
The Diversity of Volatile Compounds in Australia’s Semi-Desert Genus Eremophila (Scrophulariaceae)

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
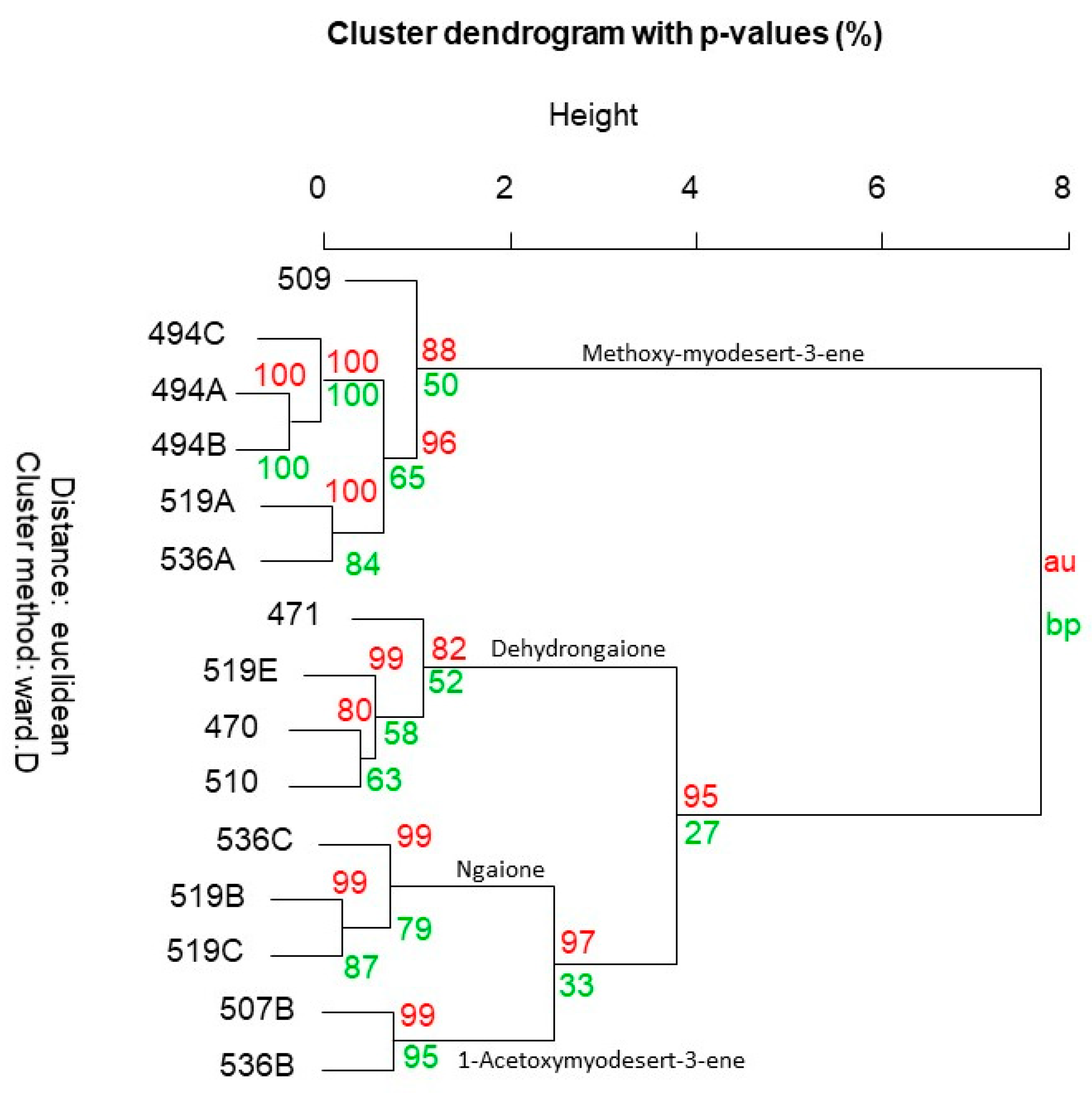
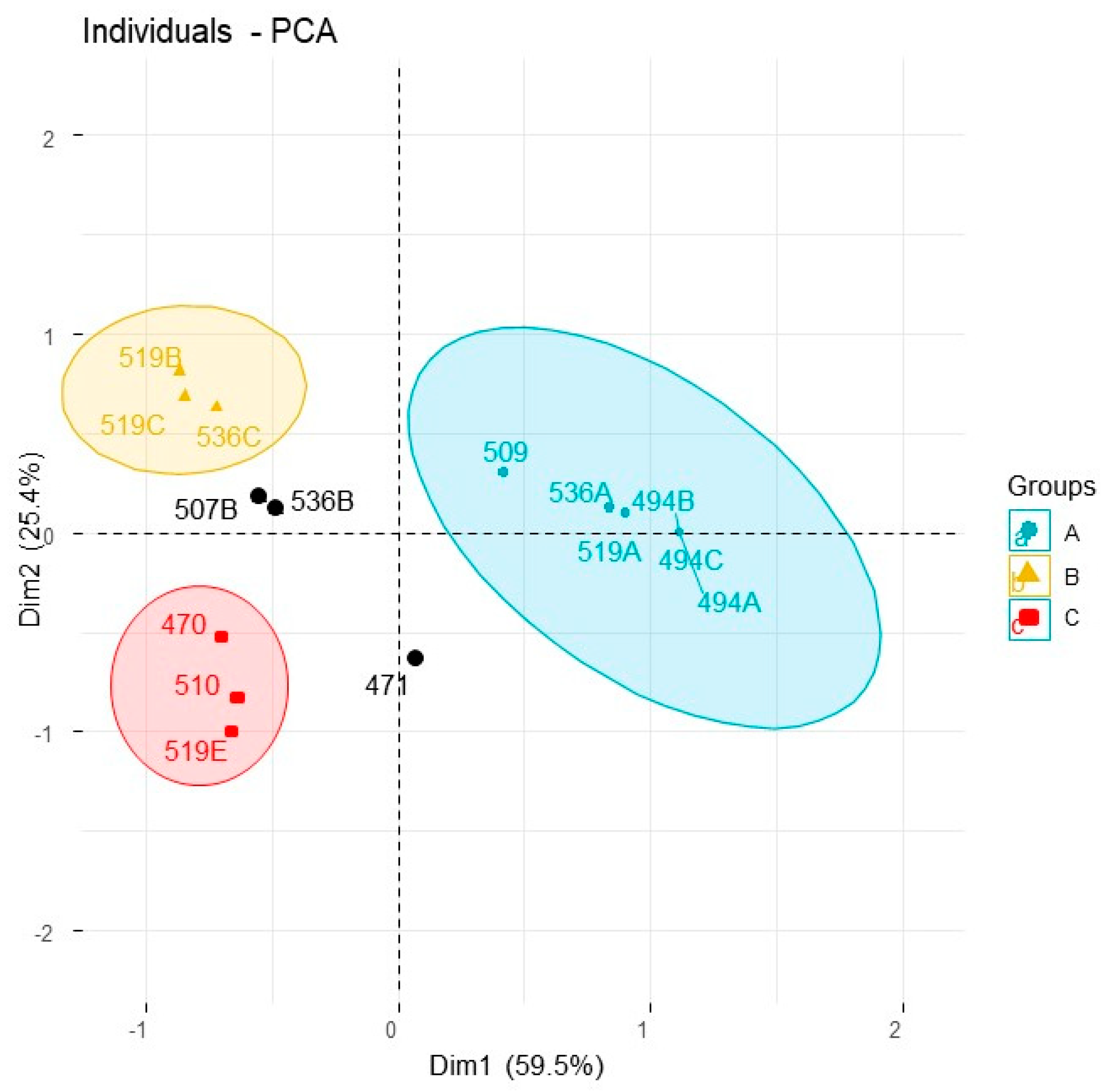
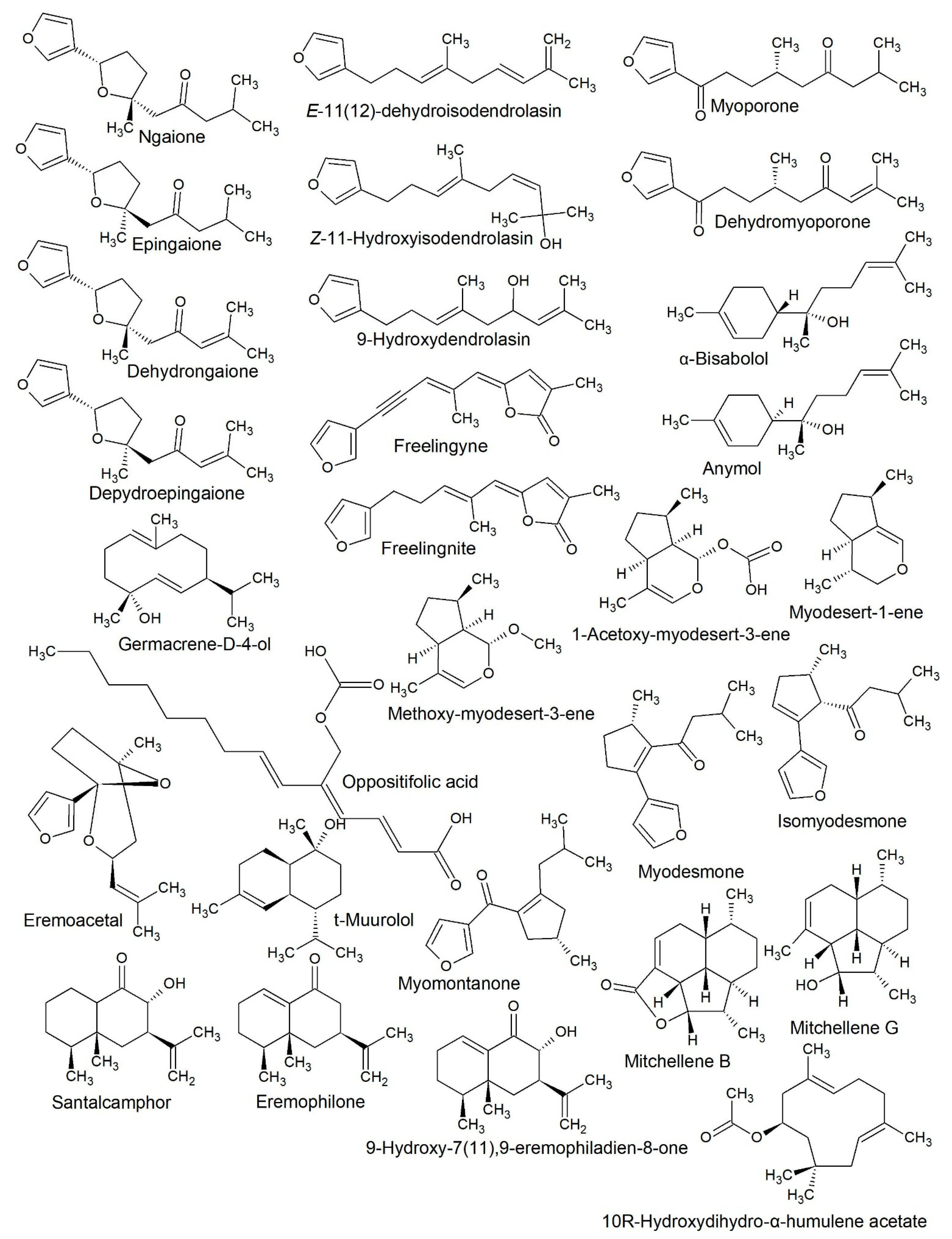
2.1. Eremophila Deserti

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Code (See Table 1 and Table 2) | 470 | 471 | 494A | 494B | 494C | 507B | 509-May | 510-Oct | 519A | 519B | 519C | 519E | 536A | 536B | 536C | 105 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield w/w % Fresh Leaves | 0.3 | 0.5 | 2.3 | 1.1 | 4.1 | 0.4 | 0.3 | 0.5 | 1.5 | 1.7 | 0.8 | 1.3 | 1.4 | 0.2 | 0.9 | 1.1 | ||
| Compound | AI | Pub AI | ||||||||||||||||
| Myodesert-1-ene | 1139 | NMR | - | - | - | - | - | 14.3 | - | - | - | - | - | - | - | 0.7 | - | - |
| β-Pinene oxide | 1161 | 1154 | - | - | - | - | - | 0.5 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 1281 | - | - | - | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - | - |
| Methoxymyodesert-3-ene | 1282 | NMR | - | 39.3 | 100.0 | 100.0 | 100.0 | - | 71.8 | - | 97.1 | - | - | - | 95.3 | - | - | 100 |
| Z,E-Iridodial-1 | 1291 | - | - | - | - | - | - | 3.3 | - | - | - | - | - | - | - | 3.7 | - | - |
| Z,E-Iridodial-2 | 1296 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 4.9 | - | - |
| 2-Ethylfenchol | 1297 | 1297 | - | - | - | - | - | 4.8 | - | - | - | - | - | - | - | - | - | - |
| E,E-Iridodial | 1314 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3.2 | - | - |
| 2E,4E-Decadienal | 1316 | 1315 | - | - | - | - | - | 3.2 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 1321 | - | - | - | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 1324 | - | - | - | - | - | - | 0.3 | - | - | - | - | - | - | - | - | - | - |
| cis,cis-Nepetalactol | 1335 | - | - | - | - | - | - | 3.4 | - | - | - | - | - | - | - | 10.2 | - | - |
| α-Cubebene | 1350 | 1345 | - | - | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - | - |
| β-Damascenone | 1381 | 1383 | - | - | - | - | - | 0.2 | - | - | - | - | - | - | - | - | - | - |
| α-Duprezianene | 1390 | 1387 | - | - | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - | - |
| β-Elemene | 1395 | 1389 | 0.7 | 0.8 | - | - | - | 1.0 | - | 1.7 | - | - | - | - | - | 0.7 | - | - |
| cis,cis-Nepetalactone | 1395 | 1391 | - | - | - | - | - | 1.2 | - | - | - | - | - | - | - | 0.6 | - | - |
| E-Caryophyllene | 1424 | 1417 | 1.2 | 0.5 | - | - | - | 1.2 | - | 2.3 | - | - | - | - | - | 1.5 | - | - |
| β-Farnesene | 1441 | 1440 | - | - | - | - | - | 0.2 | - | - | - | - | - | - | - | - | - | - |
| β-Santalene | 1455 | 1457 | - | - | - | - | - | 0.3 | - | - | - | - | - | - | - | - | - | - |
| (1S)-1-Acetoxymyodesert-3-ene * | 1460 | NMR | - | - | - | - | - | 23.8 | - | - | - | - | - | - | - | 48.4 | - | - |
| (1S)-1-Acetoxymyodesert-3-ene epimer * | 1471 | NMR | - | - | - | - | - | 0.8 | - | - | - | - | - | - | - | 3.5 | - | - |
| Germacrene D | 1485 | 1484 | 1.7 | 0.6 | - | - | - | 2.5 | - | 4.8 | - | - | - | - | - | 2.6 | 1.1 | - |
| Bicydogermlacrene | 1500 | 1500 | 3.7 | 2.5 | - | - | - | 11.1 | - | 3.1 | - | 0.3 | - | - | - | 6.1 | 2.1 | - |
| δ-Cadinene | 1524 | 1522 | - | - | - | - | - | 0.7 | - | 1.4 | - | - | - | - | - | - | - | - |
| Spathulenol | 1579 | 1577 | - | 1.0 | - | - | - | 1.9 | - | - | - | - | - | - | - | 0.9 | - | - |
| Presillhiperfolan-8-β-ol | 1585 | 1585 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.5 | - | - |
| Gleenol | 1585 | 1586 | - | - | - | - | - | 0.5 | - | - | - | - | - | - | - | - | - | - |
| AIIo-cedrol | 1589 | 1589 | - | 0.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Viridiflorol | 1593 | 1592 | - | - | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - | - |
| Carotol | 1598 | 1594 | - | - | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - | - |
| Hinesol | 1639 | 1640 | - | - | - | - | - | 0.3 | - | - | - | - | - | - | - | - | - | - |
| Myomontanone | 1647 | NMR | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.6 | - | - |
| Neo-intermedeol | 1658 | 1658 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.9 | - | - |
| Gymnomilrol | 1658 | 1658 | - | - | - | - | - | 0.2 | - | - | - | - | - | - | - | - | - | - |
| Bulnesol | 1668 | 1670 | - | - | - | - | - | 0.5 | - | - | - | - | - | - | - | - | - | - |
| Z-11-Hydroxyisodendrolasin | 1678 | NMR | - | - | - | - | - | 0.2 | - | - | - | - | - | - | - | 0.6 | - | - |
| Ngaione | 1693 | NMR | 23.6 | 0.6 | - | - | - | 19.8 | 25.8 | 3.7 | 2.9 | 99.4 | 96.6 | 4.6 | 4.7 | 8.6 | 95.7 | - |
| Epingaione | 1695 | NMR | - | - | - | - | - | 0.3 | - | - | - | - | 1.4 | - | - | - | - | - |
| Dehydroepingaione | 1702 | NMR | 1.9 | 0.8 | - | - | - | - | - | 3.2 | - | - | - | 1.6 | - | 0.6 | - | - |
| Dehydrongaione | 1759 | NMR | 66.6 | 49.5 | - | - | - | 0.3 | - | 78.5 | - | 0.3 | 2.0 | 93.8 | - | - | - | - |
| n.d. | 1762 | - | 0.7 | 3.6 | - | - | - | - | - | 1.3 | - | - | - | - | - | - | - | - |
| n.d. | 1838 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.5 | - | - |
2.2. Traditional Antibacterial Species
2.2.1. Eremophila duttonii and E. neglecta
2.2.2. Eremophila freelingii
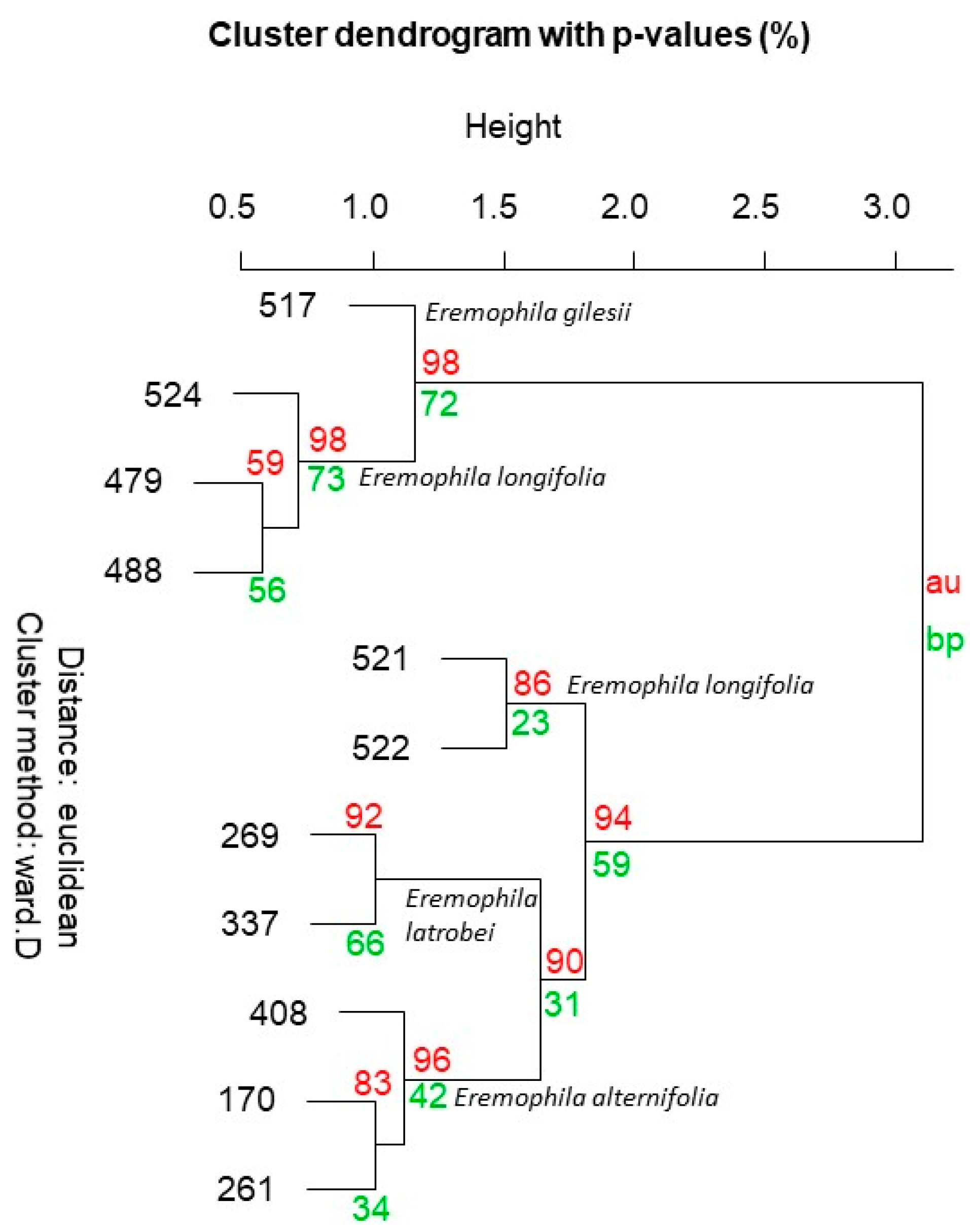
2.2.3. Eremophila alternifolia and E. latrobei
2.2.4. Eremophila longifolia
2.3. Taxonomic Misdeterminations
2.3.1. Eremophila mitchellii and E. sturtii
2.3.2. Eremophila arbuscular and E. oppositifolia
| Species Code (See Table 1 and Table 2) | Ene | Edut | Epla-53 | Efo-57 | Epla-61 | Epla-62 | Egi-341 | Eyo-345 | Efre-346 | Earb-486 | Earb-487 | EopR-535B | EopO-538A | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield w/w % Fresh Leaves | 0.3 | 0.4 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 2.8 | 0.3 | 0.3 | 0.1 | 0.2 | 0.2 | ||
| Compound | AI | Pub AI | |||||||||||||
| 1,3,5,7-Cyclooctatetraene | 898 | 900 | - | - | - | - | - | - | - | - | - | - | 8.8 | - | - |
| α-Thujene | 928 | 924 | 1.7 | - | - | 0.3 | - | 2.2 | - | - | - | - | - | - | - |
| α-Pinene | 933 | 932 | 17.1 | 74.6 | 21.3 | 7.1 | 15.6 | 27.0 | 13.7 | 1.7 | 27.3 | 2.7 | 10.1 | - | - |
| Camphene | 946 | 946 | - | 1.2 | 13.1 | - | 0.7 | 26.2 | - | - | - | - | - | - | - |
| 3,5-Dimethyl-4-heptanone | 966 | 973 | - | - | - | - | - | - | - | - | - | - | 8.1 | - | - |
| Sabinene | 970 | 969 | 34.4 | - | - | 0.5 | - | - | - | - | - | - | - | - | - |
| β-Pinene | 975 | 974 | 1.3 | 4.0 | 1.8 | 12.9 | 5.4 | 2.0 | - | 6.6 | 3.8 | - | 0.6 | - | - |
| Butanoic acid, butyl ester | 990 | 990 | - | - | - | - | - | - | - | - | - | - | - | - | 2.2 |
| Myrcene | 992 | 988 | - | - | - | 2.4 | - | - | 12.2 | 1.6 | - | - | 1.2 | - | - |
| α-Phellandrene | 1005 | 1002 | 1.4 | - | - | - | - | - | 3.2 | - | - | - | - | - | - |
| α-Terpinene | 1019 | 1014 | 4.4 | - | - | - | - | - | - | 1.2 | - | - | - | - | - |
| p-Cymene | 1026 | 1022 | - | - | - | - | - | - | - | 3.5 | - | - | - | - | - |
| Limonene | 1028 | 1024 | - | 0.9 | 7.0 | 1.7 | 0.6 | 7.7 | - | 1.7 | - | 0.9 | 2.4 | - | - |
| β-Phellandrene | 1032 | 1029 | 8.0 | - | - | - | 2.4 | 8.3 | 46.3 | - | - | - | - | - | - |
| 1,8-Cineole | 1034 | 1026 | - | - | - | - | - | - | - | 73.1 | - | - | - | - | - |
| β-Ocimene | 1048 | 1044 | - | - | - | 0.3 | - | - | 0.6 | - | - | 1.0 | - | - | - |
| γ-Terpinene | 1060 | 1054 | 7.0 | - | - | - | - | - | - | 2.1 | - | - | - | - | - |
| α-Terpinolene | 1090 | 1086 | 1.7 | - | - | - | - | - | - | - | - | - | - | - | - |
| Linalool | 1098 | 1095 | - | - | - | - | - | - | - | - | - | 1.1 | - | - | - |
| 3-Methyl-3-butenyl-methyl butanoate | 1111 | 1112 | - | - | - | - | - | - | - | - | - | 1.1 | - | - | - |
| Terpinen-4-ol | 1180 | 1174 | 4.7 | - | - | - | - | - | - | 6.1 | - | - | - | - | - |
| Butanoic acid, 1-methylhexyl ester | 1216 | 1210 | - | - | - | - | - | - | - | - | - | - | - | - | 7.9 |
