
Inhibitory Effects of Brassicaceae Cover Crop on Ambrosia artemisiifolia Germination and Early Growth

 , , and
, , and
Abstract
:1. Introduction
2. Results
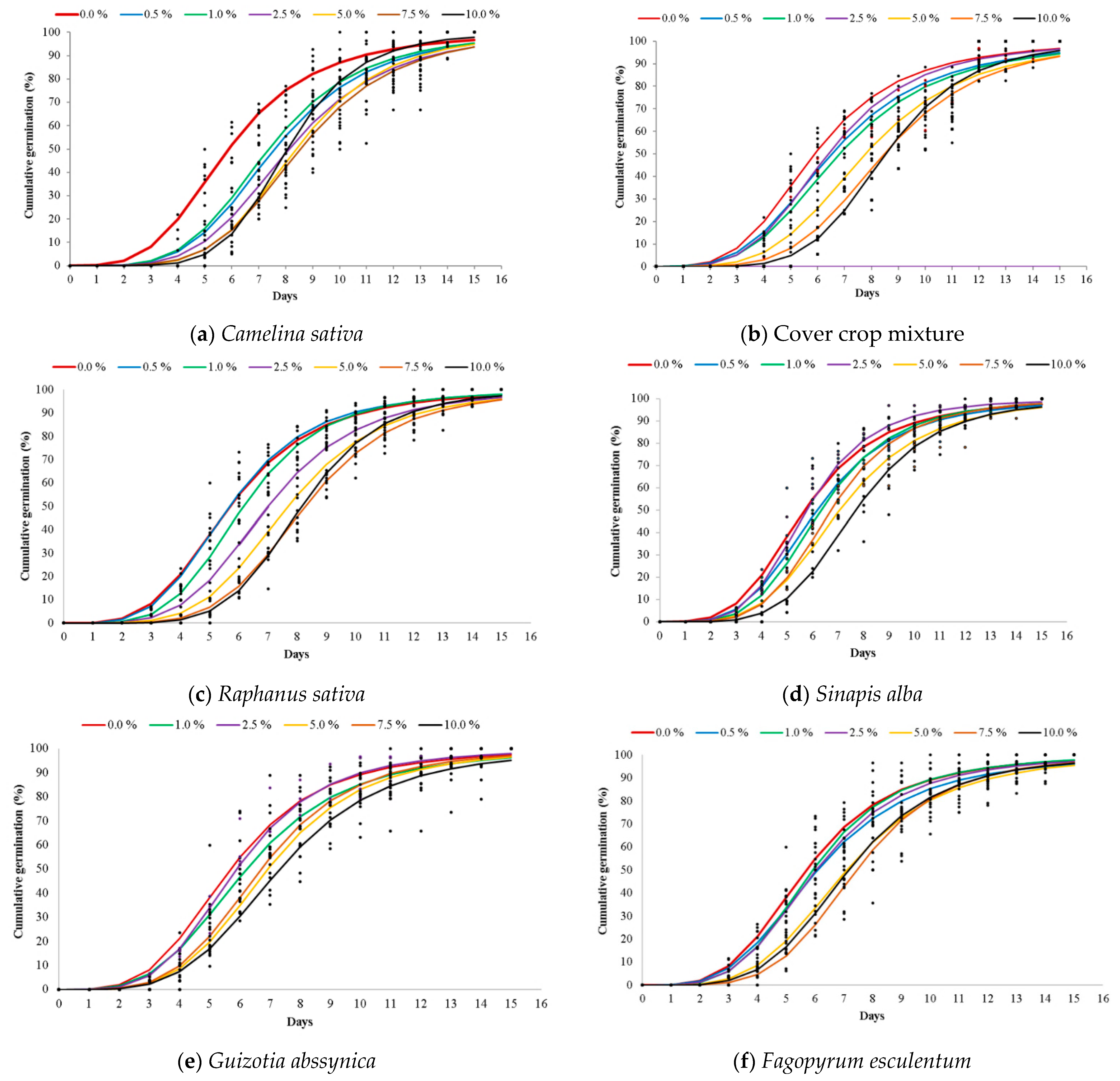
2.1. Allelopathic Effects of Aqueous Extracts on Germination and Early Growth of A. artemisiifolia
2.2. Allelopathic Effects of Aqueous Extracts on A. artemisiifolia Germination Rate
2.3. Allelopathic Effects of Aqueous Extracts of Individual Parts of C. sativa on Germination and Early Growth
2.4. Chemical Analyses of Aqueous CC Extracts and Correlation with Allelopathic Effects on A. artemisiifolia Radicle Length
3. Discussion
4. Materials and Methods
4.1. Plant Material (Sources)
4.2. Seed Bioassay
4.3. Chemical Analyses of Aqueous Extracts
4.3.1. Standards and Reagents
4.3.2. Liquid Chromatography-Tandem Mass Spectrometry
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
| Compound | Precursor m/z | Product m/z | (Fr) (V) | CE (V) | tR (min) |
|---|---|---|---|---|---|
| Gallic acid | 169.0 | 125.0 | 90 | 10 | 3.67 |
| Protocatechuic acid | 153.0 | 109.0 | 105 | 9 | 6.27 |
| Chlorogenic acid | 353.0 | 191.0 | 100 | 10 | 8.54 |
| Vanillic acid | 167.0 | 108.0 | 100 | 15 | 9.14 |
| Caffeic acid | 179.0 | 135.0 | 100 | 10 | 9.14 |
| Syringic acid | 197.0 | 182.0 | 90 | 7 | 9.71 |
| Vanilin | 151.1 151.1 | 136.0 90.0 | 100 100 | 17 21 | 10.02 |
| p-coumaric acid | 163.0 | 119.0 | 90 | 9 | 10.68 |
| Sinapinic acid | 223.0 | 193.0 | 100 | 17 | 11.13 |
| Ferulic acid | 193.0 | 134.0 | 90 | 11 | 11.14 |
| Rutin | 609.0 | 300.0 | 135 | 42 | 11.96 |
| Quercetin | 301.0 | 151.0 | 130 | 15 | 13.68 |
| Quercitin | 447.0 | 300.0 | 190 | 27 | 12.07 |
| Kaempferol | 285.0 | 285.0 | 130 | 0 | 14.25 |
| Cover Crop (s) | Parameters | ||
|---|---|---|---|
| Germination (%) | Shoot Length (cm) | Radical Length (cm) | |
| Control | 35.0 ± 8.08 a | 3.06 ± 0.89 a | 2.51 ± 0.42 a |
| CCmix | 23.0 ± 3.06 b | 0.90 ± 0.05 c | 1.05 ± 0.11 c |
