
The Tolerance of Eucalyptus globulus to Soil Contamination with Arsenic

 , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Arsenic Accumulation
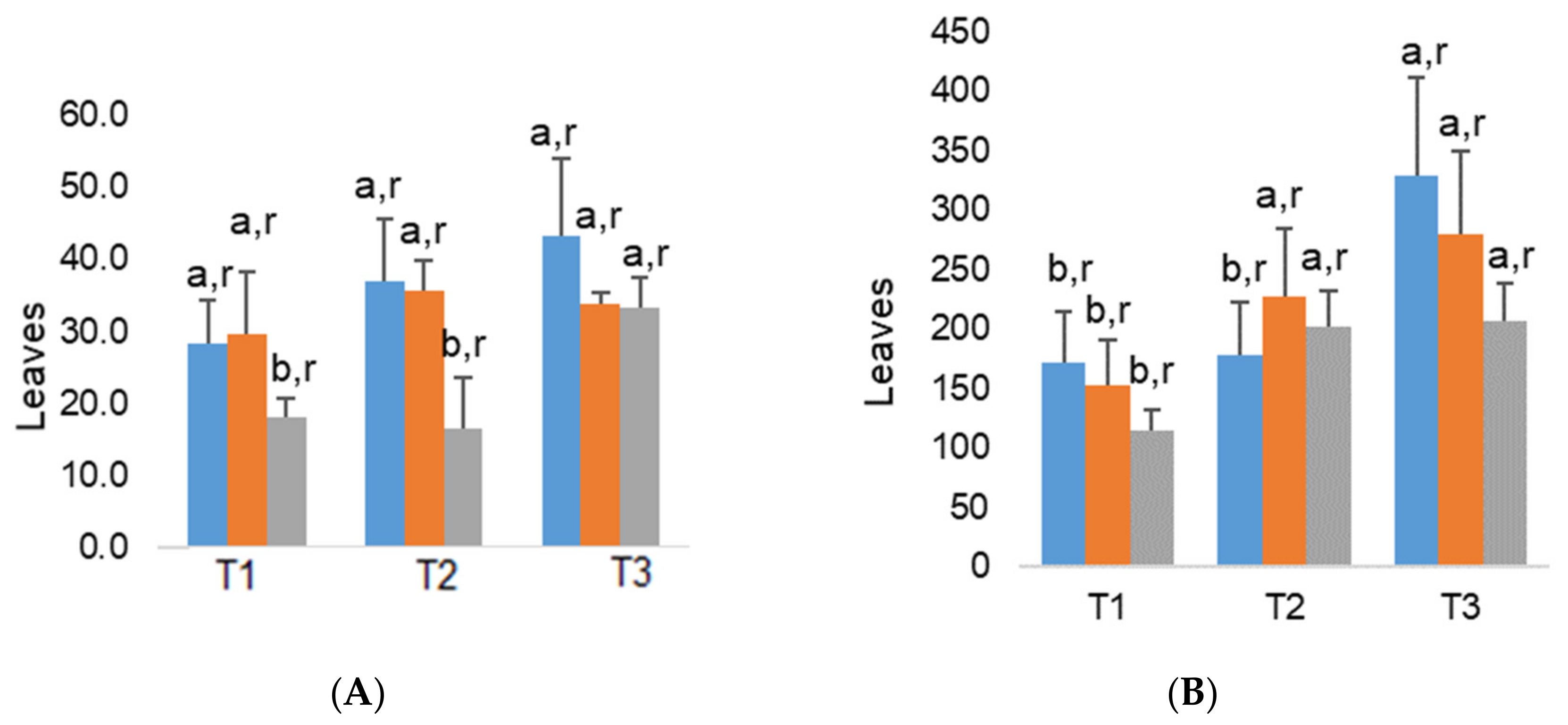
2.2. Plant Growth and Foliar Traits
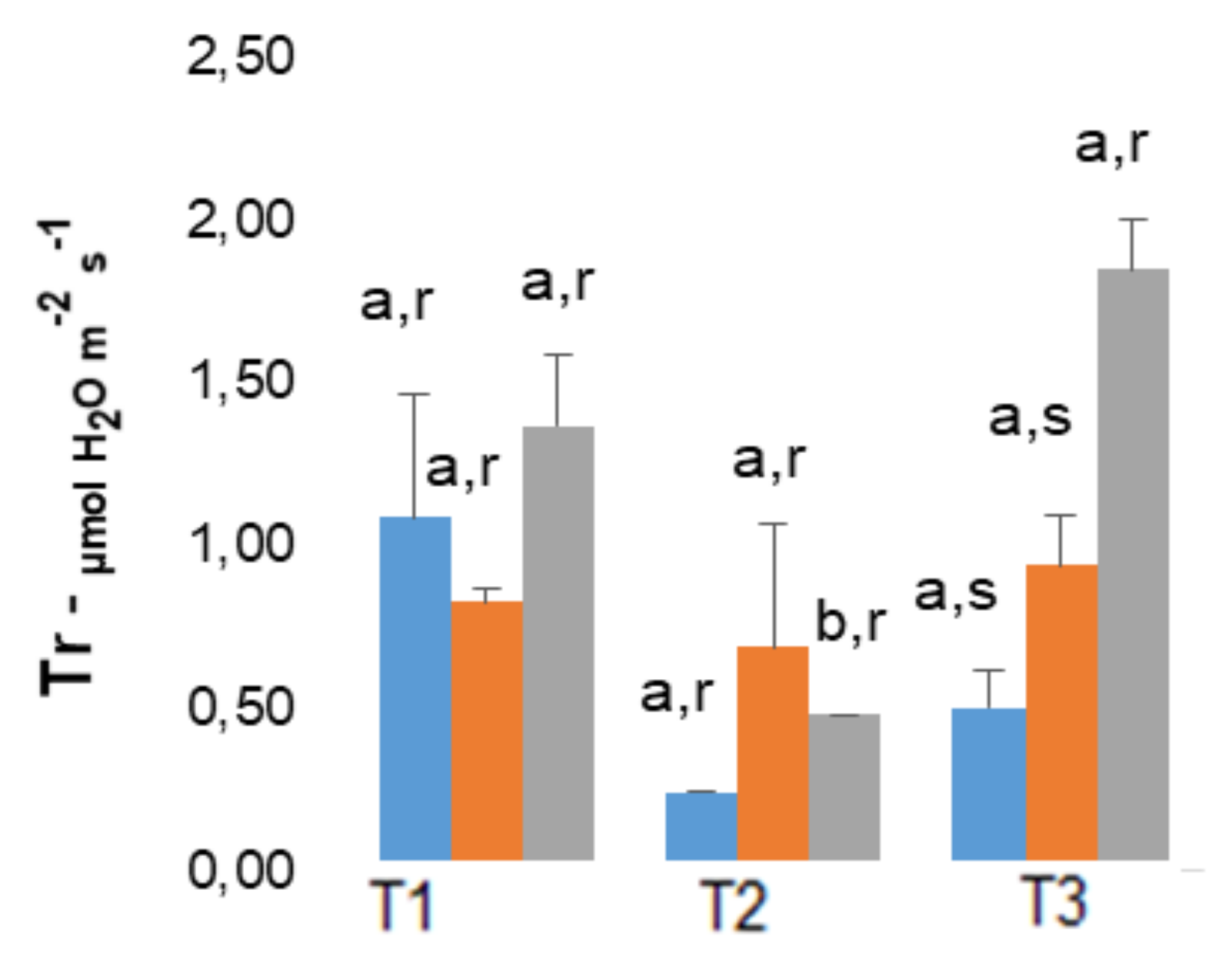
2.3. Photosynthetic Related Parameters
2.4. Minerals Accumulation
2.4.1. Potassium
2.4.2. Calcium
2.4.3. Zinc
2.4.4. Iron
3. Discussion
3.1. As Accumulation, Photosynthesis, and Biomass Production
3.2. Accumulation of Macro- (Ca and K) and Micronutrients (Fe and Zn) in As-Treated Plants
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Growth Analysis
4.3. Leaf Gas Exchanges Analysis
4.4. Chlorophyll a Fluorescence Parameters
4.5. Metal Determination in Soil and Plant Samples
4.6. Micro-Energy Dispersive X-ray Fluorescence (µ-EDXRF)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, C.; Jiang, L.Q.; Zhang, W.J. A review on heavy metal contamination in the soil worldwide: Situation, impact and remediation techniques. Environ Skep Crit. 2014, 3, 24–38. [Google Scholar]
- Masindi, V.; Muedi, K.L. Environmental Contamination by Heavy Metals. In Heavy Metals; IntechOpen: London, UK, 2018. [Google Scholar]
- Reboredo, F.H.S.; Ribeiro, C.A.G. Vertical distribution of Al, Cu, Fe and Zn in soil salt marshes of the Sado estu-ary, Portugal. Int. J. Environ. Stud. 1984, 23, 249–253. [Google Scholar] [CrossRef]
- Reboredo, F. Cadmium accumulation by Halimione portulacoides (L.) aellen. A seasonal study. Mar. Environ. Res. 1992, 33, 17–29. [Google Scholar] [CrossRef]
- Jorge, C. Os Solos Contaminados—A Situação em Portugal (Estudo Preliminar). Relatório 73/98 – NP; Ministério do Equipamento, do Planeamento e da Administração do Território: Lisboa, Portugal; Laboratório Nacional de Engenharia Civil: Lisboa, Portugal, 1998. (In Portuguese) [Google Scholar]
- Santos, S.C.P.-L.; Cruz, M.E.; Barroso, A.M.E.; Fonseca, C.P.S.; Guerra, M.; Carvalho, M.L.; Santos, J.P. Elemental characterization of plants and soils in Panasqueira tungsten mining region. J. Soils. Sed. 2014, 14, 778–784. [Google Scholar] [CrossRef]
- Ferreira da Silva, E.; Durães, N.; Reis, P.; Patinha, C.; Matos, J.; Costa, M.R. An integrative assessment of envi-ronmental degradation of Caveira abandoned mine área (Southern Portugal). J. Geochem. Explor. 2015, 159, 33–47. [Google Scholar] [CrossRef]
- Batista, M.; Abreu, M.; Locutura, J.; De Oliveira, D.; Matos, J.; Silva, C.; Bel-Lan, A.; Martins, L. Evaluation of trace elements mobility from soils to sediments between the Iberian Pyrite Belt and the Atlantic Ocean. J. Geochem. Explor. 2012, 123, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Pelica, J.; Barbosa, S.; Reboredo, F.; Lidon, F.; Pessoa, M.F.; Calvão, T. The paradigm of high concentration of metals of natural or antrophogenic origin in the soils-the case of Neves-Corvo mining area-South of Portugal. J. Geochem. Explor. 2018, 186, 12–23. [Google Scholar] [CrossRef]
