- Klimata Portāls. Available online: https://klimats.meteo.lv/laika_apstaklu_raksturojums/2021/marts/ (accessed on 25 March 2021).
- Lāce, B. Impact of Agro-Ecological Factors on Development of European Pear Rust Caused by Gymnosporangium sabinae (Dicks.) g. Winter and Integrated Pest Management Posibilities for Its Control. Ph.D. Thesis, Latvia University of Agriculture, Jelgava, Latvia, 2016. [Google Scholar]
- Lace, B.; Lacis, G. Evaluation of Pear (Pyrus Communis, L.) Cultivars in Latvia. Hortic. Sci. 2015, 42, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Kellerhals, M.; Szalatnay, D.; Hunziker, K.; Duffy, B.; Nybom, H.; Ahmadi-Afzadi, M.; Höfer, M.; Richter, K.; Lateur, M. European Pome Fruit Genetic Resources Evaluated for Disease Resistance. Trees Struct. Funct. 2012, 26, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Thaxter, R. On Certain Cultures of Gymnosporangium, with Notes on Their Rœsteliæ. Proc. Am. Acad. Arts Sci. 1886, 22, 259–269. [Google Scholar] [CrossRef]
- Laajimi, N.O.; Boussadia, O.; Skhiri, F.H.; Teixeira da Silva, J.A.; Rezgui, S.; Hellali, R. Anatomical Adaptations in Vegetative Structures of Apricot Tree (Prunus armeniaca L.) cv. “Amor El Euch” Grown under Water Stress. FVCSB 2011, 5, 46–51. [Google Scholar]
- Anyia, A.O.; Herzog, H. Water-Use Efficiency, Leaf Area and Leaf Gas Exchange of Cowpeas under Mid-Season Drought. Eur. J. Agron. 2004, 20, 327–339. [Google Scholar] [CrossRef]
- Cui, T.; He, K.; Chang, L.; Zhang, X.; Xue, J.; Liu, J. QTL Mapping for Leaf Area in Maize (Zea mays L.) under Multi-Environments. J. Integr. Agr. 2017, 16, 800–808. [Google Scholar] [CrossRef]
- Gao, M.; van der Heijden, G.W.A.M.; Vos, J.; Eveleens, B.A.; Marcelis, L.F.M. Estimation of Leaf Area for Large Scale Phenotyping and Modeling of Rose Genotypes. Sci. Hortic. 2012, 138, 227–234. [Google Scholar] [CrossRef]
- Smith, A.H.; Gill, W.M.; Pinkard, E.A.; Mohammed, C.L. Anatomical and Histochemical Defence Responses Induced in Juvenile Leaves of Eucalyptus globulus and Eucalyptus nitens by Mycosphaerella Infection. For. Pathol. 2007, 37, 361–373. [Google Scholar] [CrossRef]
- Szwacka, M.; Tykarska, T.; Wisniewska, A.; Kuras, M.; Bilski, H.; Malepszy, S. Leaf Morphology and Anatomy of Transgenic Cucumber Lines Tolerant to Downy Mildew. Biol. Plant. 2009, 53, 697–701. [Google Scholar] [CrossRef]
- Kim, J.S.; Sagaram, U.S.; Burns, J.K.; Li, J.L.; Wang, N. Response of Sweet Orange (Citrus sinensis) to ‘Candidatus Liberibacter asiaticus’ Infection: Microscopy and Microarray Analyses. Phytopathology 2009, 99, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Yadeta, K.A.; Thomma, B.P.H.J. The Xylem as Battleground for Plant Hosts and Vascular Wilt Pathogens. Front. Plant Sci. 2013, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bendix, C.; Lewis, J.D. The Enemy within: Phloem-Limited Pathogens. Mol. Plant Pathol. 2018, 19, 238–254. [Google Scholar] [CrossRef] [Green Version]
