
Light-Mediated Reduction in Photosynthesis in Closed Greenhouses Can Be Compensated for by CO2 Enrichment in Tomato Production


Abstract
:1. Introduction
2. Results
2.1. Standard Values of Photosynthesis and Transpiration
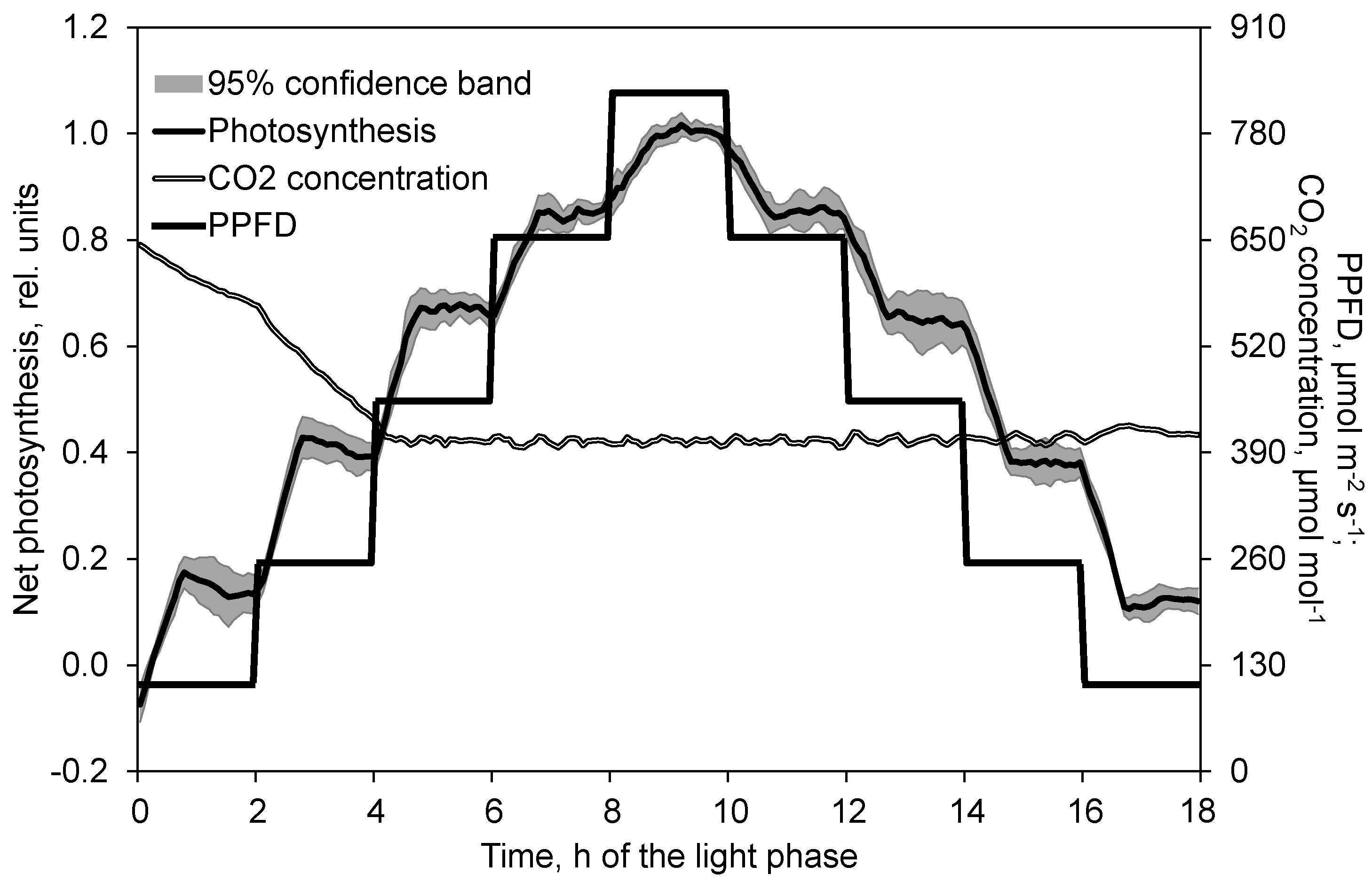
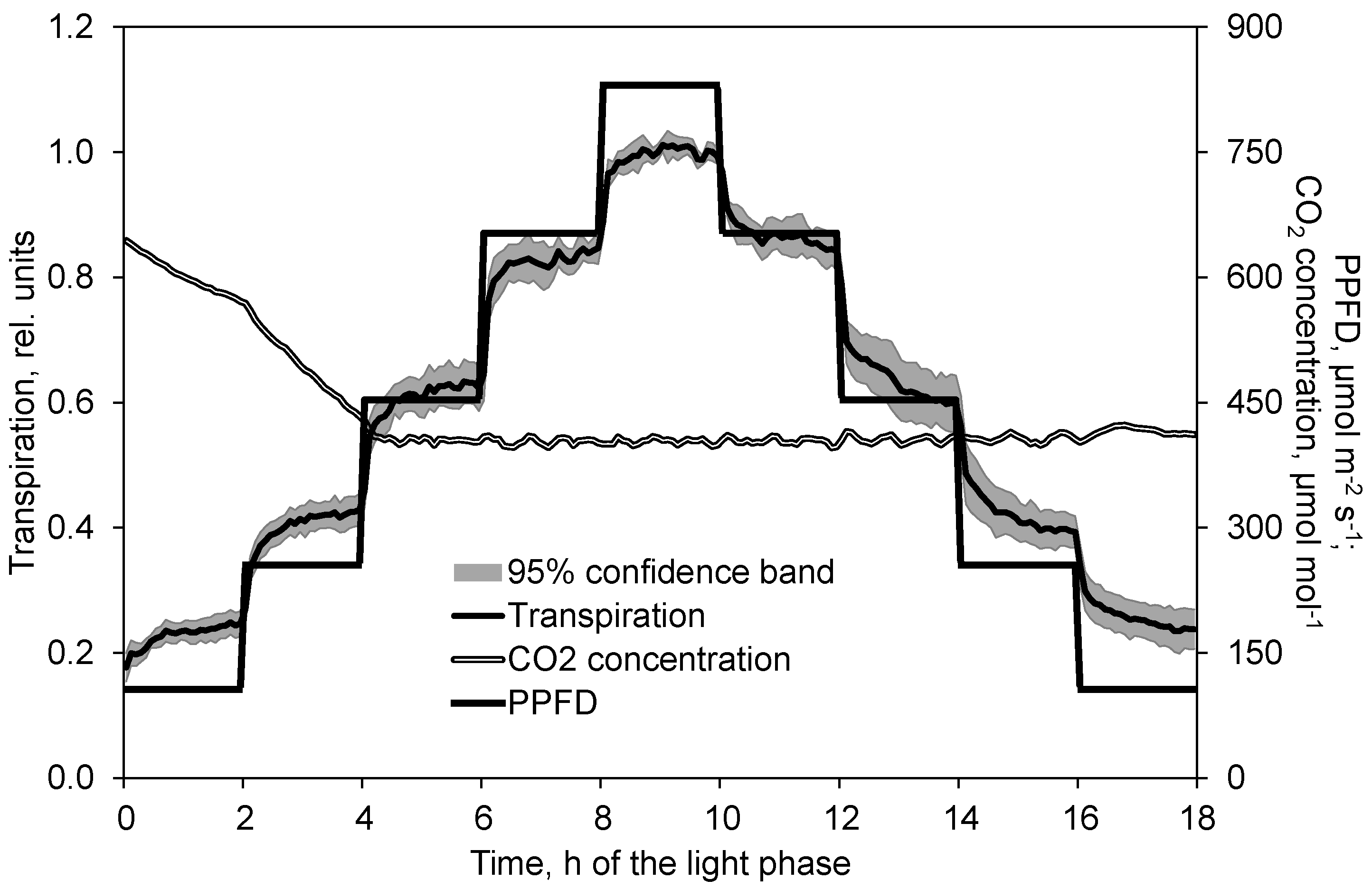
2.2. Course of Photosynthesis and Transpiration Depending on PPFD
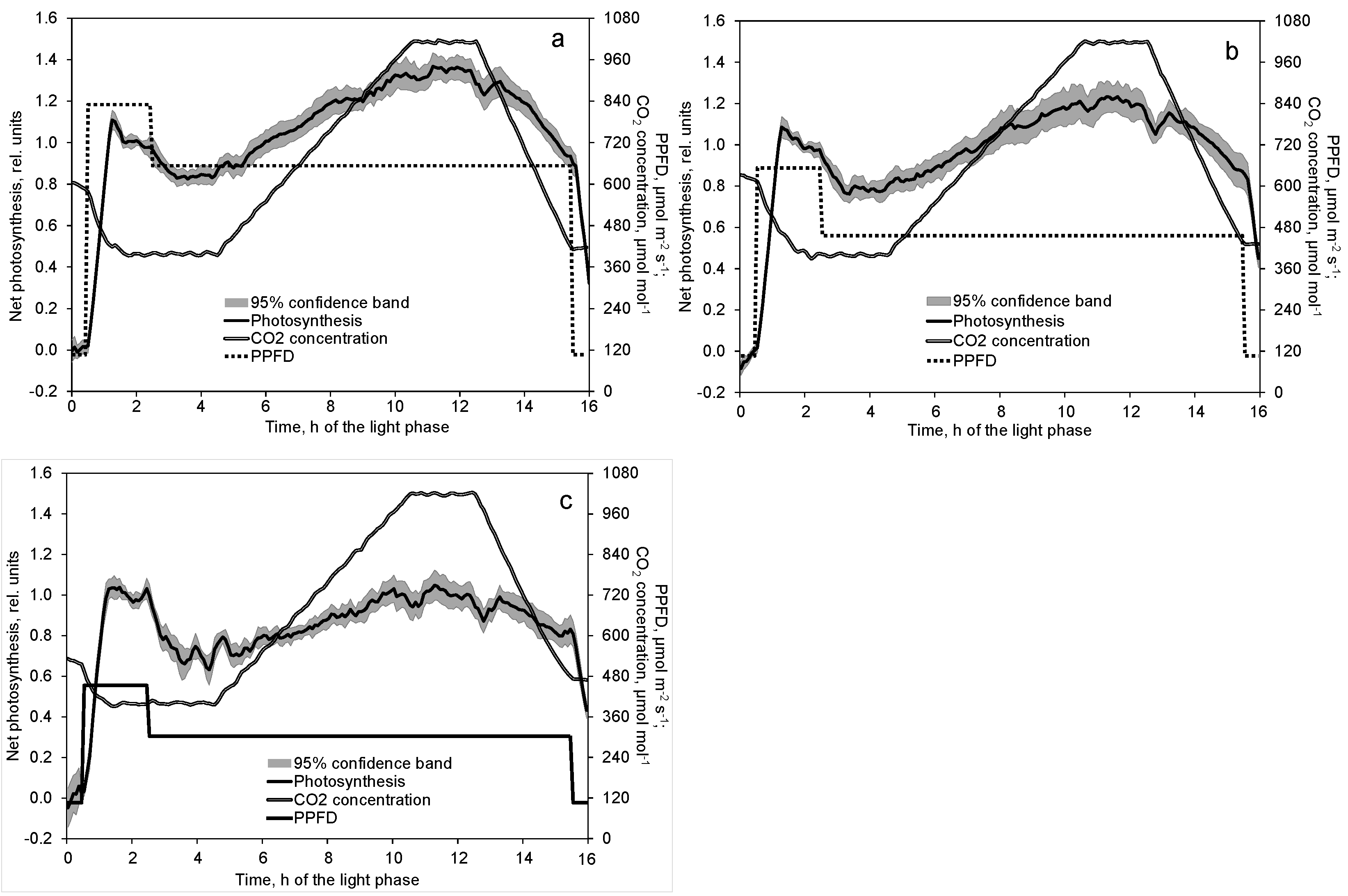
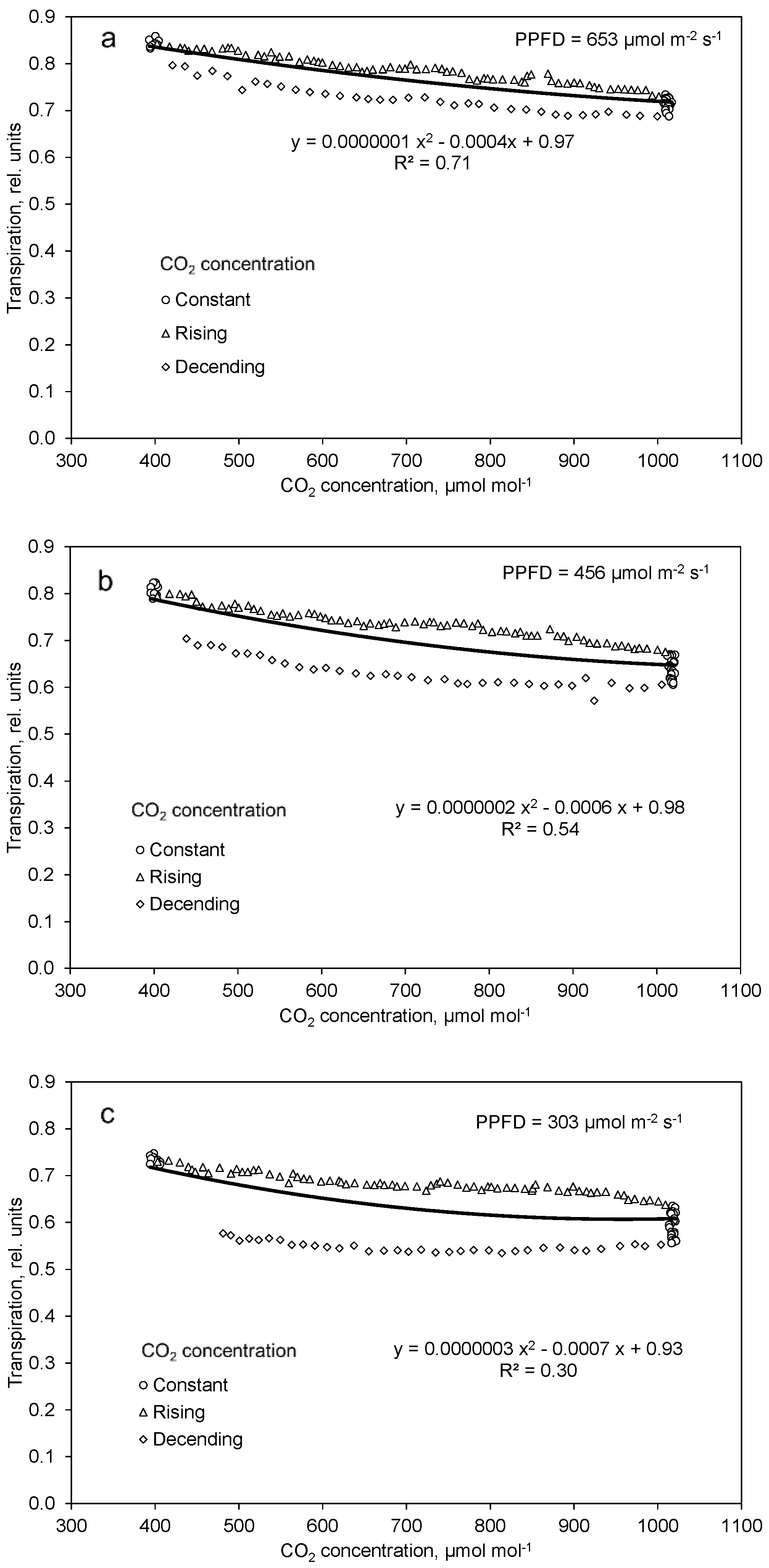
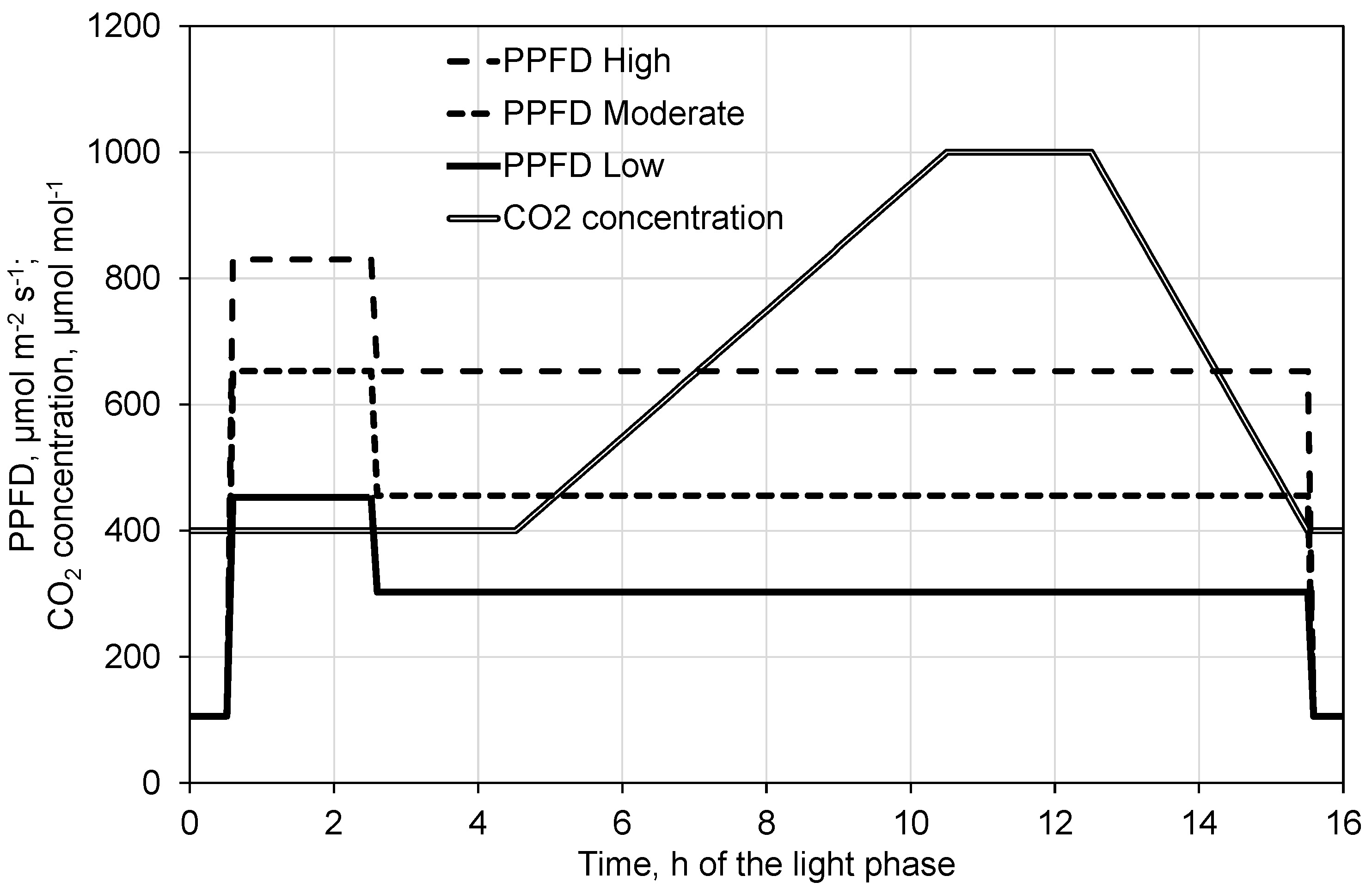
2.3. Effect of Decreased PPFD Followed by Increasing CO2 Concentration on Photosynthesis and Transpiration
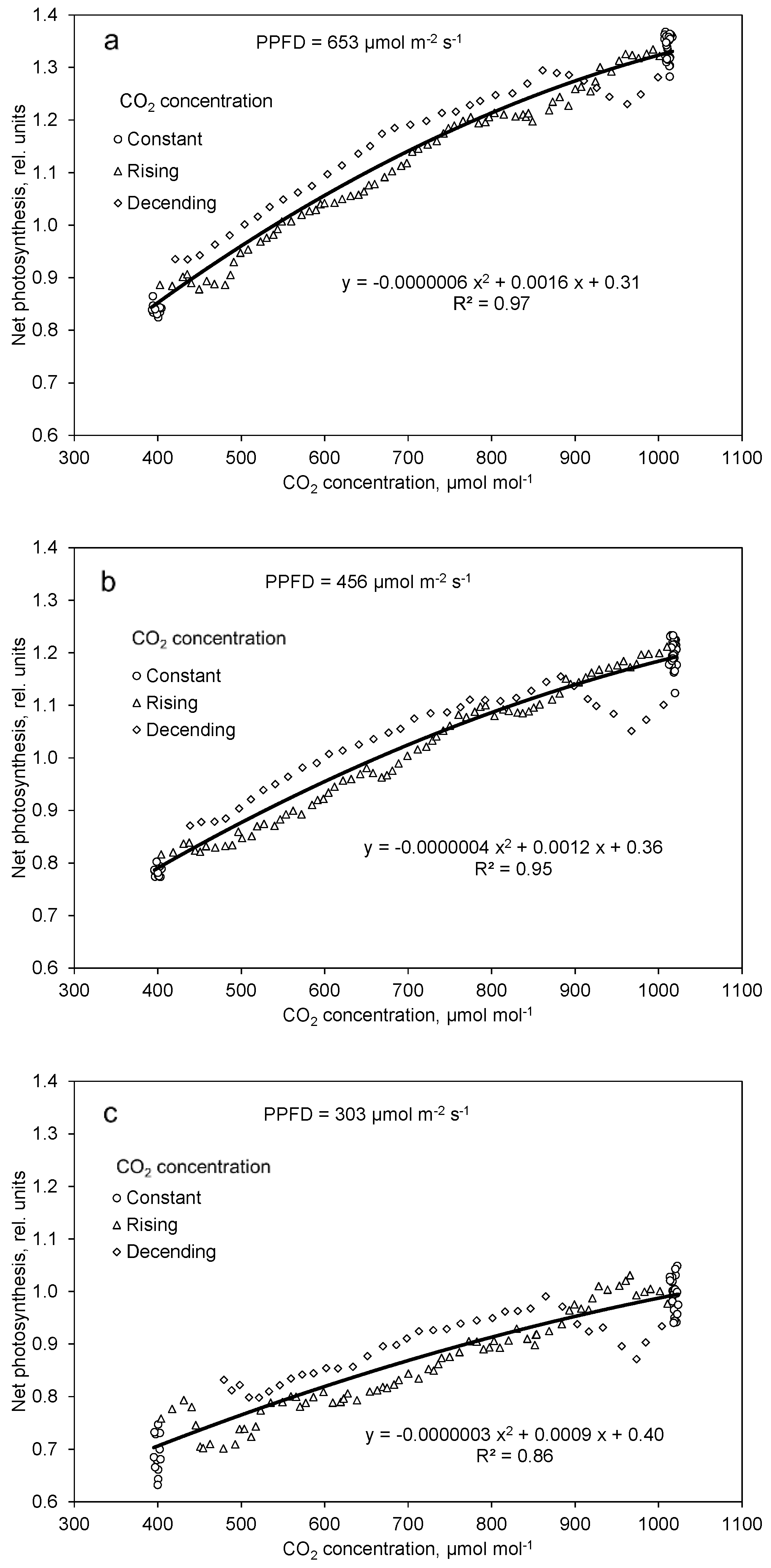
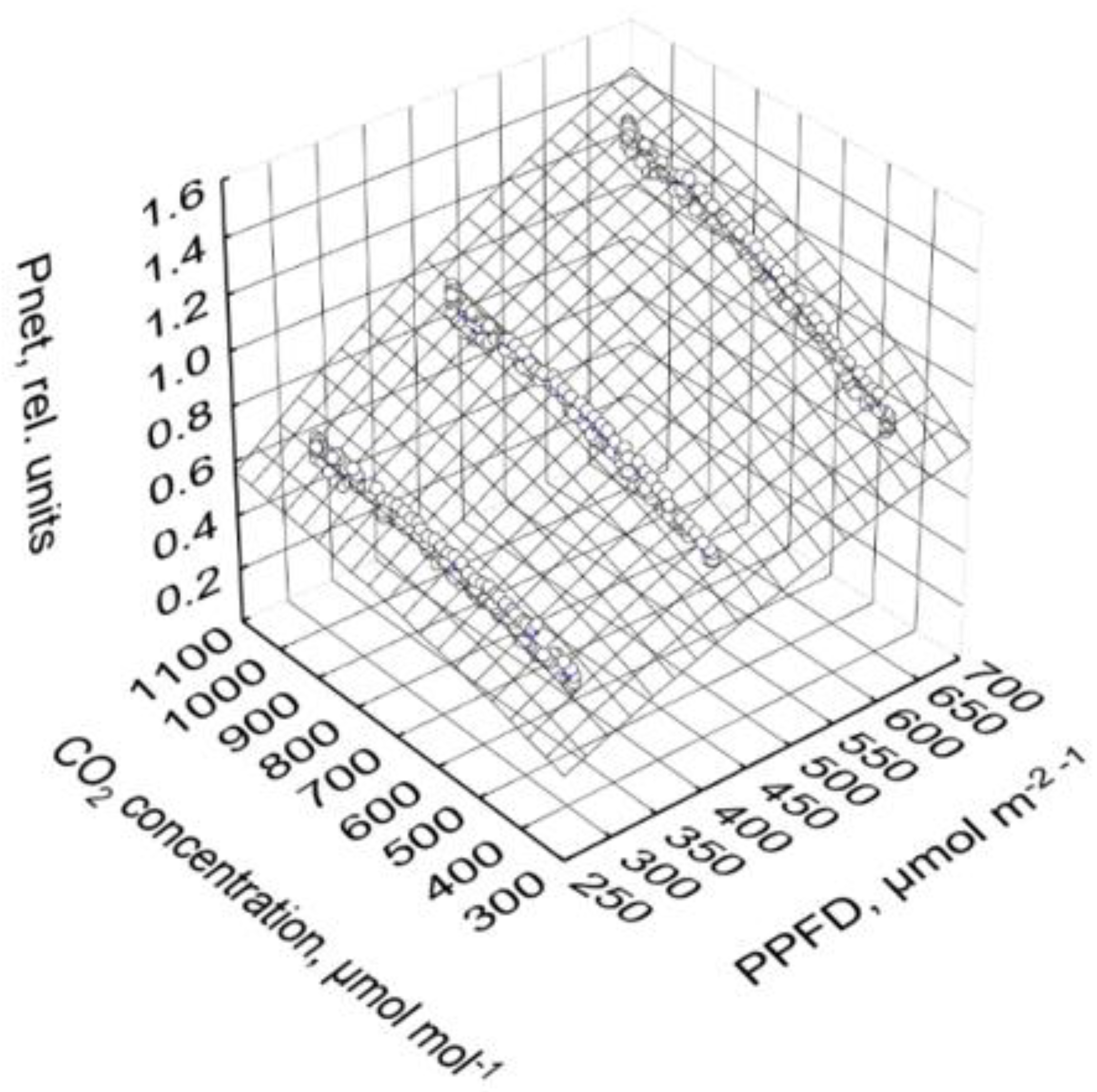
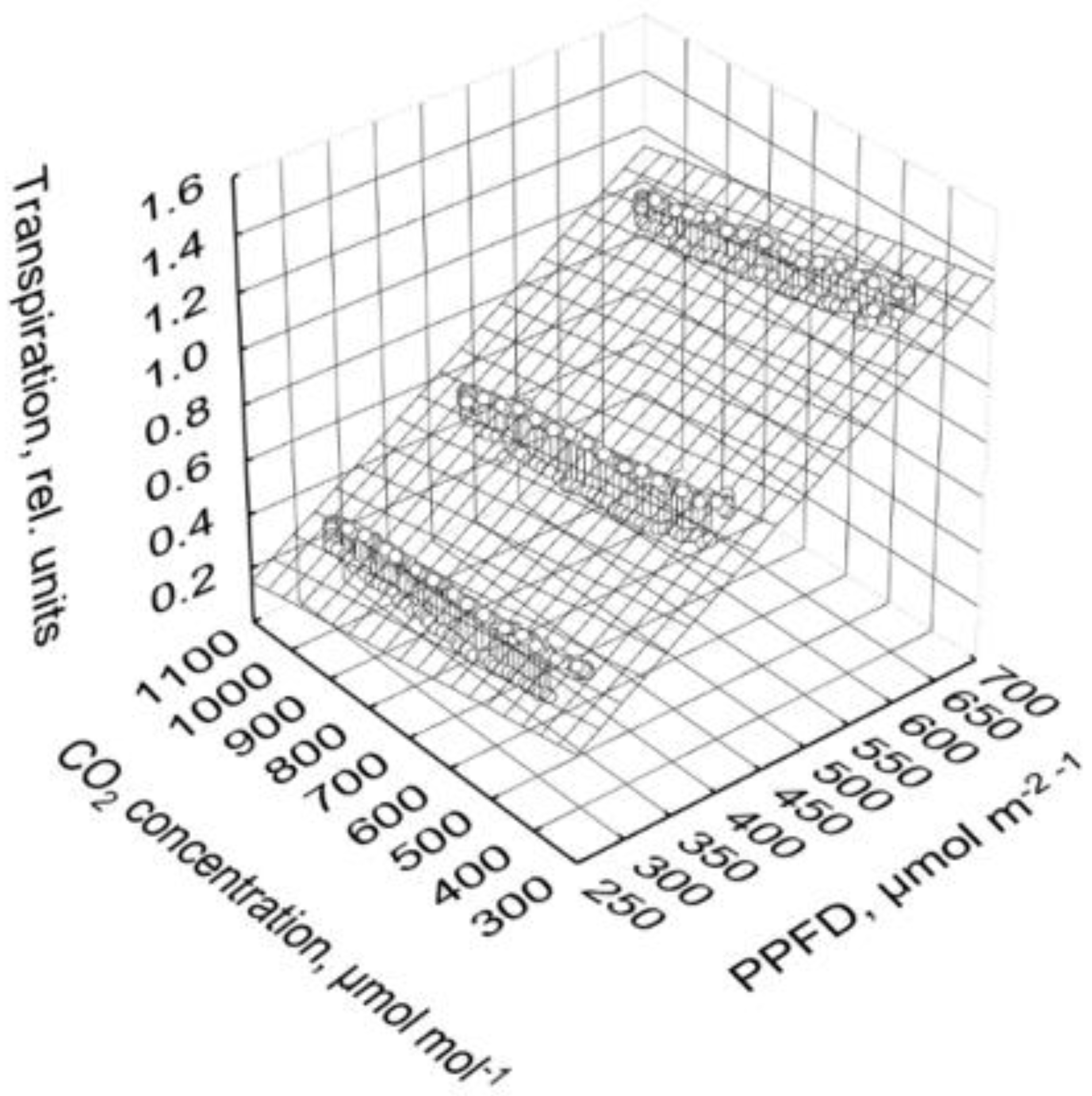
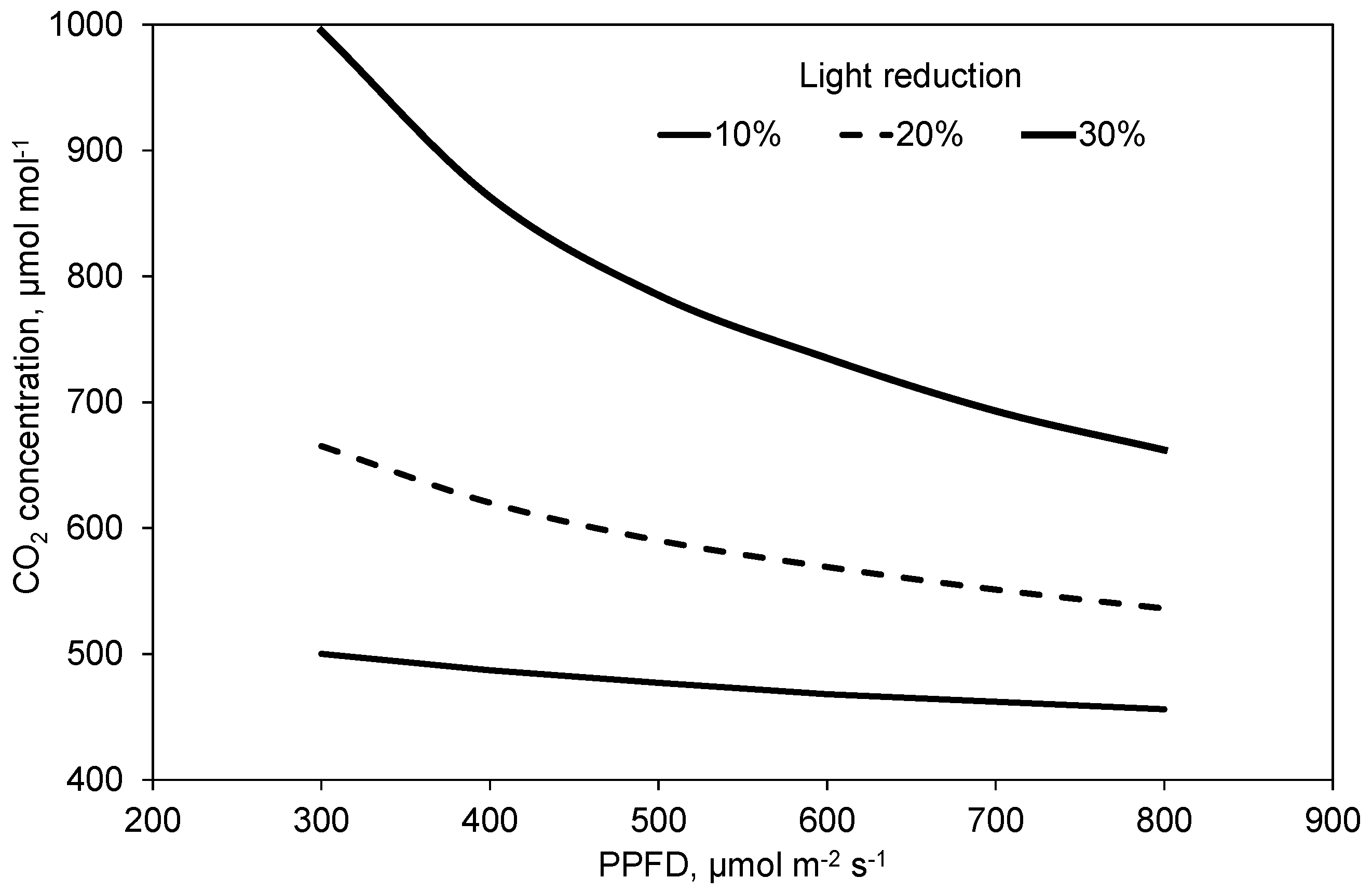
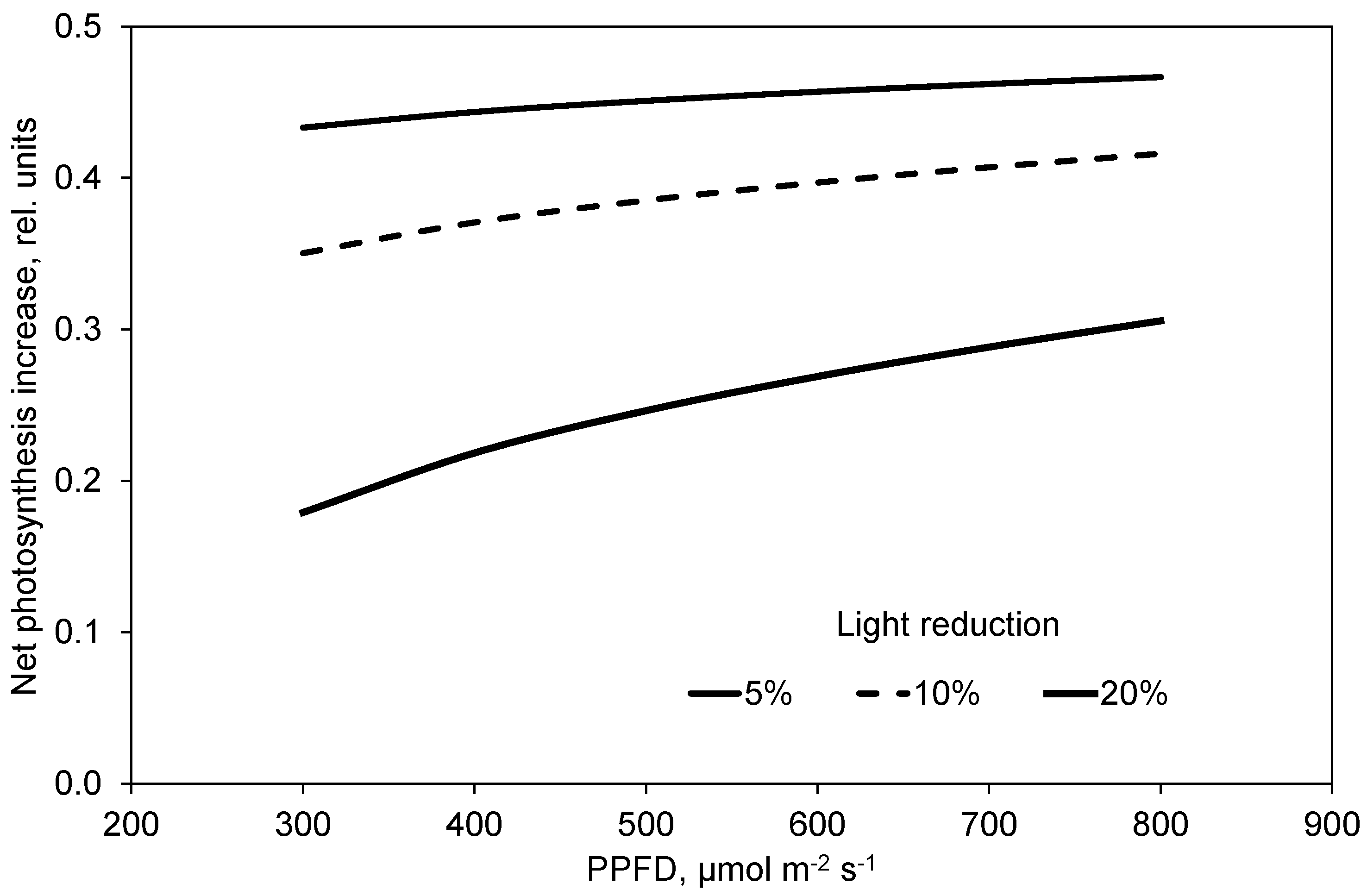
2.4. Photosynthesis and Transpiration Affected by PPFD and CO2 Concentration
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Design
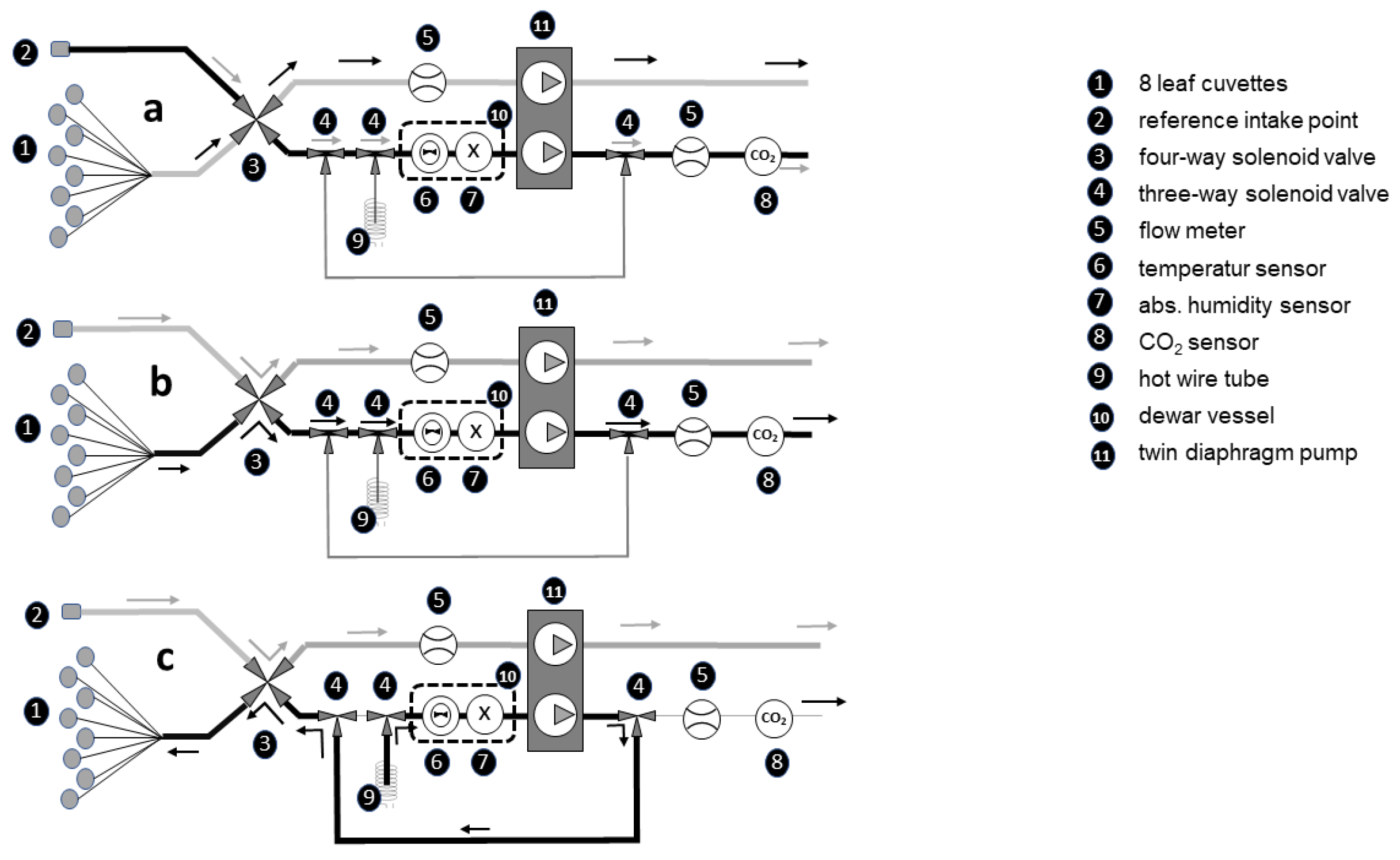
4.3. Measurements of Photosynthesis and Transpiration
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campen, J.B.; Bot, G.P.A. Dehumidification in greenhouses by condensation on finned pipes. Biosyst. Eng. 2002, 82, 177–185. [Google Scholar] [CrossRef]