| n.d. | 1240 | - | - | - | - | - | - | - | - | - | - | - | 0.6 | - | - |
| n.d. | 1279 | - | - | - | - | - | - | - | - | - | - | - | 2.0 | - | - |
| Bornyl acetate | 1294 | 1284 | - | - | 3.4 | - | 0.9 | - | - | - | - | - | - | - | - |
| Geranyl acetate | 1366 | 1362 | - | - | - | - | - | 3.2 | - | - | - | - | - | - | - |
| β-Patchoulene | 1383 | 1379 | - | - | - | - | - | 3.3 | - | - | - | - | - | - | - |
| β-Elemene | 1396 | 1389 | - | - | - | 1.5 | 1.1 | - | 1.3 | - | - | - | - | - | - |
| β-Duprezianene | 1418 | 1421 | - | - | - | - | - | - | - | - | - | - | 0.6 | - | - |
| E-Caryophyllene | 1425 | 1417 | - | - | - | - | 3.5 | - | 0.5 | - | - | - | - | 0.8 | 20.7 |
| 9-Epi-E-caryophyllene | 1462 | 1464 | - | - | - | 0.4 | - | - | - | - | - | - | - | - | - |
| 1-Amorpha-4,7(11)-diene | 1480 | 1479 | - | - | - | 0.3 | - | - | - | - | - | - | - | - | - |
| Z-β-Guaiene | 1485 | 1492 | - | - | - | 5.2 | - | - | 1.3 | - | - | - | - | - | - |
| β-Selinene | 1490 | 1489 | - | - | - | 0.4 | - | - | 0.5 | - | - | - | - | - | - |
| Bicyclogermacrene | 1497 | 1500 | 3.6 | 0.7 | 4.0 | 2.3 | 7.9 | 9.9 | 2.3 | - | 7.2 | - | 4.3 | 3.4 | - |
| Eremophilene | 1503 | 1502 | - | - | - | 1.5 | - | - | 0.6 | - | - | - | - | - | - |
| δ-Cadinene | 1522 | 1522 | 1.8 | - | - | 0.8 | - | - | 0.5 | - | - | - | - | 1.4 | - |
| α-Cadinene | 1529 | 1537 | - | - | - | 0.4 | 3.9 | - | 1.3 | - | - | - | - | 0.5 | - |
| Elemol | 1554 | 1548 | - | - | - | 11.0 | - | - | - | - | 8.2 | - | - | - | - |
| Spathulenol | 1579 | 1577 | - | 0.7 | - | - | 1.8 | - | 0.5 | 1.0 | 21.3 | - | 1.3 | 0.6 | - |
| Caryophyllene oxide | 1584 | 1582 | 2.6 | - | - | - | - | - | - | - | - | - | - | 0.9 | 29.4 |
| Globulol | 1585 | 1590 | - | - | - | 2.0 | 4.4 | 4.1 | - | - | - | - | - | - | - |
| Cubeban-11-ol | 1595 | 1595 | - | - | - | - | - | - | 0.7 | - | - | - | 0.6 | 1.5 | - |
| Ledol | 1596 | 1610 | - | - | - | 6.8 | 1.8 | - | - | - | - | - | 0.8 | - | - |
| E-11(12)-Dehydroisodendrolasin | 1597 | NMR | - | - | - | - | - | - | - | - | - | 10.8 | - | 1.2 | - |
| Humulene epoxide II | 1611 | 1608 | - | - | - | - | - | - | - | - | - | - | 0.5 | 1.4 | - |
| Guaiol | 1612 | 1600 | - | - | - | - | 4.6 | - | - | - | - | - | - | 3.1 | - |
| 10-Epi-7-eudesmol | 1621 | 1622 | - | - | 3.1 | 1.0 | - | - | - | - | - | - | 1.5 | 1.0 | - |
| γ-Eudesmol | 1638 | 1630 | - | - | 2.3 | 0.4 | 0.6 | - | 0.8 | - | - | - | - | 0.5 | - |
| t-Muurolol | 1645 | 1644 | - | - | - | - | - | - | - | - | - | - | - | 42.9 | - |
| Myomontanone | 1647 | NMR | - | - | - | - | - | - | 1.3 | - | - | - | 49.2 | - | - |
| Eudesmol isomer | 1649 | - | - | - | - | 3.9 | 2.2 | - | - | - | - | - | - | - | - |
| β-Eudesmol | 1653 | 1649 | - | - | 23.1 | 1.4 | 4.1 | 3.3 | 3.5 | - | - | - | - | - | - |
| α-Eudesmol | 1657 | 1652 | - | - | 2.1 | 24.9 | 6.8 | - | 4.3 | - | - | - | 4.7 | 4.1 | - |
| n.d. | 1677 | - | - | - | - | - | - | - | - | - | - | - | 1.4 | - | |
| Z-11-Hydroxyisodendrolasin | 1678 | NMR | - | - | - | - | - | - | - | - | - | 50.4 | - | 4.8 | - |
| Anymol | 1686 | NMR | - | - | 3.0 | - | - | - | - | - | - | - | - | - | - |
| Epignaione | 1694 | NMR | - | - | - | - | - | - | - | 1.3 | - | - | - | - | - |
| n.d. | 1700 | - | - | - | 3.7 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 1715 | - | - | - | 4.2 | - | - | - | - | - | - | - | - | - | - |
| Redbank’s Ketol | 1721 | * | - | - | - | - | - | - | - | - | 10.2 | - | 0.7 | 0.6 | - |
| Farnesol | 1724 | 1722 | 10.3 | - | 4.6 | - | 26.0 | 2.8 | 3.3 | - | - | 3.8 | 0.6 | 0.6 | - |
| 9-Hydroxydendrolasin | 1741 | NMR | - | - | - | - | - | - | - | - | - | 28.1 | - | 0.6 | - |
| Dehydrongaione | 1752 | NMR | - | 12.3 | - | - | 2.1 | - | - | - | - | - | - | 3.8 | - |
| 10R-Hydroxydihydro-α-humulene acetate | 1755 | NMR | - | - | - | - | - | - | - | - | - | - | - | - | 39.7 |
| n.d. | 1760 | - | - | - | - | 7.3 | - | - | - | - | - | - | - | - | - |
| Myoporone | 1842 | NMR | - | - | - | - | - | - | - | - | 22.0 | - | - | 0.6 | - |
| Phytol | 1914 | 1915 | - | 5.7 | - | - | - | - | - | - | - | - | - | - | - |
| Oppositifolic acid | 2149 | NMR ** | - | - | - | - | - | - | - | - | - | - | - | 24.1 | - |
| Eremophila freelingii—DCM Extract | ||||||
|---|---|---|---|---|---|---|
| Species Code (See Table 1 and Table 2) | 338-A | 338-B | 338-C | 338-D | ||
| Compound ID | AI | Pub. AI | ||||
| α-Pinene | 934 | 932 | 9.2 | 12.7 | 17.6 | 4.3 |
| Germacrene D | 1485 | 1484 | - | - | - | 2.8 |
| Bicyclogermacrene | 1500 | 1500 | 4.5 | - | 9.2 | 10.8 |
| n.d. | - | - | 1.7 | - | - | - |
| Elemol | 1554 | 1548 | 5.0 | - | 6.7 | - |
| Myoporone | 1836 | NMR | 23.2 | 17.3 | 37.2 | 54.2 |
| Dehydromyoporone | 1901 | NMR | - | - | 4.2 | 7.1 |
| n.d. | - | - | 6.0 | 4.1 | 5.2 | 6.7 |
| 2-Methyltetradecane | 1461 | 1462 ** | 32.7 | - | - | - |
| n.d. | - | - | 4.6 | 3.9 | 5.9 | 5.1 |
| n.d. | - | - | 3.8 | 2.9 | 4.8 | 3.8 |
| n.d. | - | - | 5.4 | 3.9 | 5.2 | 5.2 |
| Octacosane | 1800 | 1800 | - | 5.9 | - | - |
| n.d.-alkane | - | - | 3.9 | - | 1.9 | - |
| Nonacosane | 1900 | 1900 | - | 49.3 | 2.1 | - |
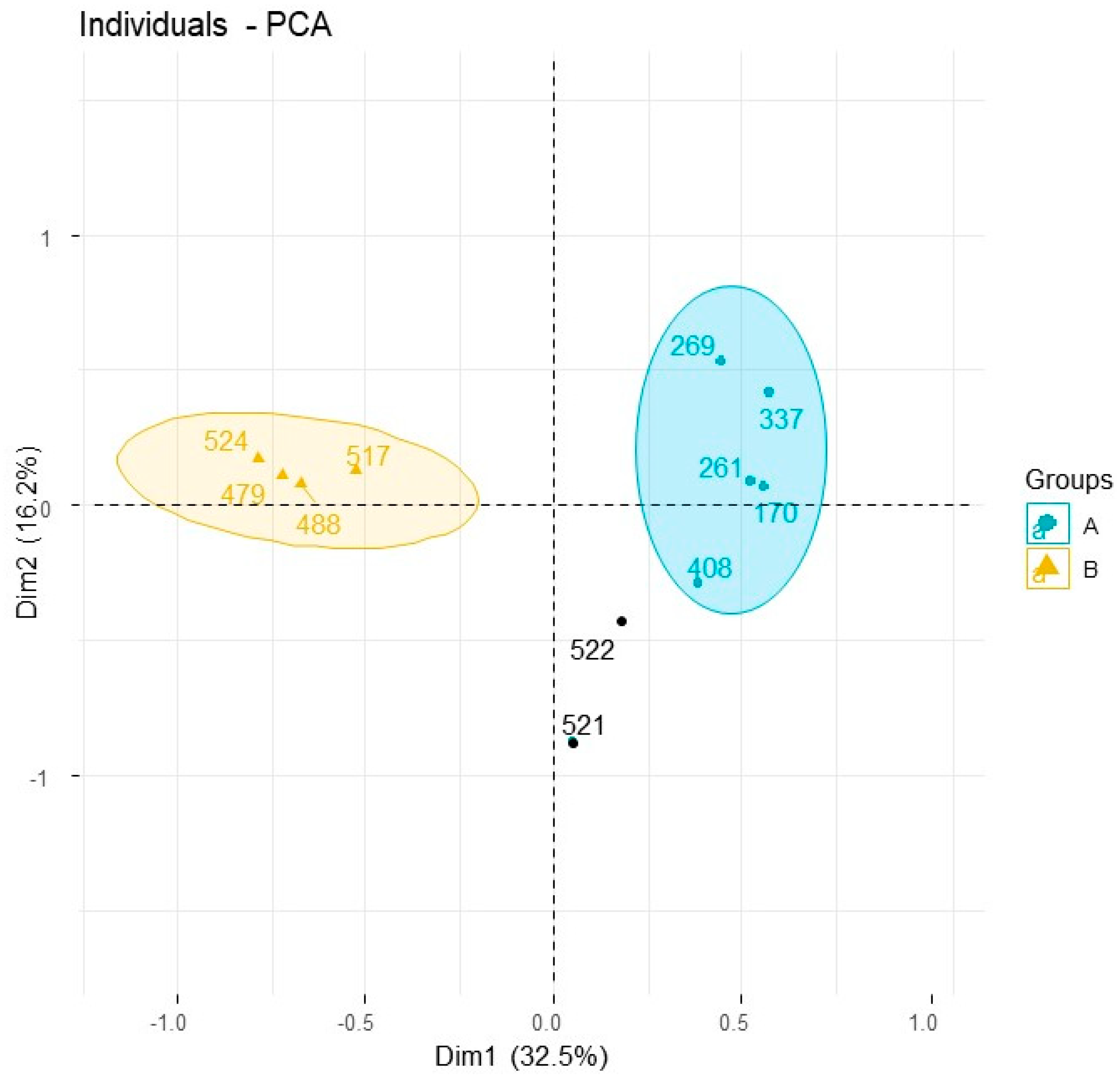
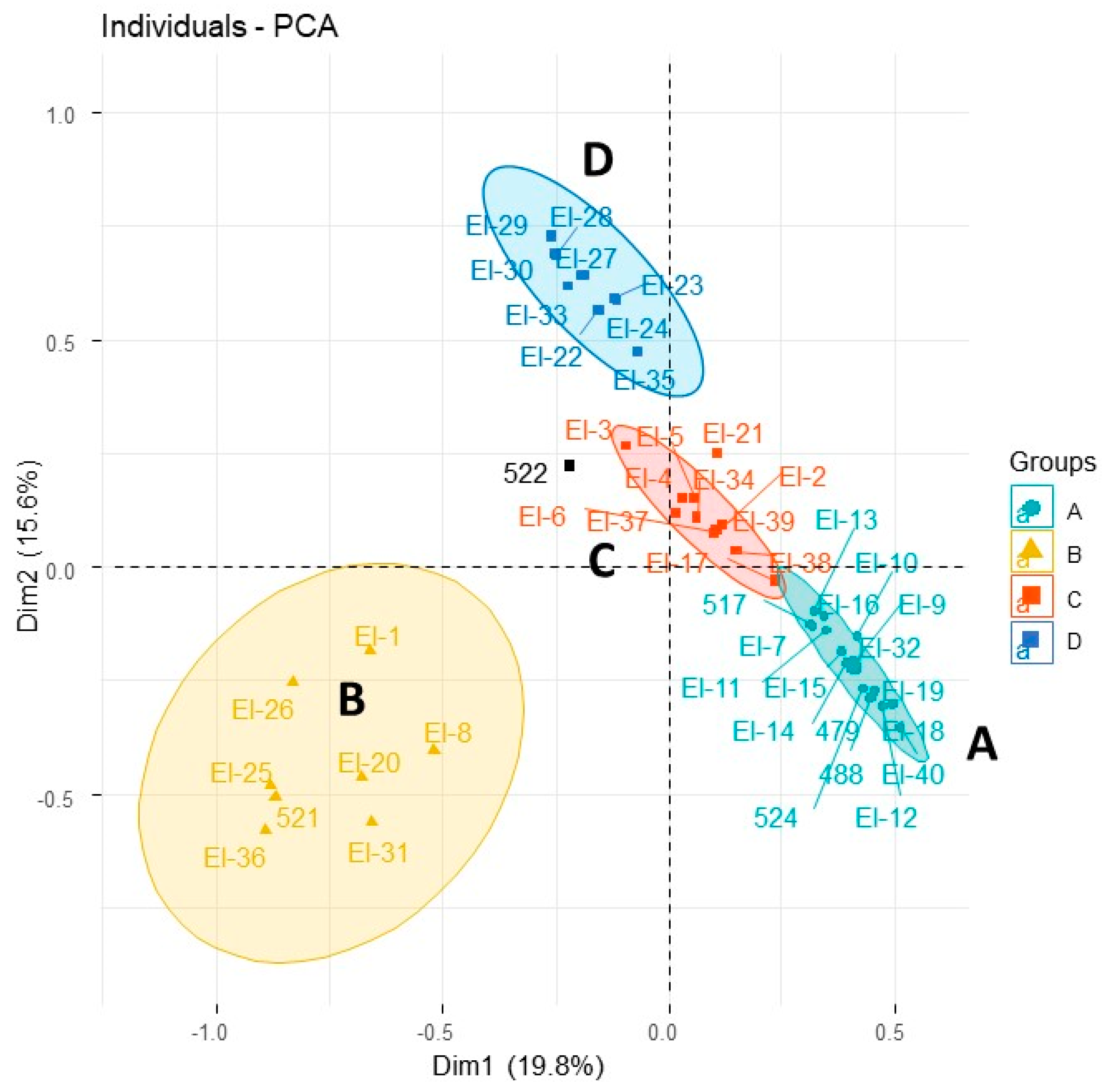
| Species Code (See Table 1 and Table 2) | Elo-517 | Elo-521 | Elo-522 | Elo-524 | Elo-479 | Elo-488 | Ealt-170 | Ealt-261 | Ealt-408 | Elat-269 | Elat-337 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield w/w % Fresh Leaves | 0.7 | 0.8 | 3.2 | 0.9 | 0.4 | 0.1 | 0.6 | 0.7 | 1.2 | 0.4 | 0.5 | ||
| Compund Common Name | AI | Pub AI | - | - | - | - | - | - | - | - | - | - | - |
| α-Thujene | 924 | 924 | - | - | - | - | - | - | - | 0.5 | 2.5 | - | - |
| α-Pinene | 932 | 932 | - | - | - | 0.9 | 5.6 | 2.0 | - | 1.5 | 6.6 | - | - |
| α-Fenchene | 945 | 945 | 2.0 | - | - | - | - | - | 2.3 | 7.0 | 23.2 | 1.3 | - |
| Sabinene | 970 | 969 | - | - | - | - | 6.5 | 11.4 | - | - | - | - | - |
| β-Pinene | 975 | 974 | - | - | - | - | - | - | - | 1.7 | 2.9 | - | - |
| δ-Carene | 998 | 1001 | - | - | 1.1 | - | 1.6 | 3.7 | - | - | - | - | - |
| α-Phellandrene | 1007 | 1002 | - | - | 2.8 | - | 0.8 | 0.8 | - | 0.8 | 2.9 | - | - |
| α-Terpinene | 1016 | 1014 | - | - | 1.1 | 0.7 | 1.4 | 2.0 | - | - | 1.5 | - | - |
| p-Cymene | 1024 | 1022 | - | - | - | - | - | - | 1.0 | 0.4 | 0.8 | - | - |
| β-Phellandrene | 1026 | 1025 | - | - | 4.9 | - | - | - | - | 3.4 | - | - | - |
| Limonene | 1028 | 1024 | 38.5 | 3.7 | - | 69.4 | 62.4 | 58.5 | 0.8 | - | 12.7 | - | - |
| 1,8-Cineole | 1031 | 1026 | - | - | - | - | - | - | 1.1 | 7.8 | 19.5 | - | - |
| γ-Terpinene | 1059 | 1054 | - | - | - | - | 1.3 | 2.3 | - | 0.4 | 1.7 | - | - |
| α-Terpinolene | 1087 | 1086 | - | - | - | 1.8 | 8.7 | 15.3 | - | - | - | - | - |
| Fenchone | 1087 | 1083 | 1.3 | - | - | - | - | - | - | - | 4.9 | - | - |
| E-Mentha-2,8-dien-1-ol | 1119 | 1119 | - | - | 3.7 | - | - | - | - | - | - | - | - |
| Geijerene | 1141 | 1138 | - | - | 1.9 | - | - | - | - | - | - | - | 0.3 |
| Karahanaenone | 1154 | 1154 | - | 80.9 | 0.5 | - | - | - | - | - | 3.1 | - | - |
| Isomenthone | 1163 | 1158 | - | - | 26.4 | - | - | - | - | - | - | - | - |
| Borneol | 1165 | 1165 | 2.7 | - | - | - | - | - | - | - | - | - | - |
| Terpinen-4-ol | 1179 | 1174 | 0.4 | 4.7 | - | - | 1.3 | 3.4 | 0.9 | 0.8 | 2.2 | - | - |
| α-Terpineol | 1190 | 1186 | 1.5 | 1.1 | 11.3 | - | - | - | - | - | 1.1 | - | - |
| Z-Piperitol | 1196 | 1195 | - | - | 42.1 | - | - | - | - | - | - | - | - |
| E-Piperitol | 1206 | 1207 | - | - | 2.2 | - | - | - | - | - | - | - | - |
| Fenchyl acetate | 1219 | 1218 | 10.0 | - | - | 1.6 | - | - | - | - | - | - | - |
| Nerol | 1227 | 1227 | - | - | - | - | - | - | 2.9 | - | - | - | - |
| Asearidole | 1237 | 1234 | - | - | 0.4 | - | - | - | - | - | - | - | - |
| Piperitone | 1252 | 1249 | - | - | 0.5 | - | - | - | - | - | - | - | - |