| S. alba | 8.0 ± 1.90 c | 0.63 ± 0.04 c,d | 0.10 ± 0.00 d |
| R. sativus | 1.0 ± 0.06 d | 0.00 ± 0.00 e | 0.27 ± 0.08 d |
| F. esculentum | 12.0 ± 1.32 c | 2.29 ± 0.09 b | 1.98 ± 0.01 b |
| C. sativa | 9.0 ± 0.50 c | 0.31 ± 0.04 d | 0.08 ± 0.03 d |
| G. abssynica | 9.0 ± 1.72 c | 0.92 ± 0.08 c | 0.82 ± 0.04 c |

References
- Essl, F.; Biró, K.; Brandes, D.; Broennimann, O.; Bullock, J.M.; Chapman, D.S.; Karrer, G. Biological flora of the British Isles: Ambrosia artemisiifolia. J. Ecol. 2015, 103, 1069–1098. [Google Scholar] [CrossRef] [Green Version]
- Medzihradszky, Z.; Járai-Komlódi, M. I came from America–My name is Ambrosia–Some feature of the ragweed. In Proceedings of the 9th EWRS Symposium, Budapest, Hungary, 10–12 July 1995; pp. 57–63. [Google Scholar]
- Šikoparija, B.; Radišić, P.; Pejak, T.; Šimić, S. Airborne grass and ragweed pollen in the southern Panonnian Valley-consideration of rural and urban environment. Ann. Agric. Environ. Med. 2006, 13, 263–266. [Google Scholar] [PubMed]
- Lommen, S.T.E.; Caspar, A.H.; Jongejans, E.; Chauvel, B.; Leitsch-Vitalos, M.; Aleksanyan, A.; Scepanovic, M.; Onen, H.; Tokarska-Guzik, B.; Anastasiu, P.; et al. Explaining variability in the production of seed and allergenic pollen by invasive Ambrosia artemisiifolia across Europe. Biol. Invasions 2018, 5, 1475–1491. [Google Scholar] [CrossRef] [Green Version]
- Šarić, T.; Ostojić, Z.; Stefanović, L.; Deneva Milanova, S.; Kazinczi, G.; Tyšer, L. The changes of the composition of weed flora in Southeastern and Central Europe as affected by cropping practices. Herbologia 2011, 12, 8–12. [Google Scholar]
- Galzina, N.; Barić, K.; Šćepanović, M.; Goršić, M.; Ostojić, Z. Distribution of Invasive Weed Ambrosia artemisiifolia L. in Croatia. Agric. Conspec. Sci. 2010, 75, 75–81. [Google Scholar]
- Heap, I. The international Survey of Herbicide Resistance Weeds. 2021. Available online: www.weedscience.org (accessed on 17 January 2021).