- Ntinas, G.; Dannehl, D.; Schuch, I.; Rocksch, T.; Schmidt, U. Sustainable greenhouse production with minimised carbon footprint by energy export. Biosyst. Eng. 2020, 189, 164–178. [Google Scholar] [CrossRef]
- Kläring, P.; Krumbein, A. The effect of constraining the intensity of solar radiation on the photosynthesis, growth, yield and product quality of tomato. J. Agro. Crop. Sci. 2013, 199, 351–359. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Broekhuijsen, A.G.M.; Meinen, E.; Nijs, E.M.F.M.; Raaphorst, M.G.M. Quantification of the growth response to light quantity of greenhouse grown crops. Acta Horticulture 2006, 711, 97–103. [Google Scholar] [CrossRef]
- Lim, S.-L.; Voon, C.P.; Guan, X.; Yang, Y.; Gardeström, P.; Lim, B.L. In planta study of photosynthesis and photorespiration using NADPH and NADH/NAD+ fluorescent protein sensors. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cen, Y.-P.; Sage, R.F. The regulation of Rubisco activity in response to variation in temperature and atmospheric CO2 partial pressure in sweet potato. Plant Physiol. 2005, 139, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Dannehl, D.; Suhl, J.; Huyskens-Keil, S.; Ulrichs, C.; Schmidt, U. Effects of a special solar collector greenhouse on water balance, fruit quantity and fruit quality of tomatoes. Agric. Water Manag. 2014, 134, 14–23. [Google Scholar] [CrossRef]