| 2-(1E)-Propenyl-phenol | 1266 | 1264 | - | - | - | - | 3.5 | - | - | - | - | - | - |
| Bornyl acetate | 1285 | 1287 | 43.0 | - | - | 5.3 | - | - | - | - | - | 1.0 | - |
| Safrole | 1288 | 1287 | - | - | - | 20.4 | 5.6 | - | - | - | - | - | - |
| 3-Thujanol acetate | 1297 | 1295 | - | - | - | - | - | - | - | - | - | 1.8 | - |
| p-Vinyguaiacol | 1311 | 1309 | - | 4.8 | 0.7 | - | - | - | - | - | - | - | - |
| Dictamnol | 1429 | 1428 | - | - | - | - | - | - | - | - | - | - | 0.7 |
| Alloaromadendrene | 1440 | 1439 | - | - | - | - | - | - | 0.4 | - | - | - | - |
| β-Macrocarpene | 1496 | 1499 | - | - | - | - | - | - | - | - | 0.5 | - | - |
| β-Germacrene | 1501 | 1508 | - | - | - | - | - | 0.7 | - | - | - | - | - |
| Bicyclogermacrene | 1501 | 1500 | - | - | - | - | 1.4 | - | - | 0.5 | - | - | - |
| δ-Cadinene | 1523 | 1522 | - | - | - | - | - | - | 0.4 | 0.5 | 0.5 | - | 0.2 |
| Kessane | 1535 | 1529 | - | - | - | - | - | - | - | - | - | 1.0 | - |
| n.d. | 1549 | - | - | - | - | - | - | - | - | - | - | 3.6 | 3.1 |
| Maaliol | 1568 | 1566 | - | - | - | - | - | - | 0.5 | - | - | - | 0.3 |
| Spathulenol | 1579 | 1577 | - | - | - | - | - | - | 8.4 | 0.6 | 0.4 | - | 1.7 |
| Cedrol | 1591 | 1600 | - | - | - | - | - | - | 1.4 | - | 0.5 | 3.0 | - |
| Rosifoliol | 1603 | 1600 | - | - | - | - | - | - | 0.5 | - | - | 3.4 | - |
| n.d. | 1614 | - | - | - | - | - | - | - | - | - | - | 2.4 | 0.2 |
| 10-Epi-γ-eudesmol | 1621 | 1622 | - | - | - | - | - | - | - | - | - | 2.0 | 1.4 |
| Eudesmol | 1624 | 1622 | - | 0.9 | - | - | - | - | 1.6 | - | 0.7 | - | - |
| γ-Eudesmol | 1632 | 1630 | - | - | - | - | - | - | - | - | - | 1.2 | 0.3 |
| Hinesol | 1642 | 1640 | - | - | - | - | - | - | 0.6 | 0.4 | 0.6 | 2.5 | 1.4 |
| Myomontanone | 1647 | NMR | - | - | - | - | - | - | 5.9 | 3.4 | 1.0 | 1.9 | 2.3 |
| Isomyodesmone | 1650 | * | - | - | - | - | - | - | 0.7 | - | - | 4.5 | 3.7 |
| Carr’s Ketol | 1658 | * | - | - | - | - | - | - | 4.5 | 39.0 | 0.8 | 1.9 | 7.3 |
| Bulnesol | 1668 | 1670 | - | - | - | - | - | - | - | - | - | 1.5 | 0.4 |
| n.d. | 1677 | - | - | - | - | - | - | - | 2.8 | - | 0.8 | 2.7 | 2.6 |
| Kindon’s Ketol | 1681 | * | - | - | - | - | - | - | - | 1.1 | - | - | 2.2 |
| Anymol | 1686 | NMR | - | - | - | - | - | - | - | - | - | 63.5 | 19.4 |
| Epignaione | 1697 | NMR | - | - | - | - | - | - | - | - | - | - | 26.8 |
| n.d. | 1726 | - | - | - | - | - | - | - | 0.4 | - | - | - | 5.9 |
| Carney’s Ketol | 1734 | * | - | - | - | - | - | - | - | 4.7 | - | - | 0.2 |
| Dehydrongaione | 1752 | NMR | - | - | - | - | - | - | - | - | - | - | 0.4 |
| Myoporone | 1836 | NMR | - | 3.3 | - | - | - | - | 19.4 | 22.7 | 5.0 | - | 17.4 |
| Dehydromyoporone | 1901 | NMR | - | - | - | - | - | - | 43.2 | 1.8 | 3.3 | - | 0.9 |
| Species Code (See Table 1 and Table 2) | EaltMm | Ealt-A | Ealt-B | Ealt-C | EaltL-A | EaltL-B | EaltL-C | ElatF | ElatG | ElatL | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Common Name | AI | Pub. AI | - | - | - | - | - | - | - | - | - | - |
| α-Thujene | 929 | 924 | 2.8 | 3.7 | 2.6 | 1.6 | - | - | - | - | - | - |
| α-Pinene | 934 | 932 | 6.7 | 10.0 | 7.3 | 5.5 | 2.6 | 1.9 | 1.8 | - | 0.7 | 0.4 |
| α-Fenchene | 945 | 945 | - | - | 12.0 | 9.7 | 14.1 | 6.9 | 6.4 | 6.3 | 0.5 | 7.4 |
| Sabinene | 970 | 969 | 4.6 | 8.9 | 5.3 | 4.4 | 0.3 | 0.5 | - | - | - | - |
| β-Pinene | 975 | 974 | 1.6 | - | - | - | - | - | - | - | - | - |
| α-Phellandrene | 1007 | 1002 | 5.5 | - | 1.4 | - | 1.5 | 1.0 | - | - | - | - |
| p-Cymene | 1026 | 1022 | 0.5 | 0.7 | 0.3 | 0.7 | - | - | - | - | - | - |
| β-Phellandrene | 1032 | 1029 | 11.5 | 11.9 | 8.9 | 7.5 | 7.1 | 9.3 | 8.5 | - | - | - |
| 1,8-Cineole | 1034 | 1026 | 10.2 | 35.1 | 15.6 | 21.2 | 0.4 | 0.7 | 1.0 | - | - | - |
| γ-Terpinene | 1059 | 1054 | 0.2 | - | 0.3 | 0.2 | 0.2 | - | - | - | - | - |
| Z-Sabinene hydrate | 1065 | 1065 | 0.3 | 1.0 | 0.7 | 1.2 | 0.3 | - | - | - | - | - |
| α-Terpinolene | 1087 | 1086 | 0.2 | - | - | - | 2.8 | 2.0 | - | - | - | - |
| E-Sabinene hydrate | 1097 | 1098 | 0.1 | 0.4 | 0.3 | 0.3 | - | - | - | - | - | - |
| Myodesert-1-ene | 1139 | NMR | - | - | - | - | 21.5 | 46.2 | 64.4 | - | - | - |
| Terpinen-4-ol | 1180 | 1174 | - | - | 0.3 | 0.4 | 0.8 | 1.2 | 1.8 | - | - | - |
| α-Terpineol | 1190 | 1186 | - | 0.4 | 0.2 | 0.3 | - | - | - | - | - | - |
| Endo-fenchyl acetate | 1219 | 1218 | - | - | - | - | - | - | - | 0.7 | - | 1.1 |
| Nerol | 1230 | 1232 | - | - | 0.8 | 1.1 | - | - | - | - | - | - |
| Piperitone | 1253 | 1249 | - | - | 0.5 | 0.7 | - | - | - | - | - | - |
| Methoxymyodesert-3-ene | 1282 | NMR | - | - | - | - | - | - | - | 1.1 | 1.4 | 1.4 |
| p-Vinylguaiacol | 1313 | 1309 | 0.3 | 1.8 | 1.0 | 2.3 | - | 0.8 | - | - | - | - |
| cis,cis-Nepetalactol | 1335 | - | - | - | 0.2 | - | - | - | - | - | - | - |
| cis,cis-Nepetalactone | 1395 | 1391 | - | - | - | 0.4 | - | - | - | - | - | - |
| E-Caryophyllene | 1425 | 1417 | - | - | - | - | - | - | - | 3.9 | 0.5 | - |
| β-Santalene | 1455 | 1457 | - | - | - | - | - | - | - | 2.3 | 0.8 | - |
| (1S)-1-Acetoxymyodesert-3-ene * | 1460 | NMR | 0.2 | - | - | - | 0.3 | 0.6 | - | - | - | - |
| (1S)-1-Acetoxymyodesert-3-ene epimer * | 1471 | - | 0.2 | - | 0.5 | - | 0.4 | 1.1 | - | - | - | - |
| Germacrene D | 1485 | 1484 | - | - | - | - | - | - | - | 1.2 | - | - |
| Bicydogermlacrene | 1500 | 1500 | 1.2 | 0.7 | 2.6 | 0.9 | 2.7 | 1.9 | - | 0.7 | 0.6 | 1.8 |
| δ-Cadinene | 1524 | 1522 | - | - | 0.2 | - | - | - | - | - | - | - |
| Elemol | 1548 | 1548 | - | - | - | - | - | - | - | - | 2.1 | - |
| n.d. | 1563 | - | - | 2.1 | 1.2 | 3.2 | - | 0.6 | - | 0.7 | 2.1 | 0.3 |
| n.d. | 1575 | - | 0.8 | 1.3 | 2.7 | 2.3 | 0.3 | 1.0 | - | - | - | - |
| Caryophyllene oxide | 1583 | 1583 | - | - | - | - | - | - | - | 0.4 | - | - |
| Cedrol | 1591 | 1600 | - | - | - | - | 1.3 | 2.7 | 5.2 | - | 0.7 | - |
| Rosifoliol | 1603 | 1600 | - | 1.6 | 0.5 | 2.0 | - | - | 1.0 | - | - | - |
| Humulene epoxide II | 1611 | 1608 | - | - | - | - | - | - | - | 5.6 | - | 4.8 |
| 10-Epi-7-eudesmol | 1621 | 1622 | - | - | 0.2 | 0.8 | - | - | - | - | - | - |
| Muurola-4,10(14)-dien-1-β-ol | 1628 | 1630 | - | - | - | - | - | - | - | - | 3.2 | 2.2 |
| t-Muurolol | 1645 | 1644 | - | - | - | - | - | - | - | 1.0 | 35.5 | 0.6 |
| Myomontanone | 1647 | 1646 | - | 1.4 | 0.2 | - | 1.4 | - | - | - | - | - |
| Isomyodesmone | 1650 | 1649 | - | - | - | - | - | - | - | - | 3.7 | 2.5 |
| Myodesmone | 1651 | - | - | 1.2 | 1.7 | - | 6.4 | 3.4 | - | - | - | - |
| Bulnesol | 1668 | 1670 | 0.4 | 0.9 | 5.5 | 10.3 | - | - | - | - | 0.4 | - |
| Ketol (Kindon, Perillup or Carr’s ketol) ** | 1681 | ** | - | - | 1.4 | - | 3.1 | 1.2 | - | - | - | - |
| Ngaione | 1682 | NMR | - | - | 0.2 | 0.7 | - | - | - | - | 0.8 | - |
| Dehydroepingaione | 1702 | NMR | - | - | - | - | - | - | - | - | 0.5 | - |
| Ketol (like Carney’s ketol) ** | 1721 | ** | 0.2 | - | 1.3 | - | 0.5 | - | - | - | - | - |
| Diisopropylnaphthalene | 1724 | 1716 | 4.4 | - | 0.9 | - | 3.7 | - | - | - | - | - |
| Ketol (like Carney’s ketol) ** | 1734 | ** | - | - | - | - | - | - | - | - | 0.4 | - |
| n.d. | 1746 | - | - | - | - | - | - | - | - | - | 0.5 | - |
| n.d. | 1788 | - | - | - | - | 0.2 | - | - | 0.9 | - | 1.0 | - |
| n.d. | 1802 | - | - | 0.4 | - | 2.5 | - | - | - | - | - | - |
| n.d. | 1811 | - | - | - | - | - | - | - | - | - | 0.4 | - |
| n.d. | 1815 | - | 0.8 | - | - | - | - | - | - | - | - | 0.6 |
| n.d. | 1819 | - | 0.4 | - | - | - | - | - | - | - | 0.6 | - |
| Myoporone | 1836 | NMR | 0.3 | 1.5 | 4.8 | 1.0 | 15.2 | 6.8 | 0.8 | - | 2.1 | - |
| n.d. | 1846 | - | 0.7 | - | 0.7 | 0.4 | 0.5 | - | - | - | - | - |
| n.d. | 1852 | - | - | - | - | - | - | - | - | - | 5.7 | - |
| n.d. | 1864 | - | 0.3 | 0.5 | 0.2 | 0.3 | - | - | - | - | 1.2 | - |
| n.d. | 1868 | - | - | - | - | - | - | - | - | - | 1.2 | - |
| n.d. | 1872 | - | - | - | 0.4 | 0.4 | - | - | - | - | 0.5 | - |