- World Health Organization (WHO). IARC Monographs. Arsenic, Metals, Fibres and Dust. A Review of Human Carcinogens. 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK304375/pdf/Bookshelf_NBK304375.pdf (accessed on 25 February 2021).
- Jayasumana, C.; Fonseka, S.; Fernando, A.; Jayalath, K.; Amarasinghe, M.; Siribaddana, S.; Gunatilake, S.; Paranagama, P. Phosphate fertilizer is a main source of arsenic in areas affected with chronic kidney disease of unknown etiology in Sri Lanka. SpringerPlus 2015, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- European Parliament (EU). Directorate General for Internal Policies, Policy Department A: Economic and Scientific Policy. Scientific Aspects Underlying the Regulatory Framework in the Area of Fertilisers-state of Play and Future Reforms 2017. Available online: https://www.europarl.europa.eu/RegData/etudes/IDAN/2016/595354/IPOL_IDA(2016)595354_EN.pdf (accessed on 12 March 2021).
- Li, N.; Wang, J.; Song, W.-Y. Arsenic Uptake and Translocation in Plants. Plant. Cell Physiol. 2016, 57, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Dietary exposure to inorganic arsenic in the European population. EFSA J. 2014, 12, 68. [CrossRef]
- Signes-Pastor, A.J.; Carey, M.; Meharg, A.A. Inorganic arsenic in rice-based products for infants and young children. Food Chem. 2016, 191, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, W.P. Effects and dose–response relationship of skin cancer and Blackfoot disease with arsenic. Environ. Health Perspect. 1977, 19, 109–119. [Google Scholar] [CrossRef]
- Finnegan, P.M.; Chen, W. Arsenic Toxicity: The Effects on Plant Metabolism. Front. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef] [Green Version]
- Freitas, H.; Prasad, M.; Pratas, J. Plant community tolerant to trace elements growing on the degraded soils of São Domingos mine in the south east of Portugal: Environmental implications. Environ. Int. 2004, 30, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Abreu, M.M.; Santos, E.; Magalhães, M.; Fernandes, E. Trace elements tolerance, accumulation and translocation in Cistus populifolius, Cistus salviifolius and their hybrid growing in polymetallic contaminated mine areas. J. Geochem. Explor. 2012, 123, 52–60. [Google Scholar] [CrossRef]
- Reboredo, F. Interaction between copper and zinc and their uptake by Halimione portulacoides (L.) Aellen. Bull. Environ. Contam. Toxicol. 1994, 52, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Caçador, I.; Duarte, B. Salt marshes: An interesting ecosystem to study phytoremediation. In Handbook of Phytoremediation; Nova Science Publishers Inc.: New York, NY, USA, 2011; p. 15, Chapter 26; ISBN 978-1-61728-753-4. [Google Scholar]