- De Gelder, A.; Dieleman, J.A.; Bot, G.P.A.; Marcelis, L.F.M. An overview of climate and crop yield in closed greenhouses. J. Hort. Sci. Biotech. 2012, 87, 193–202. [Google Scholar] [CrossRef]
- Grisey, A.; Grasselly, D.; Rosso, L.; D’Amaral, F.; Melamedoff, S. Using heat exchangers to cool and heat a closed tomato greenhouse: Application in the south of France. Acta Horiculturae 2009, 893, 405–411. [Google Scholar] [CrossRef]
- Quian, T.; Dieleman, J.A.; Elings, A.; De Gelder, A.; Van Kooten, O.; Marcelis, L.F.M. Comparison of climate and production in closed, semi-closed, and open grennhouses. Acta Horticulture 2011, 893, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Nederhoff, E.M.; Vegter, J.G. Photosynthesis of stands of tomato, cucumber and sweet pepper measured in greenhouses under various CO2-concentrations. Ann. Bot. 1994, 73, 353–361. [Google Scholar] [CrossRef]
- Körner, O.; Heuvelink, E.; Niu, Q. Quantification of temperature, CO2, and light effects on crop photosynthesis as a basis for model-based greenhouse climate control. J. Hort. Sci. Biotech. 2009, 84, 233–239. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Way, D.A. Two decades of sunfleck research: Looking back to move forward. Tree Physiol. 2012, 32, 1059–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, E.; Morales, A.; Harbinson, J.; Kromdijk, J.; Heuvelink, E.; Marcelis, L.F.M. Dynamic photosynthesis in different environmental conditions. J. Exp. Bot. 2015, 66, 2415–2426. [Google Scholar] [CrossRef] [Green Version]
- De Gelder, A.; Heuvelink, E.; Opdam, J.J.G. Tomato yield in a closed greenhouse and comparison with simulated yields in closed and conventional greenhouses. Acta Horticulture 2005, 691, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Bunce, J.A. Carbon dioxide effects on stomatal responses to the environment and water use by crops under field conditions. Oecologia 2004, 140, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shimono, H.; Nakamura, H.; Hasegawa, T.; Okada, M. Lower responsiveness of canopy evapotranspiration rate than of leaf stomatal conductance to open-air CO2 elevation in rice. Glob. Chang. Biol. 2013, 19, 2444–2453. [Google Scholar] [CrossRef] [PubMed]