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest Manag. Sci. 2012, 68, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scavo, A.; Mauromicale, G. Integrated Weed Management in Herbaceous Field Crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Kunz, C.; Strum, D.; Varnholt, D.; Walker, F.; Gerhards, R. Allelopathic effects and weed suppressive ability of cover crops. Plant Soil Environ. 2016, 62, 60–66. [Google Scholar]
- Triplett, G.B.; Warren, A.D. No-Tillage Crop Production: A Revolution in Agriculture! Agron. J. 2008. [CrossRef]
- Scavo, A.; Restuccia, A.; Lombardo, S.; Fontanazza, S.; Abbate, C.; Pandino, G.; Anastasi, U.; Onofri, A.; Mauromicale, G. Improving soil health, weed management and nitrogen dynamics by Trifolium subterraneum cover cropping. Agron. Sustain. Dev. 2020, 40, 1–12. [Google Scholar] [CrossRef]
- Finney, D.M.; Creamer, N.G. Weed Management on Organic Farms; The organic Production Publication Series; CEFS: Brussels, Belgium, 2008; pp. 1–34. [Google Scholar]
- Macias, F.A.; Molinillo, M.G.; Oliveros-Bastidas, A.; Marin, D.; Chinchilla, D. Allelopathy. A natural strategy for weed control. Comm. Appl. Biol. Sci. 2004, 69, 13–23. [Google Scholar]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop. Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Farooq, M.; Jabran, K.; Cheema, Z.A.; Wahid, A.; Siddique, K.H.M. The role of allelopathy in agricultural pest management. Pest Manag. Sci. 2011, 67, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Nichols, V.; Verhulst Cox, R.; Govaers, B. Weed dynamics in conservation agriculture principles: A review. Field Crop Res. 2015, 183, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Gentili, R.; Gilardelli, F.; Ciappetta, S.; Ghiani, A.; Citterio, S. Inducing competition: Intensive grassland seeding to control Ambrosia artemisiifolia. Weed Res. 2015, 55, 278–288. [Google Scholar] [CrossRef]
- Hiridate, S. Isolation strategies for finding bioacitve compounds: Specific activirty vs. total activity. Chem. Soc. Symp. Ser. 2006, 927, 113–126. [Google Scholar]
- Sherif, E.A.; Hegazy, A.K.; Gomaa, N.H.; Hassan, M.O. Allelopathic effect of Black Mustard Tissues and Root Exudates on some crops and weeds. Planta Daninha Vixoa MG 2013, 31, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Dhima, K.V.I.; Vasilakoglou, T.D.; Gatsis, E. Panou-Philotheou, Eleftherohorinos, I. Effects of aromatic plants incorporated as green manure on weed and maize development. Field Crops Res. 2009, 110, 235–241. [Google Scholar] [CrossRef]
- Brijačak, E.; Košćak, L.; Šoštarčić, V.; Kljak, K.; Šćepanović, M. Sensitivity of yellow foxtail (Setaria glauca L) and barnyardgrass (Echninochloa crus-galli L.) to aqueous extracts or dry biomass of cover crop. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Pandino, G.; Onofri, A.; Mauromicale, G. Allelopathic effect of Cynara cardunculus L. leaf aqueous extracts on seed germination of some Medirettanean weed species. Ital. J. Agron. 2018, 3, 119–125. [Google Scholar]