- Reboredo, F. Zinc compartmentation in Halimione portulacoides (L.) Aellen and some effects on leaf ultrastructure. Environ. Sci. Pollut. Res. 2012, 19, 2644–2657. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Válega, M.; Mohmood, I.; Gill, S.S.; Tuteja, N.; Duarte, A.C.; Pereira, E. Salt marsh halophyte services to metal-metalloid remediation: Assessment of the processes and underlying mechanisms. Critical Rev. Environ. Sci. Technol. 2014, 44, 2038–2106. [Google Scholar] [CrossRef]
- Dradrach, A.; Karczewska, A.; Szopka, K.; Lewińska, K. Accumulation of Arsenic by Plants Growing in the Sites Strongly Contaminated by Historical Mining in the Sudetes Region of Poland. Int. J. Environ. Res. Public Heal. 2020, 17, 3342. [Google Scholar] [CrossRef]
- Santos, E.S.; Abreu, M.M.; Nabais, C.; Magalhães, M.C.F. Trace element distribution in soils developed on gos-san mine wastes and Cistus ladanifer L. tolerance and bioaccumulation. J. Geochem. Expl. 2012, 123, 45–51. [Google Scholar] [CrossRef]
- Rahman, S.; Kim, K.-H.; Saha, S.K.; Swaraz, A.; Paul, D.K. Review of remediation techniques for arsenic (As) contamination: A novel approach utilizing bio-organisms. J. Environ. Manag. 2014, 134, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.-L.; Khan, M.A.; McGrath, S.P.; Zhao, F.-J. Phytoremediation of arsenic contaminated paddy soils with Pteris vittata markedly reduces arsenic uptake by rice. Environ. Pollut. 2011, 159, 3739–3743. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Sandhya, S.; Prasad, M.; Freitas, H. Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef]
- Pinto, A.P.; De Varennes, A.; Fonseca, R.; Teixeira, D.M. Phytoremediation of Soils Contaminated with Heavy Metals: Techniques and Strategies. In Phytoremediation: Management of Environmental Contaminants; Ansari, A.A., Gill, S.S., Lanza, G.R., Newman, L., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 133–155. [Google Scholar]
- Mughini, G.; Alianiello, F.; Benedetti, A.; Gras, L.M.; Gras, M.A.; Salvati, L. Clonal variation in growth, arsenic and heavy metal uptakes of hybrid Eucalyptus clones in a Mediterranean environment. Agrofor. Syst. 2013, 87, 755–766. [Google Scholar] [CrossRef]
- Luo, J.; Qi, S.; Peng, L.; Wang, J. Phytoremediation efficiency OF CD byEucalyptus globulustransplanted from polluted and unpolluted sites. Int. J. Phytoremediation 2016, 18, 308–314. [Google Scholar] [CrossRef]
- Reboredo, F. Forest Context and Policies in Portugal. Present and Future Challenges; Springer-Verlag: Cham, Switzerland, 2014; p. 239. ISBN 978-3-319-08454-1. [Google Scholar]