- Stinziano, J.R.; Morgan, P.M.; Lynch, D.J.; Saathoff, A.J.; McDermitt, D.K.; Hanson, D.T. The rapid A–Ci response: Photosynthesis in the phenomic era. Plant Cell Environ. 2017, 40, 1256–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Boulard, T.; Haxaire, R. Air speed profiles in a naturally ventilated greenhouse with a tomato crop. Agric. For. Meteorol. 1999, 96, 181–188. [Google Scholar] [CrossRef]
- Kläring, H.P.; Körner, O. Design of a real-time gas-exchange measurement system for crop stands in environmental scenarios. Agronomy 2020, 10, 737. [Google Scholar] [CrossRef]
- Nederhoff, E.M.; Rijsdijk, A.A.; De Graaf, R. Leaf conductance and rate of crop transpiration of greenhouse grown sweet pepper (Capsicum annuum L.) as affected by carbon dioxide. Sci. Horticultural 1992, 52, 283–301. [Google Scholar] [CrossRef]
- McAusland, L.; Vialet-Chabrand, S.; Davey, P.; Baker, N.R.; Brendel, O.; Lawson, T. Effects of kinetics of light-induced stomatal responses on photosynthesis and water-use efficiency. New Phytol. 2016, 211, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.D.B.; Press, M.C.; Scholes, J.D.; Watling, J.R. Relative enhancement of photosynthesis and growth at elevated CO2 is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling. Plant Cell Environ. 2002, 25, 1701–1714. [Google Scholar] [CrossRef]