| n.d. | 1882 | - | - | - | - | - | - | - | - | - | 0.6 | - |
| n.d. | 1890 | - | - | - | - | - | - | - | - | - | 4.4 | - |
| Dehydromyoporone | 1901 | NMR | 12.7 | 0.6 | 8.4 | 0.6 | 8.3 | 2.8 | - | - | 1.4 | - |
| n.d. | 1913 | - | - | - | - | - | - | - | - | - | 8.3 | - |
| n.d. | 1919 | - | 0.6 | 0.8 | 0.4 | 1.2 | - | - | - | - | 0.9 | - |
| n.d. | 1927 | - | - | - | 1.3 | 2.4 | - | - | - | - | - | - |
| n.d. | 1948 | - | 0.2 | - | - | - | - | - | - | - | - | - |
| n-Hexadecanoic acid | 1964 | 1960 | 0.6 | - | 0.9 | 0.3 | 0.6 | 1.3 | 0.7 | - | - | - |
| n.d. | 1958 | - | 0.3 | - | - | - | - | - | - | - | - | - |
| n.d. | 1962 | - | 0.4 | - | - | - | - | - | - | - | - | - |
| n.d. | 1974 | - | - | - | - | - | - | - | - | - | 1.0 | - |
| Freelingnite | 1985 | NMR | 10.6 | - | - | - | - | - | - | - | - | - |
| n.d. | 2017 | - | - | 8.0 | - | 0.4 | - | - | 0.8 | - | 1.6 | - |
| n.d. | 2058 | - | 9.5 | - | - | - | - | - | - | - | - | - |
| n.d. | 2072 | - | - | - | - | - | - | - | - | - | 0.6 | - |
| n.d. | 2078 | - | - | 1.2 | - | 0.8 | - | - | - | - | 0.9 | - |
| n.d. | 2083 | - | 0.2 | - | 0.3 | 0.9 | - | - | - | - | 1.0 | 0.6 |
| n.d. | 2089 | - | 2.6 | 1.0 | 1.4 | 3.6 | 0.3 | - | - | - | - | 0.7 |
| Phytol | 2100 | 2100 | 3.2 | 1.0 | 1.0 | 2.5 | 1.8 | 2.4 | 4.7 | 1.9 | 6.2 | 5.2 |
| n.d. | 2124 | - | 1.6 | - | 1.4 | 0.9 | 1.5 | 3.0 | 2.1 | 0.6 | 1.2 | 2.7 |
| n.d. | 2190 | - | - | - | - | - | - | - | - | 0.4 | - | 0.6 |
| n.d. | 2258 | - | 0.3 | - | - | - | - | - | - | 2.4 | - | - |
| n.d. | 2336 | - | - | - | - | - | - | - | - | 41.7 | - | 27.5 |
| n.d. | 2346 | - | - | - | - | - | - | - | - | 1.7 | - | - |
| n.d. | 2349 | - | - | - | - | - | - | - | - | 6.5 | - | 2.4 |
| n.d. | 2362 | - | - | - | - | - | - | - | - | 3.4 | - | - |
| n.d. | 2398 | - | - | - | - | - | - | - | - | 15.5 | - | - |
| n.d. | 2471 | - | - | 0.8 | - | - | - | - | - | 1.0 | - | 32.1 |
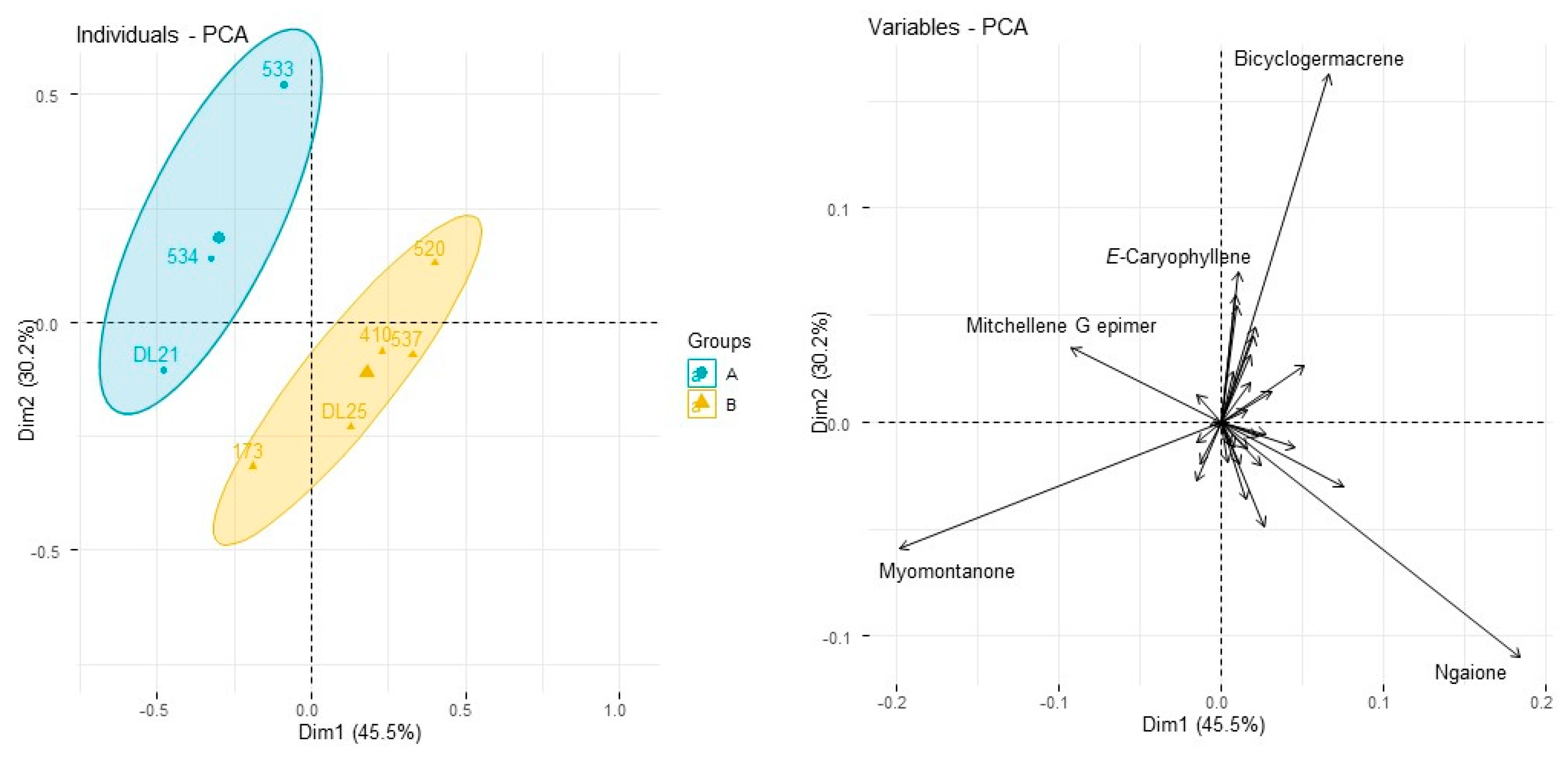
| Specimen Code (See Table 1 and Table 2) | 533 | 534 | 537 | DL-21 ** | 173 | DL-25 ** | 410 | 520 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Yield w/w Wet Leaf | 0.51 | 0.42 | 0.21 | 0.39 | 0.41 | 0.42 | 0.25 | 0.26 | ||
| Compound Name | AI | Pub. AI | ||||||||
| α-Pinene | 933 | 932 | - | - | - | 0.9 | - | 1.9 | 0.7 | 0.7 |
| Unknown | 1327 | nf | - | - | 0.9 | 1.7 | - | 0.8 | 2.2 | 1.1 |
| δ-Elemene | 1337 | 1338 | 1.0 | - | 0.3 | - | - | - | - | 0.6 |
| E-Caryophyllene | 1421 | 1417 | 3.1 | 1.5 | - | - | 0.2 | 0.9 | 1.5 | |
| Aromadendrene | 1440 | 1439 | 1.4 | 0.6 | 0.7 | - | - | - | 0.5 | 0.9 |
| α-Humulene | 1455 | 1452 | 0.8 | - | - | - | - | 0.5 | 0.3 | 0.4 |
| 9-Epi-E-caryophyllene | 1469 | 1464 | 4.2 | 2.0 | - | - | - | 0.2 | 1.1 | 2.1 |
| t-Muurolene | 1484 | 1479 | 1.4 | 0.7 | 6.1 | 1.3 | - | 0.6 | 2.3 | 0.8 |
| Bicyclogermacrene | 1497 | 1500 | 26.1 | 4.7 | 8.0 | 0.9 | - | 0.4 | 6.0 | 14.2 |
| δ-Cadinene | 1523 | 1524 | 0.5 | - | - | - | - | - | - | 0.3 |
| Cedranoxide, 8,14- (isomer) | 1529 | - | 5.3 | 7.5 | 10.2 | 11.5 | 11.2 | 17.3 | 17.6 | 11.8 |
| Cedranoxide, 8,14- | 1545 | 1541 | 3.4 | 5.2 | 5.9 | 6.9 | 7.1 | 10.6 | 10.6 | 6.9 |
| Unknown | 1565 | - | 0.4 | 0.8 | 0.7 | 1.0 | 1.4 | 1.8 | 1.4 | 1.1 |
| Spathulenol | 1578 | 1577 | 10.4 | 5.5 | 6.0 | 1.7 | 4.5 | 3.8 | 3.5 | 7.1 |
| Caryophyllene oxide | 1585 | 1582 | - | 1.5 | 1.1 | - | 1.0 | 0.5 | - | 1.6 |
| Globulol | 1593 | 1590 | 2.4 | 0.9 | 0.8 | - | - | - | - | 0.9 |
| Cedrol | 1603 | 1600 | 0.5 | 1.4 | 1.5 | - | 0.6 | 3.7 | 4.1 | 1.5 |
| Unknown | 1607 | - | - | 1.5 | 1.0 | 2.6 | 1.5 | 0.6 | 0.3 | 1.7 |
| γ-Eudesmol | 1629 | 1630 | - | - | 0.3 | - | 1.1 | 3.2 | 3.0 | 7.1 |
| Myomontanone | 1647 | NMR | 22.6 | 38.1 | 13.6 | 52.8 | 49.8 | 18.9 | 13.1 | 5.0 |
| α-Eudesmol | 1653 | 1652 | 2.0 | - | 3.0 | - | - | 0.8 | 0.6 | 1.0 |
| 7-Epi-α-eudesmol | 1666 | 1662 | - | 0.6 | 0.5 | - | - | 1.1 | 0.2 | 0.5 |
| Mitchellene isomer 1 | 1676 | - | - | 0.8 | 0.9 | 0.9 | - | 0.4 | 2.0 | 0.3 |
| Mitchellene isomer 2 | 1680 | - | - | 1.1 | 0.9 | 1.0 | - | - | 0.7 | 0.2 |
| Ngaione | 1688 | NMR | - | - | 29.4 | - | 11.3 | 11.2 | 12.9 | 15.5 |
| Mitchellene G epimer | 1689 | * | 1.3 | 5.0 | - | 6.4 | - | - | - | - |
| Mitchellene G | 1695 | NMR | 3.9 | 6.3 | 3.5 | 4.3 | 4.5 | 8.2 | 7.1 | 5.2 |
| cis-Nuciferol | 1711 | - | - | 1.4 | 0.9 | - | - | 1.6 | 0.3 | 1.0 |
| trans-Nuciferol | 1725 | 1724 | - | 1.8 | 1.0 | 1.4 | - | 1.5 | 1.0 | - |
| Dehydrongaione | 1752 | NMR | - | - | - | - | - | - | - | 0.8 |
| Myoporone | 1836 | NMR | - | 0.8 | - | - | 0.7 | - | 0.2 | - |
| Mitchellene B | 2083 | NMR *** | 2.7 | 8.5 | 1.6 | 2.0 | 2.0 | 3.1 | 4.3 | 2.3 |
| Species Code (See Table 1 and Table 2) | 93 | 181 | 436 | 541 | Emi-Wood | ||
|---|---|---|---|---|---|---|---|
| Yield g/g Wet Leaves | 0.2 | 0.13 | 0.21 | 0.11 | 1.5 | ||
| Compound Name | AI | Pub. AI | |||||
| α-Pinene | 933 | 934 | 40.1 | - | 38.3 | 52 | - |
| β-Pinene | 977 | 980 | - | - | - | 1.1 | - |
| α-Phellandrene | 1005 | 1007 | 1.9 | - | 1.8 | 7.8 | - |
| p-Cymene | 1024 | 1024 | - | - | 1.3 | - | |
| Limonene | 1028 | 1026 | 1.5 | - | 1.4 | 3.2 | - |
| Terpinolene | 1088 | 1088 | 0.9 | - | 0.9 | 1.9 | - |
| δ-Elemene | 1337 | 1335 | 1.2 | 1.7 | 1.2 | - | - |
| β-Patchoulene | 1382 | 1379 | - | 0.8 | 0.4 | 1.3 | - |
| β-Elemene | 1392 | 1389 | 1.1 | 0.6 | 1.1 | - | |
| α-Gurjunene | 1410 | 1409 | - | 0.7 | 0.6 | 1 | - |
| Caryophyllene | 1420 | 1417 | 1.2 | 1.1 | 1.1 | 1.7 | - |
| 10-Epi-γ-eudesmol | 1428 | 1422 | - | 0.6 | - | 0.7 | - |
| Aromadendrene | 1440 | 1439 | 2.4 | 4.8 | 2.3 | 5.1 | - |
| Alloaromadendrene | 1462 | 1460 | - | 0.7 | - | 0.6 | - |
| γ-Gurjunene | 1487 | 1475 | - | 0.5 | - | - | - |
| β-Selinene | 1489 | 1489 | - | 0.7 | - | - | - |
| Bicyclogermacrene | 1497 | 1500 | 31.3 | 46.8 | 29.8 | 13.7 | - |
| δ-Cadinene | 1523 | 1522 | 1.7 | - | 1.6 | - | - |
| Germacrene B | 1561 | 1559 | - | 0.6 | - | - | - |
| n.d. | 1569 | - | - | 1.2 | 0.6 | - | - |
| Spathulenol | 1578 | 1577 | 6.6 | 14.2 | 6.3 | 3.3 | - |
| Globulol | 1585 | 1590 | 2.6 | 6.8 | 2.5 | 3.1 | - |
| Viridiflorol | 1593 | 1592 | 1.6 | 4.2 | 1.5 | 1.5 | - |
| Guaiol | 1612 | 1600 | - | 1.6 | - | - | 2.5 |
| 10-Epi-γ-eudesmol | 1623 | 1622 | - | 3.9 | - | - | - |
| α-Muurolol | 1639 | 1640 | - | 1.1 | 6.7 | - | - |
| t-Muurolol | 1654 | 1644 | 5.8 | 1.6 | 0.4 | - | - |
| α-Eudesmol | 1657 | 1652 | - | - | - | - | 2.7 |
| Eremophilone | 1740 | 1736 * | - | - | - | - | 50.6 |
| Santalcamphor | 1760 | NMR * | - | - | - | - | 29.1 |
| 8-Hydroxy-1,11-eremophiladien-9-one | 1764 | NMR * | - | - | - | - | 2.8 |
| 9-Hydroxy-7(11),9-eremophiladien-8-one | 1849 | NMR * | - | - | - | - | 11.8 |
| n.d. | 2107 | - | - | 3.8 | - | - | - |
| Species Code (See Table 1 and Table 2) | Eplat | Earb-A | Ego | Ebo | EboL | EboB | EarT | Edal | Edal2 | EspB | EopO | Epur | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Common Name | AI | Pub AI | ||||||||||||
| α-Thujene | 929 | 924 | - | - | - | - | - | - | - | - | - | - | - | 3.5 |
| α-Pinene | 934 | 932 | 1.1 | 3.9 | 7.8 | 1.1 | 3.0 | 9.4 | - | - | - | 20.1 | - | 13.4 |
| Sabinene | 970 | 969 | - | - | 2.0 | - | - | - | - | - | - | 0.9 | - | 10.8 |
| β-Pinene | 975 | 974 | - | - | 2.9 | 2.2 | 6.8 | 12.3 | - | - | - | 10.0 | - | - |
| Myrcene | 992 | 988 | - | - | 5.4 | - | - | 0.9 | - | - | - | 9.4 | - | - |
| p-Cymene | 1026 | 1022 | - | - | - | - | - | - | - | - | - | - | - | 0.9 |
| Limonene | 1028 | 1024 | 2.8 | - | - | - | - | - | - | - | - | 1.4 | - | 14.6 |
| 1,8-Cineole | 1034 | 1026 | - | - | - | - | - | - | - | - | - | - | - | 50.2 |
| Myodesert-1-ene | 1138 | NMR | - | - | - | - | - | - | - | 18.7 | 83.2 | - | - | - |
| β-Cis-ocimene | 1049 | 1032 | 0.3 | 0.8 | - | - | - | - | - | - | - | - | - | - |
| γ-Terpinene | 1059 | 1054 | - | - | - | - | - | - | - | - | - | - | - | 1.3 |