- Tessio, F.; Ferrero, A. Allelopathy, a chance for sustainable weed management. Int. J. Sust. Dev. World 2010, 17, 377–389. [Google Scholar] [CrossRef]
- Gomaa, N.H.; Hassan, M.O.; Fahmy, G.M.; Gonzalez, L.; Hammouda, O.; Atteya, A.M. Allelopathic effect of Sonchus oleraceus L. on the germination and seedling growth of crop and weed species. Acta Bot. Bras. 2014, 25, 408–416. [Google Scholar] [CrossRef]
- Francis, A.; Warwick, S.I. The biology of Canadian weeds. 142. Camelina alyssum (Mill.) Thell.; C. microcarpa Andrz. Ex DC.; C. sativa (L.) Crantz. Can. J. Plant Sci. 2009, 89, 791–810. [Google Scholar]
- Kollmann, J.; Bassin, S. Effects of management on seed predation in wildflowerstrips in northern Switzerland. Agric. Ecocsyst. Environ. 2001, 83, 285–296. [Google Scholar] [CrossRef]
- Walsh, K.D.; Sanderson, D.; Hall, L.M.; Mugo, S.; Hills, M.J. Allelopathic effects of camelina (Camelina sativa) and canola (Brassica napus) on wild oat, flax and radish. Allelopath. J. 2014, 33, 83–96. [Google Scholar]
- Hiag, T. Allelochemicals in plants. In Allelopathy in Sustainable Agriculture and Forestry; Zeng, R.S., Mallik, A.U., Luo, S.S., Eds.; Springer: New York, NY, USA, 2008; pp. 63–104. [Google Scholar]
- Quasem, J.G.; Foy, C.L. Weed allelopathy, its ecological impacts and future prospects: A review. Weed Biol. Manag. 2001, 4, 43–199. [Google Scholar] [CrossRef]
- Chou, C.H. Introduction of allelopathy. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., Gonzales, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 1–9. [Google Scholar]
- Stupnicka-Rodzynkiewicz, E.; Dabkowska, T.; Stoklosa, A. The effect of selected phenolic compounds on the initial growth of four weed species. J. Plant Dis. Prot. 2006, 20, 479–486. [Google Scholar]
- Fahmy, G.M.; Al-Sawaf, N.A.; Turki, H.; Ali, H.I. Allelopathic potential of Pluchea dioscordis (L.) DC. J. Appl. Sci. Res. 2012, 8, 3129–3142. [Google Scholar]
- Li, Z.H.; Wang, Q.; Ruan, X.; De Pan, C.; Jiang, D.-A. Phenolics and Plant Allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [Green Version]
- Turk, M.A.; Shatnawi, M.K.; Tawaha, A.M. Inhibitory effects of aqueous extracts of black mustard on germination and growth of alfalfa. Weed Biol. Manag. 2003, 3, 37–40. [Google Scholar] [CrossRef]
- Iqbal, Z.; Hiradate, S.; Noda, A.; Isojima, S.-I.; Fujii, Y. Allelopathic activity of buckwheat: Isolation and characterization of phenolics. Weed Sci. 2003, 51, 657–662. [Google Scholar] [CrossRef]