- Tomimatsu, H.; Iio, A.; Adachi, M.; Saw, L.G.; Fletcher, C.D.; Tang, Y. High CO2 concentration increases relative leaf carbon gain under dynamic light in Dipterocarpus sublamellatus seedlings in a tropical rain forest, Malaysia. Tree Physiol. 2014, 34, 944–954. [Google Scholar] [CrossRef] [Green Version]
- Van Iersel, M.W.; Bugbee, B. A multiple chamber, semicontinuous, crop carbon dioxide exchange system: Design, calibration, and data interpretation. J. Am. Soc. Horticultural Sci. 2000, 125, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Körner, O. Evaluation of crop photosynthesis models for dynamic climate control. Acta Horiculturae 2003, 654, 295–302. [Google Scholar] [CrossRef]
- Teitel, M.; Atias, M.; Schwartz, A.; Cohen, S. Use of a greenhouse as an open chamber for canopy gas exchange measurements: Methodology and validation. Agric. For. Meteorol. 2011, 151, 1346–1355. [Google Scholar] [CrossRef]
- Ramirez, T.; Kläring, H.P.; Körner, O. Does interruption of electricity supply for supplementary lighting affect the long-term carbon dioxide exchange of greenhouse tomato crops? Biosystems Eng. 2019, 187, 69–80. [Google Scholar] [CrossRef]