- Reboredo, F.; Pais, J. Evolution of forest cover in Portugal: A review of the 12th–20th centuries. J. For. Res. 2014, 25, 249–256. [Google Scholar] [CrossRef]
- Tomé, M. Wood and non-wood production from plantation forests. In Ecological and So-cio-Economic Impacts of Close-to-Nature Forestry and Plantation Forestry: A Comparative Analysis; European Forest Institute: Lisbon, Portugal, 2000; pp. 37–55. [Google Scholar]
- Garg, N.; Singla, P. Arsenic toxicity in crop plants: Physiological effects and tolerance mechanisms. Environ. Chem. Lett. 2011, 9, 303–321. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, J.C.; Rodrigues, A.P.; Lidon, F.C.; Marques, L.M.C.; Leitão, A.E.; Fortunato, A.S.; Pais, I.P.; Silva, M.J.; Scotti-Campos, P.; Lopes, A.; et al. Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffea spp. PLoS ONE 2018, 13, e0198694. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.D.; Tomaz, M.A.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C. Combined effects of elevated [CO2] and high temperature on leaf mineral balance in Coffea spp. plants. Clim. Chang. 2014, 126, 365–379. [Google Scholar] [CrossRef]
- Urbina, I.; Sardans, J.; Beierkuhnlein, C.; Jentsch, A.; Backhaus, S.; Grant, K.; Kreyling, J.; Peñuelas, J. Shifts in the elemental composition of plants during a very severe drought. Environ. Exp. Bot. 2015, 111, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madejón, P.; Marañón, T.; Navarro-Fernández, C.M.; Domínguez, M.T.; Alegre, J.M.; Robinson, B.; Murillo, J.M. Potential of Eucalyptus camaldulensis for phytostabilization and biomonitoring of trace-element contaminated soils. PLoS ONE 2017, 12, e0180240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Meng, M.; Li, L. Arsenic detoxification in Eucalyptus: Subcellular distribution, chemical forms, and sulfhydryl substances. Environ. Sci. Pollut. Res. 2019, 26, 24372–24379. [Google Scholar] [CrossRef] [PubMed]
- King, D.J.; Doronila, A.I.; Feenstra, C.; Baker, A.J.; Woodrow, I.E. Phytostabilisation of arsenical gold mine tailings using four Eucalyptus species: Growth, arsenic uptake and availability after five years. Sci. Total Environ. 2008, 406, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Busch, F.; Hunter, N.P.A.; Ensminger, I. Biochemical constrains limit the potential of the photochemical reflectance index as a predictor of effective quantum efficiency of photosynthesis during the winter-spring transition in Jack pine seedlings. Funct Plant Biol. 2009, 36, 1016–1026. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New flux parameters for the determination of QA redox state and excitation fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Krause, G.H.; Jahns, P. Non-photochemical Energy Dissipation Determined by Chlorophyll Fluorescence Quenching: Characterization and Function. In Plant Cell Monographs; Springer Verlag: Heidelberg, The Netherlands, 2007; pp. 463–495. [Google Scholar]