- Einhellig, F.A. Mode of allelochemical action of phenolic compounds. In Allelopathy: Chemistry and Mode of Action of Allelochemicals; Macias, F.A., Galindo, J.C.G., Molinillo Jose, M.G., Eds.; CRC Press, LCC: London, UK, 2004. [Google Scholar]
- Batish, D.R.; Kaur, S.; Singh, H.P.; Kohli, R.K. Nature of interference potential of leaf debris of Ageratum conyzoides. Plant Growth Regul. 2009, 57, 137–144. [Google Scholar] [CrossRef]
- Liebman, M.; Davis, A. Integration of soil, crop and weed management in low-external-input farming systems. Weed Res. 2000, 40, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Šoštarčić, V.; Masin, R.; Turčinov, M.; Carin, N.; Šćepanović, M. Morfološka i funkcionalna intrapopulacijska varijabilnost sjemena korovne vrste Ambrosia artemisiifolia L. J. Cent. Eur. Agric. 2020, 21, 366–378. [Google Scholar] [CrossRef]
- Haramoto, E.R.; Gallandt, E.R. Brassica cover cropping: I. Effect on weed and crop establishment. Weed Sci. 2005, 53, 695–701. [Google Scholar] [CrossRef]
- Muscolo, A.; Panuccio, M.; Sidari, M. The effect of phenols on respiratory enzymes in seed germination. Plant Growth Regul. 2001, 35, 31–35. [Google Scholar] [CrossRef]
- Sobiech, L.; Grzanka, M.; Popowska, D.K.; Rdzikowska, D.; Sobiech, Ł.; Grzanka, M.; Kurasiak-Popowska, D.; Radzikowska, D. Phytotoxic Effect of Herbicides on Various Camelina [Camelina sativa (L.) Crantz] Genotypes and Plant Chlorophyll Fluorescence. Agriculture 2020, 10, 185. [Google Scholar] [CrossRef]
- Chaves, N.; Sosa, T.; Escudero, J.C. Plant growth inhibiting flavonoids in exudate of Cistus ladanifer and in associatedsoils. J. Chem. Ecol. 2001, 27, 623–631. [Google Scholar] [CrossRef]
- ISTA. International Rules for Seed Testing; International Association for Seed Testing: Zurich, Switzerland, 2008. [Google Scholar]
- Einhellig, F.A.; Rasmussen, J.A. Effects of three phenolic acids on chlorophyll content and growth of soybean and grain sorghum seedlings. J. Chem. Ecol. 1979, 5, 815–824. [Google Scholar] [CrossRef]
- Krogmeir, M.J.; Bremner, J.M. Effects of phenolic acids on seed germination and seedling growth in soil. Biol. Fertil. Soils 1989, 8, 116–122. [Google Scholar] [CrossRef]
| Extract Concentration (w/v) | Cover Crop (s) | |||||
|---|---|---|---|---|---|---|
| All Crops Mixed (CCMIX) | C. sativa | F. esculentum | G. abyssinica | R. sativus | S. alba | |
| Control | 75.00 ± 2.35 A–F ab | |||||
| 0.5 | 68.13 ± 2.91 C–F ab | 67.50 ± 2.08 C–F ab | 76.88 ± 2.81 A–C a | 79.38 ± 1.25 A–C a | 77.50 ± 2.54 A–C ab | 83.33 ± 3.12 A a |
| 1.0 | 76.88 ± 2.75 D–G a | 56.88 ± 1.66 E–H bc | 78.13 ± 2.27 A–C a | 76.25 ± 2.99 A–C a | 78.13 ± 2.73 A–C a | 81.67 ± 2.24 AB a |
| 2.5 | 63.75 ± 1.92 D–G ab | 53.75 ± 1.51 G–I bc | 76.88 ± 2.54 A–C a | 74.38 ± 2.39 A–D a | 72.50 ± 2.40 A–D ab | 72.50 ± 2.40 A–D ab |
| 5.0 | 55.00 ± 1.42 G–I bc | 51.25 ± 1.44 HI c | 73.75 ± 2.10 A–D ab | 72.50 ± 2.40 A–D a | 71.25 ± 1.44 A–D ab | 69.17 ± 1.61 C–E b |