- Thongbai, P.; Kozai, T.; Ohyama, K. CO2 and air circulation effects on photosynthesis and transpiration of tomato seedlings. Sci. Hortic. 2010, 126, 338–344. [Google Scholar] [CrossRef]
- Dannehl, D.; Schuch, I.; Schmidt, U. Plant production in solar collector greenhouses—Influence on yield, energy use efficiency and reduction in CO2 emissions. J. Agric. Sci. 2013, 5, 34–45. [Google Scholar] [CrossRef]
- Heuvelink, E.; Bakker, M.; Marcelis, L.F.M.; Raaphorst, M. Climate and yield in a closed greenhouse. Acta Hort. 2008, 801, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
- De Kreij, C.; Voogt, W.; Van den Bos, A.L.; Baas, R. Nutrient Solutions for the Growth of Tomato in Closed Systems. Proefstation voor Bloemistrij en Glasgroente 1997, 2, 21. (In Dutch) [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cycle/Chamber | Net Photosynthesis, µmol m−2 s−1 | ||
|---|---|---|---|
| At High PPFD 830 µmol m−2 s−1 | At Moderate PPFD 653 µmol m−2 s−1 | At Low PPFD 453 µmol m−2 s−1 | |
| 1/A | 17.09 | 15.89 | 11.63 |
| 1/B | 15.58 | 14.83 | 10.62 |
| 2/A | 15.07 | 12.43 | 10.57 |
| 2/B | 14.43 | 11.26 | 9.01 |
| 3/A | 13.81 | 11.48 | 7.78 |
| 3/B | 11.91 | 9.31 | 7.79 |
| 4/A | 8.88 | 10.58 | 7.55 |
| 4/B | 10.02 | 8.06 | 6.92 |
| 5/A | 13.13 | 11.35 | 8.63 |
| 5/B | 9.63 | 8.34 | 6.87 |