- Werner, C.; Correia, O.; Beyschlag, W. Characteristic patterns of chronic and dynamic photoinhibition of different functional groups in a Mediterranean ecosystem. Funct. Plant Biol. 2002, 29, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Gusman, G.S.; Oliveira, J.A.; Farnese, F.S.; Cambraia, J. Arsenate and arsenite: The toxic effects on photosynthesis and growth of lettuce plants. ACTA Physiol. Plant. 2013, 35, 1201–1209. [Google Scholar] [CrossRef]
- Karimi, N.; Shayesteh, L.S.; Ghasmpour, H.; Alavi, M. Effects of Arsenic on Growth, Photosynthetic Activity, and Accumulation in Two New Hyperaccumulating Populations of Isatis cappadocica Desv. J. Plant. Growth Regul. 2013, 32, 823–830. [Google Scholar] [CrossRef]
- Farnese, F.S.; Oliveira, J.A.; Farnese, M.S.; Gusman, G.S.; Silveira, N.M.; Siman, I.L. Uptake arsenic by plants: Effects on mineral nutrition, growth and antioxidant capacity. Idesia (Arica) 2014, 32, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Sghaier, D.B.; Duarte, B.; Bankaji, I.; Caçador, I.; Sleimi, N. Growth, chlorophyll fluorescence and mineral nutrition in the halophyte Tamarix gallica cultivated in combined stress conditions: Arsenic and NaCl. J. Photochem. Photobiol. B Biol. 2015, 149, 204–214. [Google Scholar] [CrossRef]
- Eichhorn, S.E.; Evert, R. Raven Biology of Plants, 8th ed.; W.H. Freeman and Company Publishers: New York, NY, USA, 2013. [Google Scholar]
- Thor, K. Calcium—Nutrient and Messenger. Front. Plant. Sci. 2019, 10. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–189. [Google Scholar]
- Mohamed, H.I.; Latif, H.H.; Hanafy, R.S. Influence of Nitric Oxide Application on Some Biochemical Aspects, Endogenous Hormones, Minerals and Phenolic Compounds of Vicia faba Plant Grown under Arsenic Stress. Gesunde Pflanz. 2016, 68, 99–107. [Google Scholar] [CrossRef]
- Shaibur, M.R.; Adjadeh, T.A.; Kawai, S. Effect of phosphorus on the concentrations of arsenic, iron and some other elements in barley grown hydroponically. J. Soil Sci. Plant. Nutr. 2013, 13, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Gunes, A.; Inal, A.; Bagci, E.G.; Kadioglu, Y.K. Combined Effect of Arsenic and Phosphorus on Mineral Element Concentrations of Sunflower. Commun. Soil Sci. Plant. Anal. 2010, 41, 361–372. [Google Scholar] [CrossRef]
- Foelkel, C. Minerais e nutrientes das árvores dos eucaliptos: Aspectos ambientais, fisiológicos, silviculturais e industriais acerca dos elementos inorgânicos presentes nas árvores. Eucalyptus Online Book & Newsletter. 2014, p. 133. Available online: https://www.eucalyptus.com.br/capitulos/capitulo_minerais.pdf (accessed on 25 March 2021).