| Terpinen-4-ol | 1180 | 1174 | - | - | - | - | - | - | - | - | - | - | - | 0.6 |
| n.d. | 1238 | - | - | - | - | - | - | - | 6.6 | - | - | - | - | - |
| Dihydronepetalactone | 1276 | - | - | - | - | - | 2.3 | - | 23.1 | 2.2 | - | - | - | - |
| n.d. | 1283 | - | - | - | - | - | - | - | 11.0 | - | - | - | - | - |
| cis,trans-Iridodial-1 | 1291 | - | - | - | - | - | - | - | - | 2.7 | 1.2 | - | - | - |
| cis,trans-Iridodial-2 | 1296 | - | - | - | - | - | - | - | - | 10.3 | 1.6 | - | - | - |
| p-Vinylguaicol | 1311 | 1309 | - | 3.9 | - | - | - | - | 14.2 | - | - | - | 0.8 | - |
| trans,trans-Iridodial | 1314 | - | - | - | - | - | - | - | - | 13.8 | 1.0 | - | - | - |
| n.d. | 1319 | - | - | - | - | - | - | - | 16.4 | - | - | - | - | - |
| cis,cis-Nepetalactol | 1335 | - | - | - | - | - | - | - | - | 20.5 | - | - | - | - |
| Cyclohexene,3-butyl- | 1370 | - | - | - | - | - | - | - | - | 1.0 | - | - | - | - |
| α-Copaene | 1374 | 1374 | - | - | - | 0.8 | - | 2.3 | - | - | - | - | - | - |
| cis,cis-Nepetalactone | 1394 | 1394 | - | - | - | - | - | - | - | 7.5 | 7.7 | - | - | - |
| n.d. | 1406 | - | - | - | - | - | - | - | 9.4 | - | - | - | - | - |
| n.d. | 1414 | - | - | - | - | - | - | - | 7.3 | - | - | - | - | - |
| n.d. | 1419 | - | - | - | - | 0.5 | - | 1.6 | - | - | - | 1.1 | - | - |
| E-Cinnamic acid | 1433 | 1435 | - | 14.8 | - | - | - | - | 0.7 | - | - | - | 31.0 | - |
| 1-Acetoxymyodesert-3-ene | 1457 | NMR | - | - | - | - | - | - | - | 6.8 | 2.8 | - | - | - |
| Veratraldehyde | 1478 | - | - | - | - | - | - | - | - | 9.7 | 0.8 | - | - | - |
| β-Copaene | 1480 | - | - | - | 2.6 | 8.1 | 3.5 | 11.2 | - | - | - | - | - | - |
| Germacrene D | 1485 | 1484 | - | - | - | 8.3 | - | - | - | - | - | 7.1 | - | - |
| n.d. | 1496 | - | - | - | - | 3.3 | 11.1 | 9.5 | - | - | - | - | - | - |
| Germacrene A | 1511 | 1508 | 0.3 | - | - | - | - | - | 2.6 | - | - | - | - | - |
| δ-Cadinene | 1524 | 1522 | - | - | - | 0.4 | - | - | - | - | - | - | - | - |
| n.d. | 1575 | - | 0.4 | - | - | 6.7 | - | 1.4 | - | - | - | - | - | - |
| Spathulenol | 1579 | 1577 | - | - | - | - | - | - | 0.4 | - | - | - | - | - |
| Globulol | 1592 | 1590 | - | - | - | - | - | - | - | - | - | - | - | 0.6 |
| Benzaldehyde, 3,4,5-trimethoxy- | 1601 | - | - | - | - | - | - | - | - | 3.2 | - | - | - | - |
| 10-Epi-7-eudesmol | 1621 | 1622 | - | - | - | - | 4.6 | 8.8 | - | - | - | - | - | - |
| t-Muurolol | 1645 | 1644 | - | - | - | - | - | - | - | - | - | 3.6 | - | - |
| α-Eudesmol | 1657 | 1652 | - | - | - | - | - | 11.1 | 0.3 | - | - | 2.0 | - | - |
| β-Eudesmol | 1665 | 1649 | - | - | - | - | - | - | - | - | - | 6.8 | - | - |
| n.d. | 1683 | - | - | - | - | - | 7.1 | 3.0 | 0.3 | - | - | - | - | - |
| 9-Hydroxydendrolasin | 1740 | NMR | - | 19.0 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 1778 | - | - | - | - | 0.9 | - | 1.5 | - | - | - | - | - | - |
| n.d. | 1787 | - | - | - | - | - | - | 5.5 | - | - | - | - | - | - |
| n.d. | 1825 | - | - | - | - | - | - | - | - | - | - | 9.2 | - | - |
| Myoporone | 1836 | NMR | - | - | - | - | 5.8 | - | 0.4 | - | - | - | - | - |
| n.d. | 1865 | - | - | - | - | - | - | - | - | - | - | 4.6 | - | - |
| n-Hexadecanoic acid | 1964 | 1960 | - | 1.0 | - | 1.1 | - | - | 0.8 | - | - | - | 0.7 | - |
| Hexadecanoic acid | 1995 | 1994 | - | - | - | - | 8.8 | - | - | - | - | - | - | - |
| Freelingnite | 1987 | NMR | - | 36.5 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2006 | - | - | - | - | 30.1 | - | - | - | - | - | - | - | - |
| n.d. | 2017 | - | - | - | - | 3.6 | - | - | - | 1.2 | - | - | - | - |
| n.d. | 2032 | - | - | - | - | 20.5 | - | - | - | - | - | - | - | - |
| n.d. | 2051 | - | - | - | - | - | - | - | - | - | - | - | 21.6 | - |
| n.d. | 2089 | - | - | - | - | 1.8 | 4.4 | - | - | - | - | 5.7 | - | - |
| n.d. | 2100 | - | 1.0 | 1.1 | 13.5 | 5.8 | 37.3 | 19.8 | 1.5 | - | - | 11.1 | 1.9 | 0.6 |
| n.d. | 2124 | - | - | 2.5 | 5.4 | 2.5 | 4.4 | - | 2.3 | 1.1 | - | 4.2 | 5.4 | - |
| Oppositifolic acid | 2149 | NMR ** | 0.3 | - | - | - | - | - | - | - | - | - | 13.0 | - |
| n.d. | 2165 | - | - | 10.0 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2186 | - | 14.9 | - | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2203 | - | 14.6 | - | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2281 | - | - | - | - | - | - | - | - | - | - | - | 21.7 | - |
| n.d. | 2287 | - | 11.8 | - | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2409 | - | - | - | - | - | - | - | 0.6 | - | - | - | - | - |
| n.d. | 2416 | - | - | - | 21.1 | - | - | - | 0.4 | - | - | - | - | - |
| n.d. | 2428 | - | - | - | 35.2 | - | - | - | - | - | - | - | - | - |
| n.d. | 2475 | - | 25.8 | - | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2517 | - | - | 3.4 | - | - | - | - | - | - | - | - | - | - |
| n.d. | 2531 | - | 23.4 | - | - | - | - | - | - | - | - | - | - | - |
| Species Code (See Table 1 and Table 2) | Epte | Ever | Earb-B | Elaa | Ehil | Epur2 | Epai | Esan | Ewel | Erec | Epus | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Common Name | AI | Pub. AI | |||||||||||
| α-Thujene | 929 | 924 | - | - | - | - | - | 6.6 | - | - | - | - | - |
| α-Pinene | 934 | 932 | - | 5.3 | 6.2 | 4.2 | - | 20.2 | - | 0.9 | 67.9 | - | 3.8 |
| Sabinene | 976 | 969 | - | - | - | - | - | 15.0 | - | - | - | - | 76.5 |
| β-Pinene | 981 | 974 | - | 2.0 | - | 22.5 | - | - | - | 6.2 | - | - | - |
| α-Phellandrene | 1010 | 1002 | - | 7.6 | - | - | - | - | - | - | - | - | - |
| β-Phellandrene | 1026 | 1025 | - | 17.5 | - | - | - | 16.0 | - | - | 9.5 | - | 19.7 |
| 1,8-Cineole | 1031 | 1024 | - | - | - | - | - | 42.2 | - | - | - | - | - |
| Methoxymyodesert-3-ene | 1289 | NMR | - | - | - | - | - | - | - | 91.3 | - | - | - |
| 2-Methoxy-4-vinylphenol | 1322 | 1315 | - | - | - | - | 31.4 | - | - | - | 6.4 | - | - |
| cis,cis-Nepetalactone | 1395 | 1391 | - | - | - | - | - | - | - | 1.6 | - | - | - |
| Alloaromadendrene | 1454 | 1458 | - | - | - | - | 18.8 | - | - | - | - | 3.5 | - |
| Germacrene D | 1500 | 1488 | - | - | - | - | - | - | - | - | - | 17.1 | - |
| Bicyclogermacrene | 1512 | 1500 | - | 9.1 | - | - | - | - | - | - | - | 7.6 | - |
| n.d. | 1529 | - | - | - | - | - | - | - | - | - | - | 10.1 | - |
| n.d. | 1533 | - | - | - | - | - | - | - | - | - | - | 6.1 | - |
| Elemol | 1553 | 1548 | - | - | - | - | 6.5 | - | 80.3 | - | - | - | - |
| Germacrene D-4-ol | 1574 | 1576 | - | - | - | - | - | - | - | - | - | 26.9 | - |
| Spathulenol | 1577 | 1577 | - | - | - | - | - | - | - | - | 3.9 | - | - |
| n.d. furanosesquiterpene | 1592 | - | - | - | - | 41.5 | - | - | - | - | - | - | - |
| γ-Eudesmol | 1627 | 1630 | - | - | - | - | - | - | 4.5 | - | - | - | - |
| Eremoacetal | 1630 | NMR | - | 33.3 | - | - | - | - | - | - | - | 14.1 | - |
| t-Muurolol | 1635 | 1640 | - | - | - | - | - | - | - | - | 3.9 | - | - |
| Ngaione | 1639 | NMR | 4.1 | - | - | 4.3 | - | - | - | - | - | - | - |
| n.d. furanosesquiterpene | 1654 | - | - | - | - | 13.1 | - | - | - | - | - | - | - |
| Dehydrongaione | 1677 | NMR | 8.5 | 2.7 | - | 11.3 | - | - | - | - | - | - | - |
| Ngaiol ketol * | 1699 | ** | - | 17.8 | - | 3.1 | - | - | - | - | - | - | - |
| n.d. | 1708 | - | - | - | - | - | - | - | 4.1 | - | 3.6 | - | - |
| 9-Hydroxydendrolasin | 1740 | NMR | - | - | 8.2 | - | - | - | 4.1 | - | 4.8 | - | - |
| n.d. furanosesquiterpene | 1771 | - | 4.1 | - | - | - | - | - | 7.0 | - | - | - | - |
| n.d. furanosesquiterpene | 1814 | - | - | - | - | - | - | - | - | - | - | 5.6 | - |
| Myoporone | 1840 | NMR | - | 4.7 | - | - | - | - | - | - | - | 9.0 | - |
| Dehymyoporone | 1901 | NMR | 11.1 | - | - | - | - | - | - | - | - | - | - |
| Phytol | 1928 | 1930 | - | - | - | - | 36.7 | - | - | - | - | - | - |
| n.d. furanosesquiterpene | 1912 | - | 3.6 | - | - | - | - | - | - | - | - | - | - |