| 7.5 | 55.00 ± 1.54 G–I ab | 48.75 ± 1.07 HI c | 69.38 ± 2.15 B–D ab | 71.88 ± 2.68 A–D a | 68.13 ± 1.53 C–F ab | 67.50 ± 1.61 C–F bc |
| 10.0 | 45.63 ± 1.11 HI c | 43.13 ± 1.04 I c | 63.75 ± 1.44 D–G b | 71.88 ± 2.27 A–D a | 63.75 ± 1.33 D–G b | 55.83 ± 1.24 F–H c |
| Parameter | Concentration (w/v) | Cover Crop (s) | |||||
|---|---|---|---|---|---|---|---|
| All Crops Mixed Together (CCMIX) | C. sativa | F. esculentum | G. abssynica | R. sativus | S. alba | ||
| Shoot length (cm) | Control | 1.90 ± 0.52 D–N a–c | |||||
| 0.5 | 2.34 ± 0.17 AB a | 1.54 ± 0.29 C d | 1.92 ± 0.67 G–K a | 2.09 ± 0.86 E–H a | 2.50 ± 0.77 CD a | 1.44 ± 0.35 L–O bc | |
| 1.0 | 2.26 ± 0.20 A a | 1.36 ± 0.24 C d | 1.81 ± 0.70 G–L a | 2.05 ± 0.62 F–I a | 2.36 ± 0.74 C–F a | 1.59 ± 0.32 K–O b | |
| 2.5 | 2.23 ± 0.22 B a | 1.39 ± 0.11 C d | 1.68 ± 0.54 I–M a | 2.07 ± 0.62 F–I a | 2.00 ± 0.62 F–J b | 1.53 ± 0.35 J–N b | |
| 5.0 | 2.08 ± 0.22 C–E a | 1.10 ± 0.15 D–G d | 1.69 ± 0.49 H–M a | 1.99 ± 0.68 F–J a | 1.88 ± 0.34 G–K bc | 1.39 ± 0.27 M–P bc | |
| 7.5 | 1.99 ± 0.23 F–J ab | 0.91 ± 0.06 P–S e | 1.60 ± 0.43 J–N a | 1.97 ± 0.63 F–J a | 1.68 ± 0.32 I–M c | 1.15 ± 0.26 O–R cd | |
| 10.0 | 1.52 ± 0.19 M–R c | 0.87 ± 0.22 S e | 1.64 ± 0.50 J–N a | 1.85 ± 0.48 G–K a | 1.27 ± 0.22 N–R d | 0.98 ± 0.14 R–S d | |
| Radicle length (cm) | Control | 3.79 ± 0.42 D–K a–c | |||||
| 0.5 | 4.09 ± 0.25 L–O a | 2.69 ± 0.55 N–S c | 4.79 ± 0.73 A–D a | 4.93 ± 0.57 A–C a | 4.41 ± 0.92 AB a | 5.46 ± 1.09 A a | |
| 1.0 | 4.45 ± 0.46 L–O c | 2.76 ± 0.28 P–T c | 4.62 ± 0.89 B–F a | 4.63 ± 0.61 B–F ab | 4.08 ± 0.91 A–D a | 4.73 ± 0.50 A–E ab | |
| 2.5 | 4.01 ± 0.39 L–O cd | 2.67 ± 0.26 P–T cd | 4.49 ± 0.68 C–F a | 4.79 ± 0.62 A–D ab | 4.05 ± 0.59 A–D a | 4.83 ± 0.72 A–D ab | |
| 5.0 | 2.48 ± 0.18 M–P d | 2.20 ± 0.23 ST d | 4.46 ± 0.86 D–G ab | 4.32 ± 0.33 D–H ab | 3.65 ± 0.37 G–J b | 3.86 ± 0.47 F–J bc | |
| 7.5 | 2.00 ± 0.40 M–R d | 1.01 ± 0.61 T e | 3.28 ± 0.58 I–K bc | 3.94 ± 0.39 E–I bc | 2.35 ± 0.17 L–N c | 3.61 ± 0.40 H–J c | |
| 10.0 | 1.32 ± 0.35 O–T e | 0.69 ± 0.28 T e | 2.27 ± 0.40 K–M c | 3.05 ± 0.21 J–L c | 1.23 ± 0.29 R–T d | 1.59 ± 0.53 N–T d | |
| Seedling fresh weight (g) | Control | 0.27 ± 0.05 A–K ab | |||||
| 0.5 | 0.40 ± 0.17 C–I a | 0.31 ± 0.04 B–G a | 0.30 ± 0.03 B–F a | 0.33 ± 0.07 A–D a | 0.32 ± 0.04 A–E a | 0.34 ± 0.04 A–C a | |
| 1.0 | 0.49 ± 0.11 A a | 0.23 ± 0.07 E–I a | 0.30 ± 0.01 B–F a | 0.35 ± 0.07 A–C a | 0.33 ± 0.04 A–E a | 0.36 ± 0.01 AB a | |
| 2.5 | 0.43 ± 0.13 A–C a | 0.22 ± 0.07 G–K a | 0.31 ± 0.06 A–F a | 0.33 ± 0.09 A–E a | 0.29 ± 0.06 B–F ab | 0.35 ± 0.04 A–C a | |
| 5.0 | 0.30 ± 0.16 I–K b | 0.20 ± 0.11 G–K c | 0.33 ± 0.04 A–E a | 0.32 ± 0.04 A–F a | 0.29 ± 0.04 B–F ab | 0.33 ± 0.06 A–D ab | |