- Gold, R.E.; Mendgen, K. Rust Basidiospore Germlings and Disease Initiation. In The Fungal Spore and Disease Initiation in Plants and Animals; Cole, G.T., Hoch, H.C., Eds.; Springer: Boston, MA, USA, 1991; pp. 67–99. [Google Scholar] [CrossRef] [Green Version]
- Hauser, P.M. Genomic Insights into the Fungal Pathogens of the Genus Pneumocystis: Obligate Biotrophs of Humans and Other Mammals. PLoS Pathog. 2014, 10, e1004425. [Google Scholar] [CrossRef] [Green Version]
- Bowen, J.K.; Mesarich, C.H.; Bus, V.G.M.; Beresford, R.M.; Plummer, K.M.; Templeton, M.D. Venturia inaequalis: The Causal Agent of Apple Scab. Mol. Plant Pathol. 2011, 12, 105–122. [Google Scholar] [CrossRef]
- Karpinets, T.V.; Park, B.H.; Syed, M.H.; Klotz, M.G.; Uberbacher, E.C. Metabolic Environments and Genomic Features Associated with Pathogenic and Mutualistic Interactions between Bacteria and Plants. Mol. Plant Microbe Interact. 2014, 27, 664–677. [Google Scholar] [CrossRef] [Green Version]
- Horsfall, J.G.; Barratt, R.W. An Improved Grading System for Measuring Plant Diseases. Phytopathology 1945, 35, 655. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 25 March 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean Values for the Morphological Parameters by Year of Measurement | |||||
|---|---|---|---|---|---|---|
| 2018 | 2019 | 2020 | ||||
| Resistant | Susceptible | Resistant | Susceptible | Resistant | Susceptible | |
| Leaf water content (WC), % | 59.32 ± 2.15 | 59.25 ± 3.57 | 57.00 ± 3.56 | 58.84 ± 2.41 | 59.59 ± 3.54 | 60.39 ± 2.84 |
| Leaf area (LA), cm2 | 14.16 ± 3.66 | 11.93 ± 2.85 | 17.42 ± 3.41 | 16.79 ± 4.49 | 15.54 ± 4.48 | 14.49 ± 4.59 |
| Leaf dry mass per area (LDMA), mg/cm2 | 6.80 ± 0.84 | 7.42 ± 1.04 | 6.40 ± 1.19 | 6.52 ± 1.47 | 6.73 ± 1.35 | 6.68 ± 1.25 |
| Upper epidermis, µm | 16.57 ± 2.10 | 15.89 ± 2.29 | 13.57 ± 1.35 | 14.10 ± 1.56 | 13.44 ± 1.50 | 12.43 ± 1.27 |
| Lower epidermis, µm | 12.47 ± 1.85 | 12.25 ± 1.31 | 11.77 ± 1.17 | 11.36 ± 0.89 | 10.42 ± 0.72 | 10.67 ± 0.63 |
| Palisade mesophyll, µm | 76.88 ± 6.08 | 84.45 ± 8.16 | 86.79 ± 7.61 | 92.63 ± 9.86 | 68.60 ± 7.66 | 69.77 ± 6.73 |
| Spongy mesophyll, µm | 93.47 ± 8.96 | 96.76 ± 8.46 | 82.40 ± 12.58 | 83.74 ± 13.03 | 80.25 ± 8.35 | 78.95 ± 7.40 |
| Palisade/spongy mesophyll ratio | 0.95 ± 0.08 | 1.03 ± 0.08 | 0.94 ± 0.07 | 0.98 ± 0.11 | 0.87 ± 0.10 | 0.91 ± 0.10 |
| Xylem, µm | 47.98 ± 13.91 | 47.22 ± 10.21 | 44.23 ± 6.42 | 42.60 ± 7.57 | 42.55 ± 5.83 | 40.29 ± 5.50 |
| Phloem, µm | 37.72 ± 11.12 | 42.78 ± 9.13 | 40.16 ± 5.38 | 41.27 ± 5.39 | 35.08 ± 5.60 | 34.92 ± 3.85 |
| Stomata density, stomata/mm2 | 126 ± 17.06 | 126 ± 22.35 | 107 ± 17.40 | 113 ± 23.22 | 106 ± 17.09 | 109 ± 20.69 |
| Water Content | Leaf Area | Leaf Mass Per Area | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | |||||||||
| 2018 | 2019 | 2020 | 2018 | 2019 | 2020 | 2018 | 2019 | 2020 | |
| 1-3-2v | 60.94 | 60.35 | 62.56 | 12.62 | 25.11 | 13.46 | 5.76 | 5.57 | 5.20 |