| n.d. furanosesquiterpene | 1932 | - | 49.1 | - | - | - | - | - | - | - | - | - | - |
| n.d. furanosesquiterpene | 1935 | - | 10.0 | - | - | - | - | - | - | - | - | - | - |
| n.d. furanosesquiterpene | 1944 | - | 9.5 | - | - | - | 6.6 | - | - | - | - | - | - |
| Freelingnite | 1987 | NMR | - | - | 85.6 | - | - | - | - | - | - | - | - |
| Species Code (See Table 1 and Table 2) | Emi-436 | Egi-518 | EopR-540 | EopO-538 B | EopR-535 | EopR-539 | ||
|---|---|---|---|---|---|---|---|---|
| Common Name | AI | Pub AI | ||||||
| α-Thujene | 928 | 924 | - | - | - | 1.0 | - | - |
| α-pinene | 933 | 932 | 28.6 | 9.2 | - | - | 1.5 | - |
| 2E-Heptenal | 941 | 947 | - | 0.6 | 0.9 | 0.6 | 1.6 | 2.8 |
| Camphene | 946 | 946 | - | - | - | 0.6 | - | 1.4 |
| β-Pinene | 975 | 974 | - | 0.5 | - | - | - | - |
| Myrcene | 992 | 988 | - | 9.2 | - | - | - | - |
| Butyl butanoate | 994 | 993 | - | - | - | 5.5 | - | - |
| n-Decane | 997 | 1000 | - | - | - | 1.5 | - | - |
| α-Phellandrene | 1005 | 1002 | 1.7 | 0.6 | - | 0.6 | - | - |
| α-Terpinene | 1019 | 1014 | - | - | 1.3 | - | 1.2 | 0.8 |
| β-Phellandrene | 1025 | 1025 | 0.9 | - | - | - | - | - |
| p-Cymene | 1026 | 1022 | - | 0.5 | - | - | - | - |
| Limonene | 1028 | 1024 | - | 1.5 | - | - | - | - |
| β-Ocimene | 1048 | 1044 | - | 0.6 | - | 0.8 | - | - |
| n-Undecane | 1097 | 1100 | - | - | - | 1.0 | - | - |
| Karahanaenone | 1154 | 1154 | - | 2.4 | - | - | - | - |
| Isomenthone | 1162 | 1158 | - | 0.7 | - | - | - | - |
| Hexyl butyrate | 1190 | 1191 | - | - | - | 4.8 | - | - |
| p-Vinylguaiacol | 1313 | 1309 | 4.9 | - | - | - | - | - |
| δ-Elemene | 1337 | 1335 | 1.0 | - | - | - | - | - |
| α-Copaene | 1375 | 1374 | - | - | - | - | 1.2 | - |
| β-Elemene | 1396 | 1389 | - | 1.6 | - | - | - | - |
| β-Duprezianene | 1418 | 1421 | - | 0.6 | 0.8 | - | - | - |
| E-Caryophyllene | 1425 | 1417 | - | - | - | 1.3 | 1.4 | - |
| 1-Amorpha-4,7(ll)-diene | 1480 | 1479 | - | 1.7 | - | - | - | - |
| Z-β-Guaiene | 1485 | 1492 | - | 0.5 | - | - | - | - |
| Bicydogermacrene | 1502 | 1500 | 18.8 | 4.4 | - | - | 1.5 | - |
| Germacrene A | 1511 | 1508 | - | 1.7 | - | - | - | - |
| δ-Cadinene | 1522 | 1522 | - | 2.3 | - | 0.7 | 3.1 | - |
| 8,14-Cedranoxide | 1544 | 1541 | - | - | - | 0.7 | - | - |
| n.d. | 1568 | - | - | 0.8 | - | - | - | - |
| Spathulenol | 1579 | 1577 | 3.5 | 0.7 | - | 0.7 | - | - |
| n.d. | 1584 | - | - | 1.2 | - | - | - | - |
| Humulene epoxide II | 1611 | 1608 | - | - | - | 1.2 | - | - |
| t-Muurolol | 1645 | 1644 | - | - | - | - | 1.3 | 1.1 |
| β-Eudesmol | 1653 | 1649 | 1.6 | - | - | - | - | - |
| Z-11-Hydroxyisodendrolasin | 1678 | NMR | - | - | - | 2.6 | - | - |
| Epignaione | 1694 | NMR | - | - | - | 1.0 | - | - |
| 9-Hydroxydendrolasin | 1741 | NMR | - | - | - | 0.7 | - | - |
| 10R-Hydroxydihydro-α-humulene acetate | 1755 | NMR | - | - | - | 3.1 | - | - |
| 8-Cedren-13-ol acetate | 1788 | 1788 | - | - | 1.1 | - | 1.0 | 0.5 |
| n.d. | 1895 | - | - | - | 2.3 | - | - | 0.6 |
| n.d. | 2100 | - | 5.4 | - | - | - | - | - |
| Oppositifolic acid | 2149 | NMR ** | - | - | 69.2 | 69.8 | 84.6 | 56.2 |
| n.d. | 2260 | - | - | - | - | - | - | 0.7 |
| n.d. | 2269 | - | - | - | - | - | - | 2.2 |
| n.d. | 2270 | - | - | 11.7 | - | - | - | |
| n.d. chain 1 | 2272 | - | - | - | 10.0 | - | - | 0.7 |
| n.d. | 2318 | - | 15.6 | - | - | - | - | - |
| n.d. chain 2 | 2359 | - | - | - | 12.7 | - | - | 30.7 |
| n.d. isomer 2 | 2412 | - | - | 46.8 | - | - | - | - |
| n.d. | 2449 | - | 6.7 | - | - | - | - | - |
| n.d. | 2471 | - | 11.2 | - | - | - | - | - |
2.4. Miscellaneous Eremophila by Endemic Location
2.4.1. Central and South Australian Eremophila
2.4.2. Eremophila of the Murchison District, Western Australia
2.4.3. South-Western Australian Eremophila (Kalgoorlie Region)
2.4.4. Central Lowlands Region
3. Materials and Methods
3.1. Specimen Collection
3.2. Production of Essential Oils
3.3. Chromatography Methods and Compound Identification (GC–MS and NMR Analysis)
3.4. Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chinnock, R. Eremophila and Allied Genera. A Monograph of the Myoporaceae; Rosenberg Publishing: Kenthurst, Australia, 2007. [Google Scholar]
- Sadgrove, N.J.; Jones, G.L. Cytogeography of essential oil chemotypes of eremophila longifolia f. Muell (schrophulariaceae). Phytochemistry 2014, 105, 43–51. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Collins, T.L.; Legendre, S.V.A.-M.; Klepp, J.; Jones, G.L.; Greatrex, B.W. The iridoid myodesert-1-ene and elemol/eudesmol are found in distinct chemotypes of the australian aboriginal medicinal plant eremophila dalyana(scrophulariaceae). Natl. Prod. Commun. 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Sadgrove, N.J.; Klepp, J.; Legendre, S.V.A.-M.; Lyddiard, D.; Sumby, C.J.; Greatrex, B.W. Revision of the phytochemistry of eremophila sturtii and e. mitchellii. J. Natl. Prod. 2018, 81, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.E.; Tucker, D.J.; Alter, D.; Watson, K.; Jones, G.L. Intraspecific variation in essential oil composition of eremophila longifolia f. Muell (myoporaceae): Evidence for three chemotypes. Phytochemistry 2010, 71, 1521–1527. [Google Scholar] [CrossRef]
- Sadgrove, N.; Mijajlovic, S.; Tucker, D.J.; Watson, K.; Jones, G.L. Characterization and bioactivity of essential oils from novel chemotypes of eremophila longifolia (f. Muell) (myoporaceae): A highly valued traditional australian medicine. Flavour Fragr. J. 2011, 26, 341–350. [Google Scholar] [CrossRef]
- Beattie, K.; Waterman, P.G.; Forster, P.I.; Thompson, D.R.; Leach, D.N. Chemical composition and cytotoxicity of oils and eremophilanes derived from various parts of Eremophila mitchellii benth. (myoporaceae). Phytochemistry 2011, 71, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadgrove, N.J.; Jones, G.L. A contemporary introduction to essential oils: Chemistry, bioactivity and prospects for australian agriculture. Agriculture 2015, 5, 48–102. [Google Scholar] [CrossRef] [Green Version]
- Barlow, B.A. Cytogeography of the genus eremophila. Aust. J. Bot. 1971, 19, 295–310. [Google Scholar] [CrossRef]
- Williams, N.J.; Harle, K.J.; Gale, S.J.; Heijnis, H. The vegetation history of the last glacial-interglacial cycle in eastern new south wales, australia. J. Quat. Sci. 2006, 21, 735–750. [Google Scholar] [CrossRef]
- Singab, A.N.; Youssef, F.S.; Ashour, M.L.; Wink, M. The genus eremophila (scrophulariaceae): An ethnobotanical, biological and phytochemical review. J. Pharm. Pharmacol. 2013, 65, 1239–1279. [Google Scholar] [CrossRef] [PubMed]
- Grant, H.G.; O’Regan, P.J.; Park, R.J.; Sutherland, M.D. Terpenoid chemistry. XXIV. (1r)-1-methoxymyodesert-3-ene, an iridoid constituent of myoporum deserti (myoporaceae). Aust. J. Chem. 1980, 33, 853–878. [Google Scholar] [CrossRef]
- Hegarty, B.F.; Kelly, J.R.; Park, R.J.; Sutherland, M.D. Terpenoid chemistry: XVII. (-)-gnaione, a toxic constituent of myoporum deserti. The absolute configuration of (-)-ngaione. Aust. J. Chem. 1970, 23, 107–117. [Google Scholar] [CrossRef]
- Usuki, Y.; Deguchi, T.; Iio, H. A new concise synthesis of (+)-ipomeamarone, (−)-ngaione, and their stereoisomers. Chem. Lett. 2014, 43, 1882–1884. [Google Scholar] [CrossRef]
- Hamilton, W.D.; Park, R.J.; Perry, G.J.; Sutherland, M.D. XXI. (-)-epignaione, (-)-dehydrongaione, (-)-dehydroepingaione, and (-)-deisopropylngaione, toxic furanoid sesquiterpenoid ketones from myoporum deserti. Aust. J. Chem. 1973, 26, 375–387. [Google Scholar] [CrossRef]
- Grant, H.G.; Russell-Maynard, C.A.; Sutherland, M.D. Terpenoid chemistry. XXVII further iridoid constituents (1r)- and (1s)-1-acetoxymyodesert-3-ene, from myoporum deserti (myoporaceae). Aust. J. Chem. 1985, 38, 325–336. [Google Scholar] [CrossRef]