| 7.5 | 0.27 ± 0.17 KLb c | 0.17 ± 0.01 M c | 0.32 ± 0.05 A–D a | 0.31 ± 0.03 A–F a | 0.23 ± 0.01 F–J bc | 0.31 ± 0.03 A–F ab | |
| 10.0 | 0.22 ± 0.15 J–L bc | 0.13 ± 0.09 LM c | 0.27 ± 0.03 B–H a | 0.28 ± 0.03 B–G a | 0.18 ± 0.02 H–K c | 0.25 ± 0.01 D–I c | |
| Aqueous Extract Concentration (w/v) | Cover Crop (s) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| All Crops Mixed Together (CCMIX) | C. sativa | F. esculentum | G. abssynica | R. sativus | S. alba | |||||||
| t10 | t50 | t10 | t50 | t10 | t50 | t10 | t50 | t10 | t50 | t10 | t50 | |
| Control | 3.156 PR ab | 5.520 T a | 3.205 PR ab | 5.882 P–T a–c | 3.245 PR ab | 5.717 R–T ab | 3.067 R a | 5.575 ST a | 3.152 PR ab | 5.615 ST a | 3.237 PR ab | 5.605 ST a |
| 0.5 | 3.487 O–R a–c | 6.514 I–R c–g | 4.614 E– I h–l | 7.597 C–G k–p | 3.390 O–R a–c | 6.120 M–T a–e | 3.595 N–R a–d | 8.210 K–S o–t | 3.296 PR ab | 5.692 R–T ab | 3.514 O–R a–c | 6.163 L–T a–e |
| 1.0 | 3.729 L–R b–f | 6.828 G–N e–j | 4.596 E–I h–l | 7.437 D–H i–n | 3.515 O–R ab | 5.983 O–T a–d | 3.634 M–R a–e | 6.400 K–S b–f | 3.785 K–P b–g | 6.165 L–T a–e | 4.003 I–O c–h | 6.380 K–Sb–f |
| 2.5 | 3.632 M–R a–e | 6.681 D–H d–h | 5.036 B–E l–o | 8.177 A–D n–s | 3.484 O–R a–c | 6.058 N–T a–d | 3.522 O–R a–c | 5.928 O–T a–c | 4.410 E–K g–l | 6.946 F–M f–k | 3.579 N –R a–d | 5.774 R–Ta–c |
| 5.0 | 4.522 E–I h–l | 7.787 A–E m–s | 5.607 AB op | 8.447 AB r–t | 4.286 G–M e–j | 7.171 E–K g–m | 4.223 H–N d–i | 6.954 F–L f–l | 4.908 C–G j–n | 7.695 B–F l–p | 4.211 H–N d–i | 7.076 E–K f–m |
| 7.5 | 5.323 A–D m–p | 8.473 AB st | 5.529 A–C n–p | 8.591 A t | 4.824 D– H i–m | 7.468 C–H j–o | 4.037 I–O c–h | 6.697 H–P d–i | 5.473 A–D m–p | 8.293 A–C p–t | 4.237 H– N d–i | 6.727 H–O d–j |
| 10.0 | 5.810 A p | 8.549 A t | 5.895 A p | 8.163 A–D n–t | 4.445 E– K g–l | 7.180 E–K g–m | 4.368 F–L f–k | 7.316 E–I h–m | 5.658 AB op | 8.179 A–D n–t | 5.024 B–F k–o | 7.718 B–F m–r |
| Parameters | Control | Camelina sativa Aqueous Extracts | ||||
|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Fruit | Whole Plant | ||
| Viable seed (%) | 95.2 ± 5.22 a | 95.3 ± 3.01 a | 93.3 ± 3.27 a | 95.2 ± 6.57 a | 92.8 ± 5.22 a | 90.7 ± 6.02 a |
| Germinated seed (%) | 73.6 ± 6.69 a | 58.7 ± 10.33 b | 63.3 ± 5.89 a,b | 55.2 ± 7.69 b | 18.4 ± 8.76 c | 54.0 ± 10.04 b |
| Shoot length (cm) | 1.43 ± 0.11 a | 0.65 ± 0.36 b | 1.25 ± 0.57 a | 0.66 ± 0.39 b | 0.61 ± 0.40 b | 0.52 ± 0.34 b |
| Radical length (cm) | 3.55 ± 0.70 a | 0.95 ± 0.47 b,c | 1.22 ± 0.69 b | 1.06 ± 0.46 b | 0.32 ± 0.13 d | 0.41 ± 0.09 c,d |
| Seedling fresh weight (g) | 0.37 ± 0.07 a | 0.16 ± 0.07 c | 0.25 ± 0.07 b | 0.14 ± 0.07 c | 0.09 ± 0.04 c | 0.12 ± 0.05 c |
| Chemical Class | Compound | Content of Phenolic Compounds (µg/g of Dry Extract) | |||||
|---|---|---|---|---|---|---|---|
| S. alba | F. esculentum | R. sativa | G. abssynica | C. sativa | CCMIX | ||