| Cycle/Chamber | Transpiration, mmol m−2 s−1 | ||
|---|---|---|---|
| At High PPFD 830 µmol m−2 s−1 | At Moderate PPFD 653 µmol m−2 s−1 | At Low PPFD 453 µmol m−2 s−1 | |
| 1/A | 4.58 | 3.74 | 3.03 |
| 1/B | 4.33 | 3.55 | 2.95 |
| 2/A | 4.11 | 3.25 | 2.63 |
| 2/B | 4.00 | 3.18 | 2.38 |
| 3/A | 3.81 | 2.74 | 2.15 |
| 3/B | 3.23 | 2.24 | 2.00 |
| 4/A | 2.94 | 2.99 | 2.01 |
| 4/B | 3.26 | 2.59 | 2.02 |
| 5/A | 3.95 | 3.13 | 2.19 |
| 5/B | 3.41 | 2.56 | 2.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dannehl, D.; Kläring, H.-P.; Schmidt, U. Light-Mediated Reduction in Photosynthesis in Closed Greenhouses Can Be Compensated for by CO2 Enrichment in Tomato Production. Plants 2021, 10, 2808. https://doi.org/10.3390/plants10122808
Dannehl D, Kläring H-P, Schmidt U. Light-Mediated Reduction in Photosynthesis in Closed Greenhouses Can Be Compensated for by CO2 Enrichment in Tomato Production. Plants. 2021; 10(12):2808. https://doi.org/10.3390/plants10122808
Chicago/Turabian StyleDannehl, Dennis, Hans-Peter Kläring, and Uwe Schmidt. 2021. "Light-Mediated Reduction in Photosynthesis in Closed Greenhouses Can Be Compensated for by CO2 Enrichment in Tomato Production" Plants 10, no. 12: 2808. https://doi.org/10.3390/plants10122808