| 1-5-2v | 60.73 | 57.78 | 57.80 | 16.86 | 17.50 | 20.75 | 6.99 | 6.75 | 6.39 |
| 1-6-2v | 61.40 | 56.01 | 64.80 | 16.28 | 20.19 | 11.22 | 7.03 | 6.66 | 5.77 |
| 1-7-2v | 59.74 | 54.66 | 55.99 | 14.65 | 19.92 | 11.82 | 6.74 | 7.75 | 8.17 |
| 1-10-2v | 68.71 | 62.85 | 59.46 | 5.12 | 5.59 | 5.13 | 5.50 | 5.15 | 7.46 |
| 2-9-2v | 58.46 | 54.74 | 55.52 | 10.78 | 18.34 | 15.02 | 8.05 | 5.04 | 8.81 |
| 2-10-2v | 58.74 | 58.35 | 58.48 | 11.55 | 17.21 | 12.73 | 7.96 | 7.72 | 8.97 |
| 4-1-1v | 56.48 | 58.68 | 61.17 | 13.84 | 16.32 | 16.92 | 7.20 | 5.42 | 5.93 |
| 4-4-1v | 58.30 | 62.49 | 62.27 | 11.80 | 15.16 | 16.56 | 7.37 | 5.50 | 5.75 |
| 4-7-1v | 61.05 | 59.42 | 62.93 | 11.92 | 22.19 | 16.41 | 7.22 | 5.61 | 6.44 |
| 4-8-1v | 60.14 | 60.32 | 65.10 | 9.45 | 14.16 | 10.27 | 7.92 | 5.93 | 4.96 |
| 4-9-1v | 54.71 | 58.42 | 60.08 | 11.38 | 14.63 | 10.11 | 9.74 | 5.69 | 7.44 |
| 5-3-1v | 57.26 | 60.67 | 60.09 | 10.67 | 16.35 | 22.69 | 8.27 | 10.58 | 6.59 |
| 5-7-1v | 52.87 | 58.99 | 59.13 | 10.10 | 12.38 | 19.81 | 8.18 | 7.89 | 5.63 |
| 1-1-2v | 61.95 | 56.43 | 60.33 | 19.75 | 26.09 | 20.74 | 5.69 | 8.88 | 8.11 |
| 2-1-2v | 60.68 | 57.17 | 58.52 | 9.12 | 15.19 | 10.11 | 6.61 | 6.34 | 7.21 |
| 2-2-2v | 61.13 | 59.06 | 62.94 | 15.70 | 19.43 | 13.41 | 8.29 | 5.34 | 6.48 |
| 2-4-2v | 60.10 | 47.75 | 57.44 | 14.32 | 16.04 | 15.30 | 6.44 | 6.60 | 6.46 |
| 2-8-2v | 63.01 | 52.35 | 61.86 | 20.23 | 17.09 | 16.63 | 6.03 | 6.54 | 7.45 |
| 4-2-1v | 57.43 | 59.10 | 60.41 | 14.23 | 13.25 | 11.75 | 8.18 | 5.77 | 6.57 |
| 4-3-1v | 59.36 | 60.22 | 62.77 | 14.17 | 16.39 | 15.26 | 5.65 | 4.70 | 5.42 |
| 4-10-1v | 56.17 | 55.55 | 52.14 | 11.45 | 15.79 | 15.14 | 7.00 | 5.75 | 9.69 |
| 5-1-1v | 59.42 | 59.78 | 63.77 | 10.44 | 16.97 | 12.83 | 7.66 | 5.37 | 4.84 |
| 5-2-1v | 56.33 | 57.30 | 54.66 | 16.37 | 21.35 | 24.30 | 6.89 | 5.70 | 7.82 |
| 5-4-1v | 56.74 | 60.62 | 63.22 | 15.95 | 18.04 | 21.84 | 6.88 | 7.96 | 5.03 |
| 5-5-1v | 59.52 | 58.67 | 57.00 | 8.16 | 13.40 | 9.22 | 6.35 | 7.82 | 5.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kārkliņa, K.; Lācis, G.; Lāce, B. Differences in Leaf Morphological Parameters of Pear (Pyrus communis L.) Based on Their Susceptibility to European Pear Rust Caused by Gymnosporangium sabinae (Dicks.) Oerst. Plants 2021, 10, 1024. https://doi.org/10.3390/plants10051024
Kārkliņa K, Lācis G, Lāce B. Differences in Leaf Morphological Parameters of Pear (Pyrus communis L.) Based on Their Susceptibility to European Pear Rust Caused by Gymnosporangium sabinae (Dicks.) Oerst. Plants. 2021; 10(5):1024. https://doi.org/10.3390/plants10051024
Chicago/Turabian StyleKārkliņa, Katrīna, Gunārs Lācis, and Baiba Lāce. 2021. "Differences in Leaf Morphological Parameters of Pear (Pyrus communis L.) Based on Their Susceptibility to European Pear Rust Caused by Gymnosporangium sabinae (Dicks.) Oerst." Plants 10, no. 5: 1024. https://doi.org/10.3390/plants10051024