- Merino, A.; Rodriguez-Lapez, A.; Braes, J.; Rodriguez-Soalleiro, R. Nutrition and growth in newly established plantations of Eucalyptus globulus in northwestern Spain. Annals Forest Sci. 2003, 60, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Carbonell-Barrachina, A.A.; Burló-Carbonell, F.; Mataix-Beneyto, J. Effect of sodium arsenite and sodium chloride on bean plant nutrition (macronutrients). J. Plant Nutr. 1997, 20, 1617–1633. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, V.P.; Tripathi, D.K.; Prasad, S.M.; Chauhan, D.K. Effect of Arsenic on Growth, Arsenic Uptake, Distribution of Nutrient Elements and Thiols in Seedlings ofWrightia arborea (Dennst.) Mabb. Int. J. Phytoremediation 2015, 17, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Ma, L.Q. Effects of arsenic on concentration and distribution of nutrients in the fronds of the arsenic hyperaccumulator Pteris vittata L. Environ. Pollut. 2005, 135, 333–340. [Google Scholar] [CrossRef]
- Rebelo, M.C.; Nunes, M.A.; Ramalho, J.C.; Santos, M.E.; Antunes, M.L. Effects of calcium deficiency on Coffea arabica. Nutrient changes and correlation of calcium levels with some photosynthetic parameters. Plant. Soil 1995, 172, 87–96. [Google Scholar] [CrossRef]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Calcium-dependent regulation of photosynthesis. Biochim. et Biophys. ACTA (BBA)-Bioenerg. 2015, 1847, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Carbonell, A.A.; Aarabi, M.A.; DeLaune, R.D.; Gambrell, R.P.; Patrick, W.H. Arsenic in wetland vegetation: Availability, phytotoxicity, uptake and effects on plant growth and nutrition. Sci. Total Environ. 1998, 217, 189–199. [Google Scholar] [CrossRef]
- Reboredo, F. Cu and Zn uptake by Halimione portulacoides (L.) aellen. A long-term accumulation experiment. Bull. Environ. Contam. Toxicol. 1991, 46, 442–449. [Google Scholar] [CrossRef] [PubMed]
- IPMA. Instituto Português do Mar e da Atmosfera (IPMA). 2016. Available online: http://www.ipma.pt/pt/index.html (accessed on 25 February 2021).
- Lambers, H.; Poorter, H. Inherent Variation in Growth Rate Between Higher Plants: A Search for Physiological Causes and Ecological Consequences. Adv. Ecol. Res. 2004, 34, 283–362. [Google Scholar] [CrossRef]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitao, A.E.; Colwell, F.; Goulão, L.; et al. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Glob. Chang. Biol. 2015, 22, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Stirbet, A. Govindjee On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, S.-B.; Cao, K.-F. Cyclic Electron Flow Plays an Important Role in Photoprotection of Tropical Trees Illuminated at Temporal Chilling Temperature. Plant. Cell Physiol. 2010, 52, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.Q.; Rodrigues, W.P.; Fortunato, A.S.; Leitão, A.E.; Rodrigues, A.P.; Pais, I.P.; Martins, L.D.; Silva, M.J.; Reboredo, F.H.; Partelli, F.L.; et al. Protective Response Mechanisms to Heat Stress in Interaction with High [CO2] Conditions in Coffea spp. Front. Plant. Sci. 2016, 7, 947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EPA. Field Portable X-Ray Fluorescence Spectrometry for the Determination of Elemental Concentration in Soil and Sediment; U.S. Environmental Protection Agency (EPA): Washington DC, WA, USA, 1988.
