
Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat Against Rhizoctonia solani

 ,
, 
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
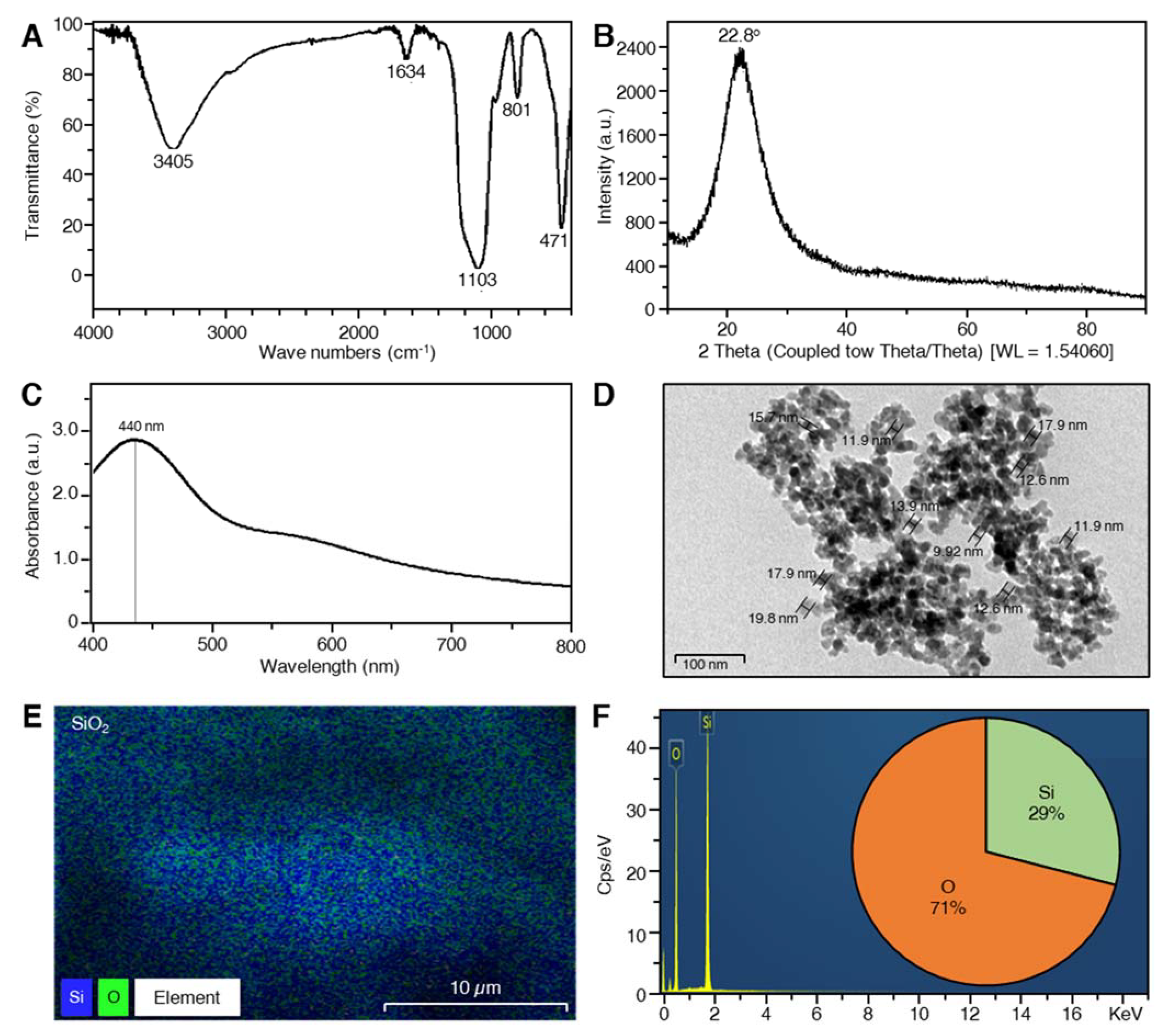
2.1. Characterization of SiO2 NPs
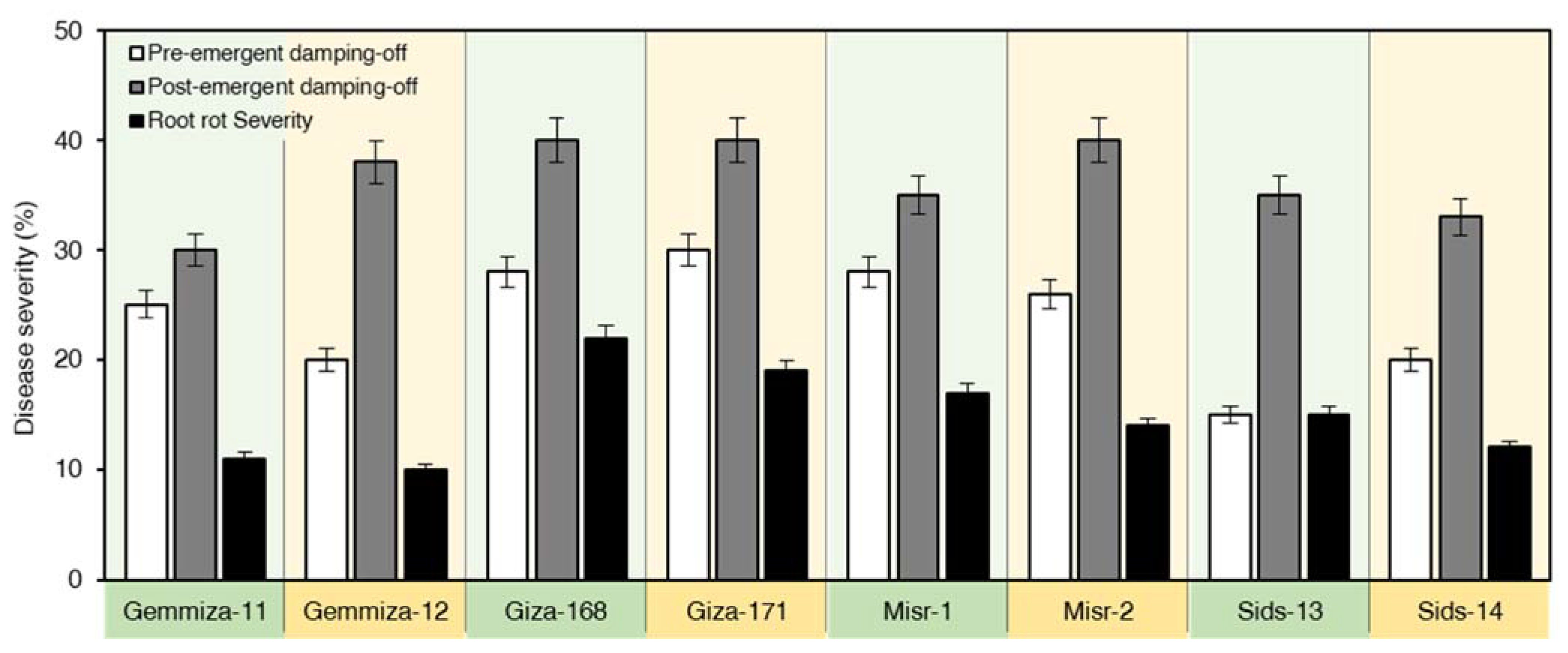