| Phenolic aldehydes | Vanillin | 19.0 ± 0.28 b | 13.3 ± 0.22 c | 19.0 ± 0.23 b | 13.8 ± 0.11 c | 44.8 ± 0.39 a | n.d. d |
| Bioflavonoids | Kaempferol | 46.5 ± 0.59 a | n.d. c | n.d. c | 40.0 ± 0.44 b | n.d. c | n.d. c |
| Flavonoid glycosides | Rutin | n.d. e | 1844.3 ± 1.48 a | 6.0 ± 0.09 d | 28.8 ± 0.25 c | 413.5 ± 0.86 b | 416.5 ± 0.87 b |
| Flavonols | Quercetin | 71.6 × 103 ± 21.18 d | 895.6 × 103 ± 10.90 a | 189.9 × 103 ± 25.15 b | 980.5 ± 35.65 e | 68.8 ± 0.75 f | 88.8 × 103 ± 5.25 c |
| Quercitin | 40.8 ± 0.86 c | 135.8 ± 1.11 a | 8.3 ± 0.05 e | 7.0 ± 0.09 e | 33.3 ± 0.48 d | 59.5 ± 0.60 b | |
| Hydroxybenzoic acids | Gallic acid | 65.5 ± 0.47 a | 32.3 ± 0.65 b | 18.5 ± 0.43 c | 17.5 ± 0.34 c | 17.5 ± 0.35 c | 26.3 ± 0.50 b |
| Protocatechuic acid | 55.5 ± 0.60 e | 386.3 ± 0.74 a | 63.5 ± 0.76 d | 38.3 ± 0.53 f | 100.5 ± 0.50 c | 113.5 ± 0.52 b | |
| p-Hydroxybenzoic acid | 222.3 ± 0.86 a | 38.5 ± 0.38 c | 154.3 ± 1.0 b | 35.8 ± 0.43 c | 36.8 ± 0.45 c | 27.0 ± 0.43 d | |
| Syringic acid | 7.0 ± 0.15 d | 12.3 ± 0.11 c | 6.8 ± 0.10 d | 13.0 ± 0.10 b | 27.3 ± 0.11 a | n.d. e | |
| p-coumaric acid | 25.3 ± 0.53 c | 26.5 ± 0.43 c | 84.5 ± 0.59 a | 16.3 ± 0.24 d | 74.8 ± 0.61 b | 29.5 ± 0.30 c | |
| Hydroxycinnamic acids | Chlorogenic acid | n.d. d | 87.8 ± 0.47 c | n.d. d | 37.5 ± 0.33 c | 1057.0 ± 1.58 a | 587.8 ± 1.20 b |
| Vanillic acid | 31.8 ± 0.56 c | n.d. d | 37.0 ± 0.22 b | n.d. d | 79.3 ± 0.65 a | n.d. d | |
| Caffeic acid | 14.5 ± 0.11 f | 46.3 ± 0.38 c | 26.8 ± 0.30 e | 35.0 ± 0.52 d | 102.5 ± 0.65 b | 134.8 ± 0.60 a | |
| Sinapinic acid | 4.0 ± 0.09 b | n.d. c | n.d. c | n.d. c | n.d. c | 11.8 ± 0.07 a | |
| Ferulic acid | 39.8 ± 0.34 c | 11.8 ± 0.19 d | 552.0 ± 1.11 b | 2.5 ± 0.05 e | 35.5 ± 0.33 c | 1143.0 ± 2.86 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šćepanović, M.; Sarić-Krsmanović, M.; Šoštarčić, V.; Brijačak, E.; Lakić, J.; Špirović Trifunović, B.; Gajić Umiljendić, J.; Radivojević, L. Inhibitory Effects of Brassicaceae Cover Crop on Ambrosia artemisiifolia Germination and Early Growth. Plants 2021, 10, 794. https://doi.org/10.3390/plants10040794
Šćepanović M, Sarić-Krsmanović M, Šoštarčić V, Brijačak E, Lakić J, Špirović Trifunović B, Gajić Umiljendić J, Radivojević L. Inhibitory Effects of Brassicaceae Cover Crop on Ambrosia artemisiifolia Germination and Early Growth. Plants. 2021; 10(4):794. https://doi.org/10.3390/plants10040794
Chicago/Turabian StyleŠćepanović, Maja, Marija Sarić-Krsmanović, Valentina Šoštarčić, Ema Brijačak, Josip Lakić, Bojana Špirović Trifunović, Jelena Gajić Umiljendić, and Ljiljana Radivojević. 2021. "Inhibitory Effects of Brassicaceae Cover Crop on Ambrosia artemisiifolia Germination and Early Growth" Plants 10, no. 4: 794. https://doi.org/10.3390/plants10040794