- Pessanha, S.; Guilherme, A.; Carvalho, M.L. Comparison of matrix effects on portable and stationary XRF spectrometers for cultural heritage samples. Appl Phys. A 2009, 97, 497–505. [Google Scholar] [CrossRef]
- Gallardo, H.; Queralt, I.; Tapias, J.; Guerra, M.; Carvalho, M.L.; Margúi, E. Possibilities of low-power X-ray fluorescence spectrometry methods for rapid multielemental analysis and imaging of vegetal foodstuffs. J. Food Compos. Anal. 2016, 50, 1–9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| March- T1 | Control | 100 μg As mL−1 | 200 μg As mL−1 | |
| As concentration Leaf | BDL | 3.8 ± 0.3 a | 3.6 ± 0.6 a | |
| Stem | BDL | BDL | BDL | |
| Root | BDL | 49.2 ± 5.9 a,s | 74.3 ± 16.9 b,r | |
| Total As (mg plant−1) | – | 2.38 | 2.52 | |
| Total biomass | 92.7 | 148 | 86.2 | |
| Leaf | 37.8 ± 17.3 ab,r | 50.7 ± 19.0 a,r | 23.0 ± 6.1 b,s | |
| Stem | 39.5 ± 15.6 a,r | 50.3 ± 17.0 a,r | 33.3 ± 12.7 a,r | |
| Root | 15.4 ± 1.9 a,t | 47.4 ± 7.8 a,s | 30.9 ± 16.3 ab,s | |
| Plant height | 113 ± 4 a,r | 121 ± 3 a,r | 114 ± 10 a,r | |
| May—T2 | As concentration Leaf | BDL | 5.7 ± 0.5 b | 10.8 ± 0.4 a |
| Stem | BDL | BDL | BDL | |
| Root | BDL | 52.9 ± 11.9 a,s | 102 ± 27.1 a,r | |
| Total As (mg plant−1) | – | 6.48 | 9.84 | |
| Total biomass | 167 | 202 | 194 | |
| Leaf | 32.0 ± 10.0 b,r | 42.8 ± 21.3 ab,r | 52.4± 6.0 a,r | |
| Stem | 41.4 ± 18.2 a,r | 41.1 ± 22.0 a,r | 51.0 ± 19.7 a,r | |
| Root | 93.6 ± 23.8 a,s | 118 ± 76.2 a,r | 91.0 ± 31.0 a,r | |
| Plant height | 133 ± 4 a,r | 120 ± 8 a,r | 134 ± 3 a,r | |
| July—T3 | As concentration Leaf | BDL | 6.7 ± 1.6 a | 4.4 ± 0.5 a |
| Stem | BDL | BDL | BDL | |
| Root | BDL | 54.3 ± 28.7 a,s | 82.3 ± 22.1 ab,r | |
| Total As (mg plant−1) | – | 8.19 | 8.91 | |
| Total biomass | 317 | 235 | 196 | |
| Leaf | 40.2 ± 8.4 a,r | 39.6 ± 15.0 a,r | 42.9 ± 18.8 a,rs | |
| Stem | 66.1 ± 13.8 a,r | 49.8 ± 23.6 a,r | 46.8 ± 24.0 a,r | |
| Root | 211 ± 36 a,r | 146 ± 59 ab,r | 106 ± 47.6 b,r | |
| Plant height | 136 ± 8 a,r | 133 ± 7 a,r | 138 ± 5 a,r |
| Treatment | March (T1) | May (T2) | July (T3) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Fo | |||||||||
| Control | 0.13 | ± | 0.01 b,r | 0.21 | ± | 0.01 a,r | 0.23 | ± | 0.03 a,r |
| 100 As | 0.13 | ± | 0.01 b,r | 0.19 | ± | 0.03 a,r | 0.23 | ± | 0.02 a,r |
| 200 As | 0.14 | ± | 0.00 b,r | 0.12 | ± | 0.01 b,s | 0.23 | ± | 0.04 a,r |
| Fv/Fm | cv | ||||||||
| Control | 0.77 | ± | 0.01 a,r | 0.77 | ± | 0.04 a,r | 0.76 | ± | 0.03 a,r |
| 100 As | 0.78 | ± | 0.03 a,r | 0.77 | ± | 0.02 a,r | 0.76 | ± | 0.02 a,r |
| 200 As | 0.74 | ± | 0.04 a,r | 0.77 | ± | 0.02 a,r | 0.76 | ± | 0.04 a,r |
| Y(II) | |||||||||
| Control | 0.13 | ± | 0.04 a,r | 0.17 | ± | 0.02 a,rs | 0.17 | ± | 0.01 a,r |