- Blackburne, I.D.; Sutherland, M.D. Terpenoid chemistry xix. Dehydro- and dehydroiso-myodesmone, toxic furanoid sesquiterpene ketones from myoporum deserti. Aust. J. Chem. 1972, 25, 1779–1786. [Google Scholar] [CrossRef]
- Blackburne, I.D.; Park, R.J.; Sutherland, M.D. Terpenoid chemistry: XX. Myoporone and dehydromyoporone, toxic furanoid ketones from myoporum and eremophila species. Aust. J. Chem. 1972, 25, 1787–1796. [Google Scholar] [CrossRef]
- Sutherland, M.D.; Rodwell, J.L. Terpenoid chemistry. XXVIII furanosesquiterpene β-ketols from myoporum betcheanum, m. Deserti, m. Montanum and other myoporaceae. Aust. J. Chem. 1989, 42, 1995–2019. [Google Scholar] [CrossRef]
- Richmond, G.S. A review of the use of eremophila (myoporaceae) by australian aborigines. J. Adel. Bot. Gard. 1993, 15, 101–107. [Google Scholar]
- Ghisalberti, E.L. The ethnopharmacology and phytochemistry of eremophila species (myoporaceae). J. Ethnopharmacol. 1994, 44, 1–9. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Jones, G.L. Reviewing the importance of aromatic medicinal plants in the traditional pharmacopoeia of australian aboriginal people. Acta Hortic. 2016. [Google Scholar] [CrossRef]
- Sadgrove, N.J. Southern africa as a ‘cradle of incense’ in wider african aromatherapy Sci. Afr. 2020, 9, e00502. [Google Scholar]
- Langat, M.K.; Mayowa, Y.; Sadgrove, N.J.; Danyaal, M.; Prescott, T.A.K.; Kami, T.; Schwikkard, S.; Barker, J.; Cheek, M. Multi-layered antimicrobial synergism of (e)-caryophyllene with alkaloids and 13s-hydroxy-9z,11e,15e-octadecatrienoic acid from the leaves of vepris gossweileri (i. Verd.) mziray. Natl. Prod. Res. 2021, in press. [Google Scholar] [CrossRef]
- Nsangou, M.F.; Happi, E.N.; Fannang, S.V.; Atangana, A.F.; Waffo, A.F.K.; Wansi, J.D.; Isyaka, S.M.; Sadgrove, N.J.; Sewald, N.; Langat, M.K. Chemical composition and synergistic antimicrobial effects of a vegetatively propagated cameroonian lemon, citrus x limon (l.) osbeck. ACS Food Sci. Technol. 2021, in press. [Google Scholar] [CrossRef]
- Ndi, C.P.; Semple, S.J.; Griesser, H.J.; Pyke, S.M.; Barton, M.D. Antimicrobial compounds from the australian desert plant eremophila neglecta. J. Natl. Prod. 2007, 70, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Mon, H.H.; Christo, S.N.; Ndi, C.P.; Jasieniak, M.; Rickard, H.; Hayball, J.D.; Griesser, H.J.; Semple, S.J. Serrulatane diterpenoid from eremophila neglecta exhibits bacterial biofilm dispersion and inhibits release of pro-inflammatory cytokines from activated macrophages. J. Natl. Prod. 2015, 78, 3031–3040. [Google Scholar] [CrossRef]
- Smith, J.E.; Tucker, D.; Watson, K.; Jones, G.L. Identification of antibacterial constituents from the indigenous australian medicinal plant eremophila duttonii f. Muell. (myoporaceae). J. Ethnopharmacol. 2007, 112, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Lyddiard, D.; Greatrex, B.W. Serrulatic acid diastereomers identified from an antibacterial survey of eremophila. Fitoterapia 2018, 126, 29–34. [Google Scholar] [CrossRef]
- Vasilev, K.; Cook, J.; Griesser, H.J. Antibacterial surfaces for biomedical devices. Expert Rev. Med. Dev. 2009, 6, 553–567. [Google Scholar] [CrossRef]
- Kong, E.F.; Tsui, C.; Kuchariková, S.; Dijck, P.V.; Jabra-Rizk, M.A. Modulation of staphylococcus aureus response to antimicrobials by the candida albicans quorum sensing molecule farnesol. Antimicrob. Agents Chemother. 2017, 61, e01573-17. [Google Scholar] [CrossRef] [Green Version]
- Carman, R.M.; Handley, P.N.; Kadirvelraj, R.; Robinson, W.T.; Sutherland, M.D. The stereochemical structures of the furanosesquiterpenoidal β-hydroxy ketones from the myoporaceae. Aust. J. Chem. 1999, 52, 727–732. [Google Scholar]
- von der Ohe, F.; Bruckner, R. Z- or e-configurated γ-alkylidenebutenolides from a 2-(trimethylsiloxy)furan and iodomethacrolein-stereoselective synthesis of z- and e-freelingyne. Tetrahedron Lett. 1998, 39, 1909–1910. [Google Scholar] [CrossRef]
- Knight, D.W.; Pattenden, G. Freelingnite, a new furanosesquiterpene from eremophila freelingii. Tetrahedron Lett. 1975, 13, 1115–1116. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Telford, I.R.H.; Padilla-González, G.F.; Greatrex, B.W.; Bruhl, J.J. Gc–ms ‘chemophenetics’ on australian pink-flowered phebalium (rutaceae) using herbarium leaf material demonstrates phenetic agreement with putative new species. Phytochem. Lett. 2020, 38, 112–120. [Google Scholar] [CrossRef]
- Toyota, M.; Koyama, H.; Mizutani, M.; Asakawa, Y. (−)-ent-spathulenol isolated from liverworts is an artefact. Phytochemistry 1996, 41, 1347–1350. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Lyddiard, D.; Collins, T.L.; Greatrex, B.W.; Jones, G.L. Genifuranal and other derivatives: Smoking desert plants. Acta Hortic. 2016, 1125, 181–188. [Google Scholar] [CrossRef]
- Sadgrove, N.; Jones, G.L.; Greatrex, B.W. Isolation and characterisation of (-)-genifuranal: The principal antimicrobial component in traditional smoking applications of eremophila longifolia (scrophulariaceae) by australian aboriginal peoples. J. Ethnopharmacol. 2014, 154, 758–766. [Google Scholar] [CrossRef]
- Sadgrove, N.J. Comparing essential oils from australia’s ‘victorian christmas bush’ (prostanthera lasianthos labill., lamiaceae) to closely allied new species: Phenotypic plasticity and taxonomic variability. Phytochemistry 2020, 176, 112403. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-González, G.F.; Telford, I.R.H.; Greatrex, B.W.; Jones, G.L.; Andrew, R.; Bruhl, J.J.; Langat, M.K.; Melnikovova, I.; Fernandez-Cusimamani, E. Prostanthera (lamiaceae) as a ‘cradle of incense’: Chemophenetics of rare essential oils from both new and forgotten australian ‘mint bush’ species. Plants 2020, 9, 1570. [Google Scholar] [CrossRef]
- Barr, A. Traditional Bush Medicines: An Aboriginal Pharmacopoeia; Greenhouse Publications Pty Ltd.: Richmond, Australia, 1988. [Google Scholar]
- Latz, P. Bushfires and Bushtucker: Aboriginal Plant Use in Central Australia; IAD Press: Alice Springs, Australia, 2004. [Google Scholar]
- Odonnell, G.W.; Sutherland, M.D. Terpenoid chemistry. Xxix.(-)-anymol from myoporum crassifolium forst, the c7 epimer of (-)-α-bisabolol from camomile. Aust. J. Chem. 1989, 42, 2021–2034. [Google Scholar] [CrossRef]
- Liu, Q.; Harrington, D.; Kohen, J.L.; Vemulpad, S.; Jamie, J.F. Bactericidal and cyclooxygenase inhibitory diterpenes from eremophila sturtii. Phytochemistry 2006, 67, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.C.; Carroll, A.R.; Davis, R.A. Mitchellene a-e, cyclic sesquiterpenes from the australian plant eremophila mitchellii. J. Natl. Prod. 2011, 74, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Burka, L.T.; Felice, L.J.; Jackson, S.W. 6-oxodendrolasin, 6-hydroxydendrolasin, 9-oxofarnesol and 9-hydroxyfarnesol, stress metabolites of the sweet potato. Phytochemistry 1981, 20, 647–652. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. The chemistry of unusual terpenoids from the genus eremophila. In Studies in Natural Product Chemistry; Atta-ur-Rahman, Ed.; Elsevier Science: Amsterdam, The Netherlands, 1995; Volume 15, Part C; pp. 225–287. [Google Scholar]
- Jefferies, P.R.; Knox, J.R. The chemistry of eremophila species: Ii. A novel unsaturated fatty acid from e. Oppositifolia r.Br. Aust. J. Chem. 1961, 14, 628–636. [Google Scholar] [CrossRef]
- Mosaddik, A.; Waterman, P.G. A sesquiterpene, clerodane diterpenes and a furanone from the roots of casearia multinervosa (flacourtiaceae/salicaceae). Natl. Prod. Commun. 2006, 1, 601–607. [Google Scholar] [CrossRef]
- Simionatto, E.; Porto, C.; Dalcol, I.I.; da Silva, U.F.; Morel, A.F. Essential oil from zanthoxylum hyemale. Planta Med. 2005, 71, 759–763. [Google Scholar] [CrossRef]
- AVH. Australasian Virtual Herbarium. Available online: https://avh.chah.org.au/ (accessed on 25 March 2020).