| 100 As | 0.10 | ± | 0.04 b,r | 0.24 | ± | 0.03 a,r | 0.14 | ± | 0.03 ab,r |
| 200 As | 0.17 | ± | 0.06 a,r | 0.15 | ± | 0.02 a,s | 0.12 | ± | 0.01 a,s |
| Y(NPQ) | |||||||||
| Control | 0.55 | ± | 0.01 a,r | 0.71 | ± | 0.02 a,r | 0.66 | ± | 0.01 a,rs |
| 100 As | 0.52 | ± | 0.12 a,r | 0.59 | ± | 0.05 a,s | 0.65 | ± | 0.02 a,s |
| 200 As | 0.50 | ± | 0.02 b,r | 0.64 | ± | 0.04 b,s | 0.71 | ± | 0.03 a,r |
| Y(NO) | |||||||||
| Control | 0.32 | ± | 0.03 a,r | 0.12 | ± | 0.01 b,s | 0.17 | ± | 0.02 b,r |
| 100 As | 0.38 | ± | 0.11 a,r | 0.17 | ± | 0.03 b,rs | 0.21 | ± | 0.01 b,r |
| 200 As | 0.34 | ± | 0.08 a,r | 0.22 | ± | 0.03 b,r | 0.18 | ± | 0.03 b,r |
| qN | |||||||||
| Control | 0.85 | ± | 0.02 b,r | 0.94 | ± | 0.01 a,r | 0.92 | ± | 0.01 a,r |
| 100 As | 0.82 | ± | 0.09 a,r | 0.88 | ± | 0.04 a,r | 0.91 | ± | 0.02 a,r |
| 200 As | 0.83 | ± | 0.03 b,r | 0.89 | ± | 0.03 ab,r | 0.92 | ± | 0.01 a,r |
| qL | |||||||||
| Control | 0.33 | ± | 0.07 a,r | 0.44 | ± | 0.06 a,r | 0.51 | ± | 0.05 a,r |
| 100 As | 0.23 | ± | 0.04 a,r | 0.56 | ± | 0.18 a,r | 0.44 | ± | 0.07 a,r |
| 200 As | 0.44 | ± | 0.07 a,r | 0.36 | ± | 0.03 a,r | 0.33 | ± | 0.01 a,r |
| Fv’/Fm’ | |||||||||
| Control | 0.32 | ± | 0.03 a,r | 0.33 | ± | 0.09 a,r | 0.28 | ± | 0.04 a,r |
| 100 As | 0.32 | ± | 0.07 a,r | 0.39 | ± | 0.04 a,r | 0.28 | ± | 0.06 a,r |
| 200 As | 0.32 | ± | 0.33 a,r | 0.33 | ± | 0.05 a,r | 0.30 | ± | 0.02 a,r |
| Fs/Fm’ | |||||||||
| Control | 0.87 | ± | 0.04 a,r | 0.83 | ± | 0.02 a,rs | 0.84 | ± | 0.01 a,r |
| 100 As | 0.90 | ± | 0.04 ab,r | 0.76 | ± | 0.03 b,s | 0.86 | ± | 0.03 a,r |
| 200 As | 0.83 | ± | 0.06 a,r | 0.85 | ± | 0.02 a,r | 0.88 | ± | 0.01 a,r |
| PlChr | |||||||||
| Control | 7.12 | ± | 3.29 a,r | 5.54 | ± | 5.50 a,r | 7.28 | ± | 3.09 a,r |
| 100 As | 5.16 | ± | 3.44 a,r | 6.69 | ± | 2.47 a,r | 7.33 | ± | 2.54 a,r |
| 200 As | 10.3 | ± | 5.9 a,r | 7.18 | ± | 2.30 a,r | 7.12 | ± | 5.40 a,r |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reboredo, F.H.; Pelica, J.; Lidon, F.C.; Pessoa, M.F.; Silva, M.M.; Guerra, M.; Leitão, R.; Ramalho, J.C. The Tolerance of Eucalyptus globulus to Soil Contamination with Arsenic. Plants 2021, 10, 627. https://doi.org/10.3390/plants10040627
Reboredo FH, Pelica J, Lidon FC, Pessoa MF, Silva MM, Guerra M, Leitão R, Ramalho JC. The Tolerance of Eucalyptus globulus to Soil Contamination with Arsenic. Plants. 2021; 10(4):627. https://doi.org/10.3390/plants10040627
Chicago/Turabian StyleReboredo, Fernando Henrique, João Pelica, Fernando C. Lidon, Maria F. Pessoa, Maria Manuela Silva, Mauro Guerra, Roberta Leitão, and José C. Ramalho. 2021. "The Tolerance of Eucalyptus globulus to Soil Contamination with Arsenic" Plants 10, no. 4: 627. https://doi.org/10.3390/plants10040627