- Yamauchi, S.; Yamaguchi, M. Synthesis of (+)-aptosimon, a 4-oxofurofuran lignan, by erythro selective aldol condensation and stereoconvergent cyclization as the key reactions. Biosci. Biotechnol. Biochem. 2003, 67, 838–846. [Google Scholar] [CrossRef] [Green Version]
- Algreiby, A. Isolation and Identification of Bioactive Compounds from eremophila glabra and biserrula pelecinus. Ph.D. Thesis, University of Western Australia, Perth, Australia, 2018. [Google Scholar] [CrossRef]
- Russell, C.A.; Sutherland, M.D. Terpenoid chemistry. XXV the absolute configuration of (-)-ngaione, (-)-epingaione and related substances. Aust. J. Chem. 1982, 35, 1881–1894. [Google Scholar] [CrossRef]
- Rajeswara, R.B.R. Hydrosols and water-soluble essential oils: Their medicinal and biological properties. In Recent progress in medicinal plants: Essential oils I; Studium Press LLC.: Houston, TX, USA, 2013; pp. 119–140. [Google Scholar]
- NIST. Nist Chemistry Webbook: Nist Standard Reference Database Number 69. Available online: https://webbook.nist.gov/chemistry/ (accessed on 16 January 2021).
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 978-1-932633-21-4. [Google Scholar]
- Carman, R.M.; Duffield, A.R. (+)-a-bisabolol and (+)-anymol. A repetition of the synthesis from the limonene 8,9-epoxides Aust. J. Chem. 1989, 42, 2035–2039. [Google Scholar]
- Ding, L.; Pfoh, R.; RuhL, S.; Qin, S.; Laatsch, H. T-muurolol sesquiterpenes from the marine streptomyces sp. M491 and revision of the configuration of previously reported amorphanes1. J. Natl. Prod. 2009, 72, 99–101. [Google Scholar] [CrossRef]
- Jefford, C.W.; Sledeski, A.W.; Rossier, J.-C.; Boukouvalas, J. A short route to furanosesquiterpenes using a new siloxyfuran building block. The synthesis of freelingnite and dehydrolasiosperman. Tetrahedron Lett. 1990, 31, 5741–5744. [Google Scholar] [CrossRef]
- Blackburne, I.D.; Park, R.J.; Sutherland, M.D. Terpenoid chemistry: XVIII. Myodesmone and isomyodesmone, toxic furanoid ketones from myoporum deserti and m. Acuminatum. Aust. J. Chem. 1974, 24, 995–1007. [Google Scholar] [CrossRef]
- Metra, P.L.; Sutherland, M.D. Further skeletal variety in the toxic furanosesquiterpene ketones in the myoporum genus. Tetrahedron Lett. 1983, 24, 1749–1752. [Google Scholar] [CrossRef]
- Dimitriadis, E.; Massy-Westropp, R.A. The structure of eremoacetal, a sesquiterpene from eremophila rotundifolia. Aust. J. Chem. 1979, 32, 2003–2015. [Google Scholar] [CrossRef]
- Nordin, O.; Hedenstrom, E.; Hogberg, H.-E. Stereochemistry of 1,6-germacradien-5-ol, a constituent of the needles of scots pine (pinus sylvestris) and of the defence secretion from larvae of the pine sawfly neodiprion sertifer. Acta Chem. Scand. 1999, 53, 124–132. [Google Scholar] [CrossRef] [Green Version]
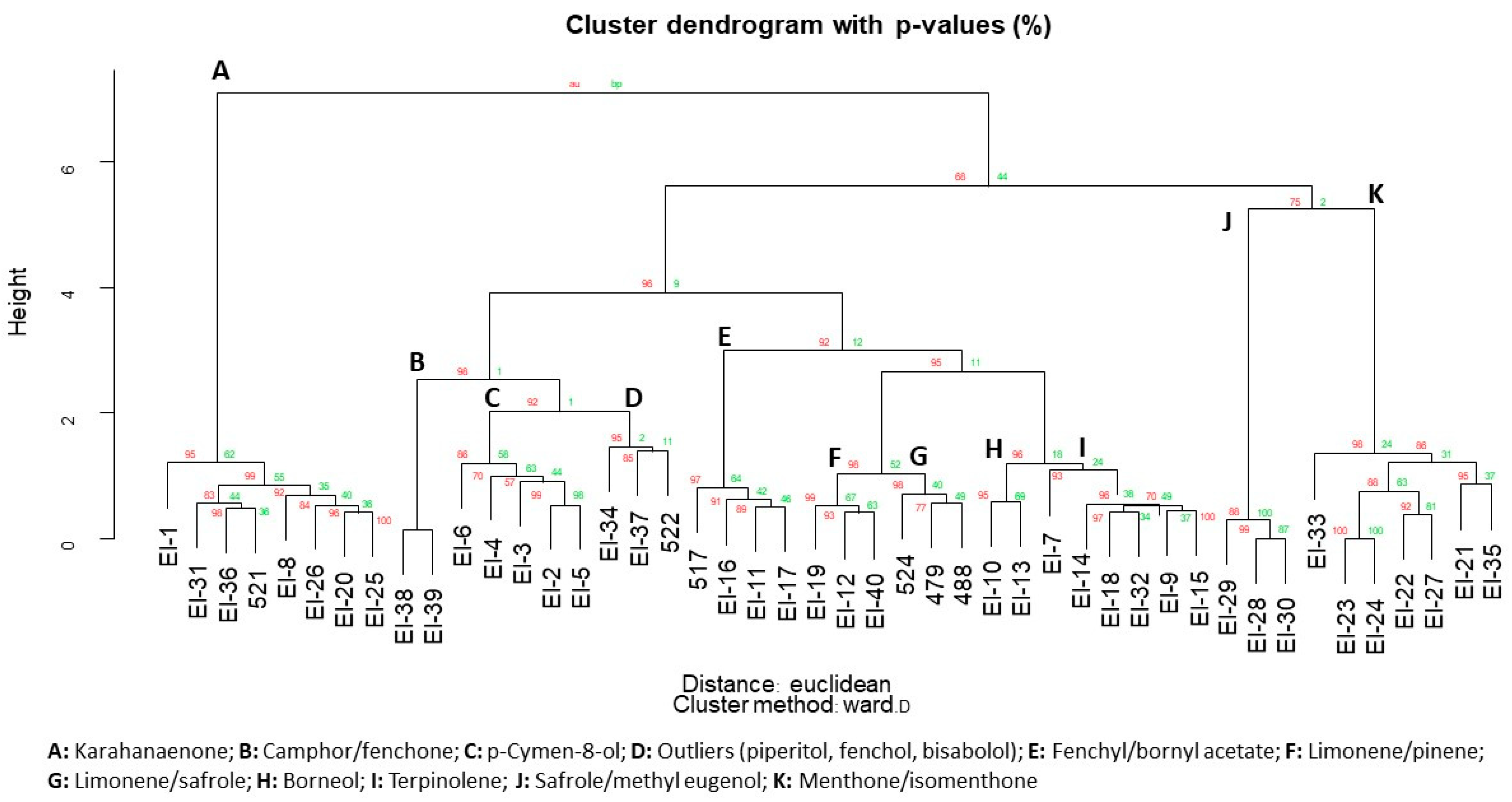
- Suzuki, R.; Shimodaira, H. Pvclust: An r package for assessing the uncertainty in hierarchical clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef] [PubMed]
- Padilla-González, G.F.; Aldana, J.A.; Costa, F.B.D. Chemical characterization of two morphologically related espeletia (asteraceae) species and chemometric analysis based on essential oil components. Rev. Bras. Farmacogn. 2016, 26, 697–700. [Google Scholar] [CrossRef] [Green Version]
- Padilla-González, G.F.; Frey, M.; Gómez-Zeledón, J.; Da Costa, F.B.; Spring, O. Metabolomic and gene expression approaches reveal the developmental and environmental regulation of the secondary metabolism of yacón (smallanthus sonchifolius, asteraceae). Sci. Rep. 2019, 9, 13178. [Google Scholar] [CrossRef] [PubMed]









| Species | Abr. | Location | Species | Abr. | Location |
|---|---|---|---|---|---|
| E. pterocarpa * | Epte | Murchison, WA | E. goodwinnii | Ego | Private Garden—Inverell, NSW |
| E. veronica * | Ever | Kalgoorlie, WA | E. spectabilis subsp. brevis | EspB | Private Garden—Inverell, NSW |
| E. laanii * | Elaa | Murchison, WA | E. purpurascens | Epur2 | Private Garden—Inverell, NSW |
| E. hillii * | Ehil | SA-WA border, Great Australian Bight | E. bowmannii | Ebo | Private Garden—Inverell, NSW |
| E. purpurascens * | Epur | Kalgoorlie, WA | E. bowmannii subsp. bowmannii | EboB | Private Garden—Inverell, NSW |
| E. paisleyi * | Epai | Alice Springs – Port Augusta | E. bowmannii subsp. latifolia | EboL | Private Garden—Inverell, NSW |
| E. santalina * | Esan | Port Augusta, SA | E. arachnoides subsp. tenera | EarT | Private Garden—Inverell, NSW |
| E. weldii * | Ewel | SA-WA border, Great Australian Bight | E. dalyana | Edal | Private Garden—Inverell, NSW |
| E. recurva * | Erec | Gascoyne Junction, near Murchison, WA | E. dalyana | Edal2 | Private Garden—Inverell, NSW |
| E. pustulata * | Epus | Kalgoorlie, WA | E. arbuscular | Earb | Private Garden—Inverell, NSW |
| E. platycalyx | Epla | Private Garden—Inverell, NSW | E. arbuscular | Earb2 | Private Garden—Inverell, NSW |
| E. oppositifolia subsp. oppositifolia | EopO | Private Garden—Inverell, NSW | E. freelingii | Efre-338A | Alice Springs, NT |
| E. oppositifolia subsp. rubra | EopR-535 | Broken Hill to Wiilcannia, NSW | E. freelingii | Efre-338B | Alice Springs, NT |
| E. oppositifolia subsp. rubra | EopR-538B | Broken Hill to Wiilcannia, NSW | E. freelingii | Efre-338C | Alice Springs, NT |
| E. oppositifolia subsp. rubra | EopR-539 | Broken Hill to Wiilcannia, NSW | E. freelingii | Efre-338D | Alice Springs, NT |
| E. oppositifolia subsp. rubra | EopR-540 | Broken Hill to Wiilcannia, NSW | E. alternifolia X Myoporum montanum | EaltMm | Private Garden—Inverell |
| E. gilesii | Egil-518 | SW Qld | E. alternifolia | Ealt-A | Private Garden—Inverell |
| E. latrobei subsp. latrobei | ElatL | Private Garden—Inverell, NSW | E. alternifolia | Ealt-B | Private Garden—Inverell |
| E. latrobei subsp. glabra | ElatG | Private Garden—Inverell, NSW | E. alternifolia | Ealt-C | Private Garden—Inverell |
| E. latrobei subsp. filiform | ElatF | Private Garden—Inverell, NSW | E. alternifolia subsp. latifolia | EaltL-A | Private Garden—Inverell |
| E. deserti | Ede | Private Garden—Inverell, NSW | E. alternifolia subsp. latifolia | EaltL-B | Private Garden—Inverell |
| E. mitchellii | Emi-436 | SE Qld | E. alternifolia subsp. latifolia | EaltL-C | Private Garden—Inverell |
| Species | Collector No. | Location | Species | Collector No. | Location |
|---|---|---|---|---|---|
| E. gilesii (Egi) | 341 | Alice Springs, NT | E. alternifolia (Ealt) | 170 | Broken Hill, NSW |
| E. platycalyx (Epla) | 53 | Sandstone, WA | 261 | Broken Hill, NSW | |
| 61 | Mt Magnet, WA | 408 | Broken Hill, NSW | ||
| 62 | Mt Magnet, WA | E. longifolia (Elo) | 517 | Cunnamulla, Qld | |
| E. deserti (Ede) | 105 | Emerald, Qld | 521 | Cunnamulla, Qld | |
| 470 | Blackall, Qld | ||||
| 471 | Blackall, Qld | 522 | Grey Ranges, Thargomindah, Qld | ||
| 494A | Miles, Qld | 524 | Cunnamulla, Qld | ||
| 494B | Miles, Qld | 479 | Blackall, Qld | ||
| 494C | Miles, Qld | 488 | Blackall, Qld | ||
| 507B | Goondawindi, Qld | E. neglecta (Ene) | Private Land | SA border to NT | |
| 509-May * | Moonie, SW Qld | E. arbuscular (Earb) | 487 | Idalia, NP, Qld | |
| 510-Oct * | Moonie, SW Qld | 486 | Idalia, NP, Qld | ||
| 519A | Cunnamulla, Qld | E. oppositifolia subsp. rubra (EopR) and subsp. oppositifolia (EopO) | EopR-535B | Wilcannia, NSW | |
| 519B | Cunnamulla, Qld | EopO-538A | Broken Hill, NSW | ||
| 519C | Cunnamulla, Qld | E. sturtii (Est) | 533 | Cobar, NSW | |
| 519E | Cunnamulla, Qld | 534 | Cobar, NSW | ||
| 536A | Broken Hill, NSW | 537 | Wilcannia, NSW | ||
| 536B | Broken Hill, NSW | DL-21 ** | Broken Hill, NSW | ||
| 536C | Broken Hill, NSW | 173 | Mutawintji NP, NSW | ||
| E. mitchellii (Emi) | 93 | North Star, NSW | DL-25 ** | Mutawintji NP, NSW | |
| 410 | Mutawintji NP, NSW | ||||
| 181 | Warren, NSW | 520 | Cunnamulla, Qld | ||
| 436 | Collarenebri, NSW | E. latrobei (Elat) | 269 | Mutawintji NP, NSW | |
| 541 | Wilcannia, NSW | 337 | Coober Pedy, NT | ||
| Emi-Wood EO | Private Collection | E. youngii (Eyo) | 345 | Alice Springs, NT | |
| E. foliosissima (Efo) | 57 | Sandstone, WA | E. freelingii (Efre) | 346 | Alice Springs, NT |
| E. duttonii (Edut) | DL-27 ** | Mutawintji NP NSW | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadgrove, N.J.; Padilla-González, G.F.; Green, A.; Langat, M.K.; Mas-Claret, E.; Lyddiard, D.; Klepp, J.; Legendre, S.V.A.-M.; Greatrex, B.W.; Jones, G.L.; et al. The Diversity of Volatile Compounds in Australia’s Semi-Desert Genus Eremophila (Scrophulariaceae). Plants 2021, 10, 785. https://doi.org/10.3390/plants10040785
Sadgrove NJ, Padilla-González GF, Green A, Langat MK, Mas-Claret E, Lyddiard D, Klepp J, Legendre SVA-M, Greatrex BW, Jones GL, et al. The Diversity of Volatile Compounds in Australia’s Semi-Desert Genus Eremophila (Scrophulariaceae). Plants. 2021; 10(4):785. https://doi.org/10.3390/plants10040785
Chicago/Turabian StyleSadgrove, Nicholas J., Guillermo F. Padilla-González, Alison Green, Moses K. Langat, Eduard Mas-Claret, Dane Lyddiard, Julian Klepp, Sarah V. A.-M. Legendre, Ben W. Greatrex, Graham L. Jones, and et al. 2021. "The Diversity of Volatile Compounds in Australia’s Semi-Desert Genus Eremophila (Scrophulariaceae)" Plants 10, no. 4: 785. https://doi.org/10.3390/plants10040785