2.2. Responses of Wheat Cultivars to R. solani
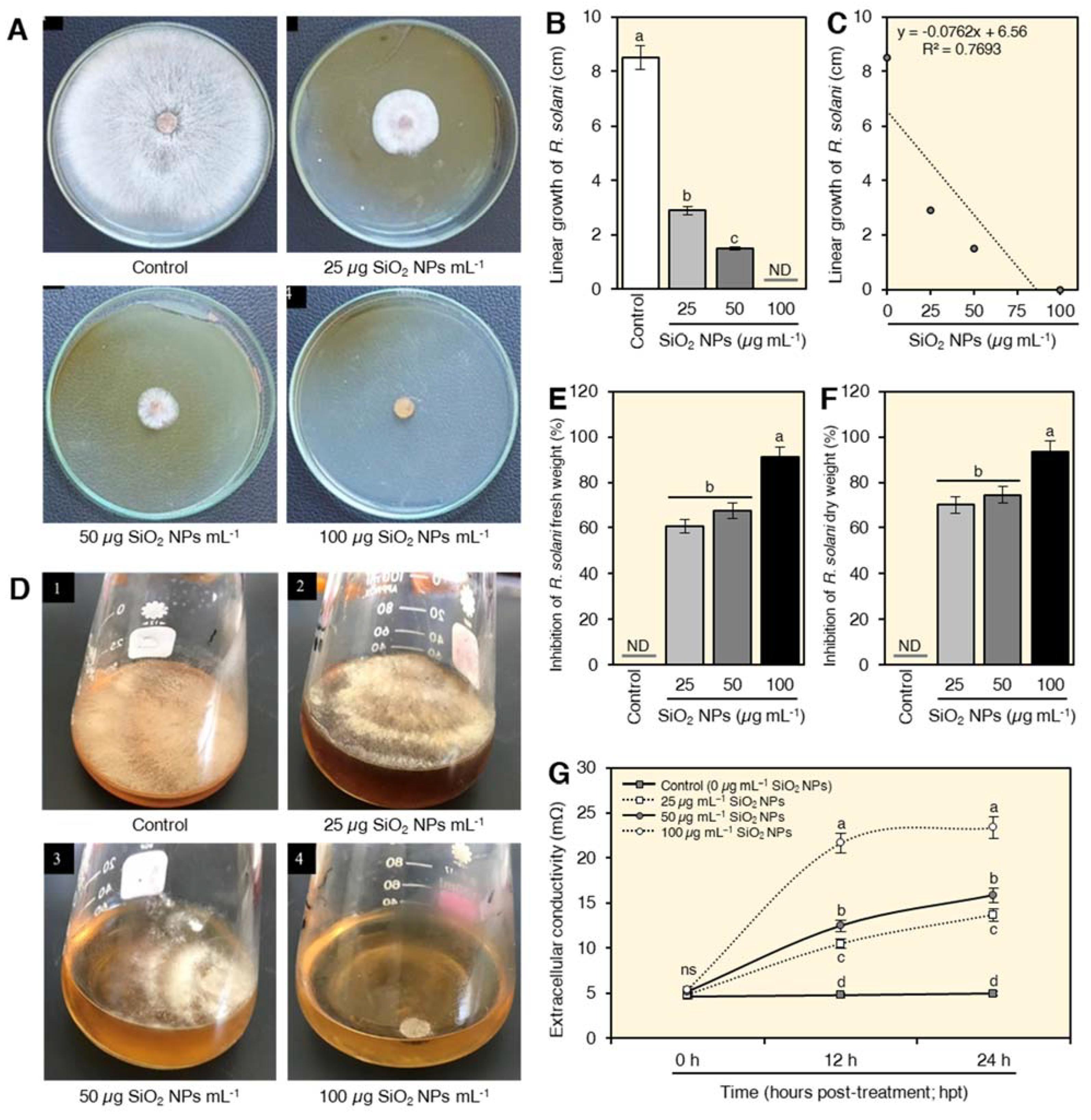
2.3. In Vitro Antifungal Activity of SiO2 NPs on R. solani
2.4. SiO2 NPs Increased the Extracellular Conductivity of Treated Fungal Suspension
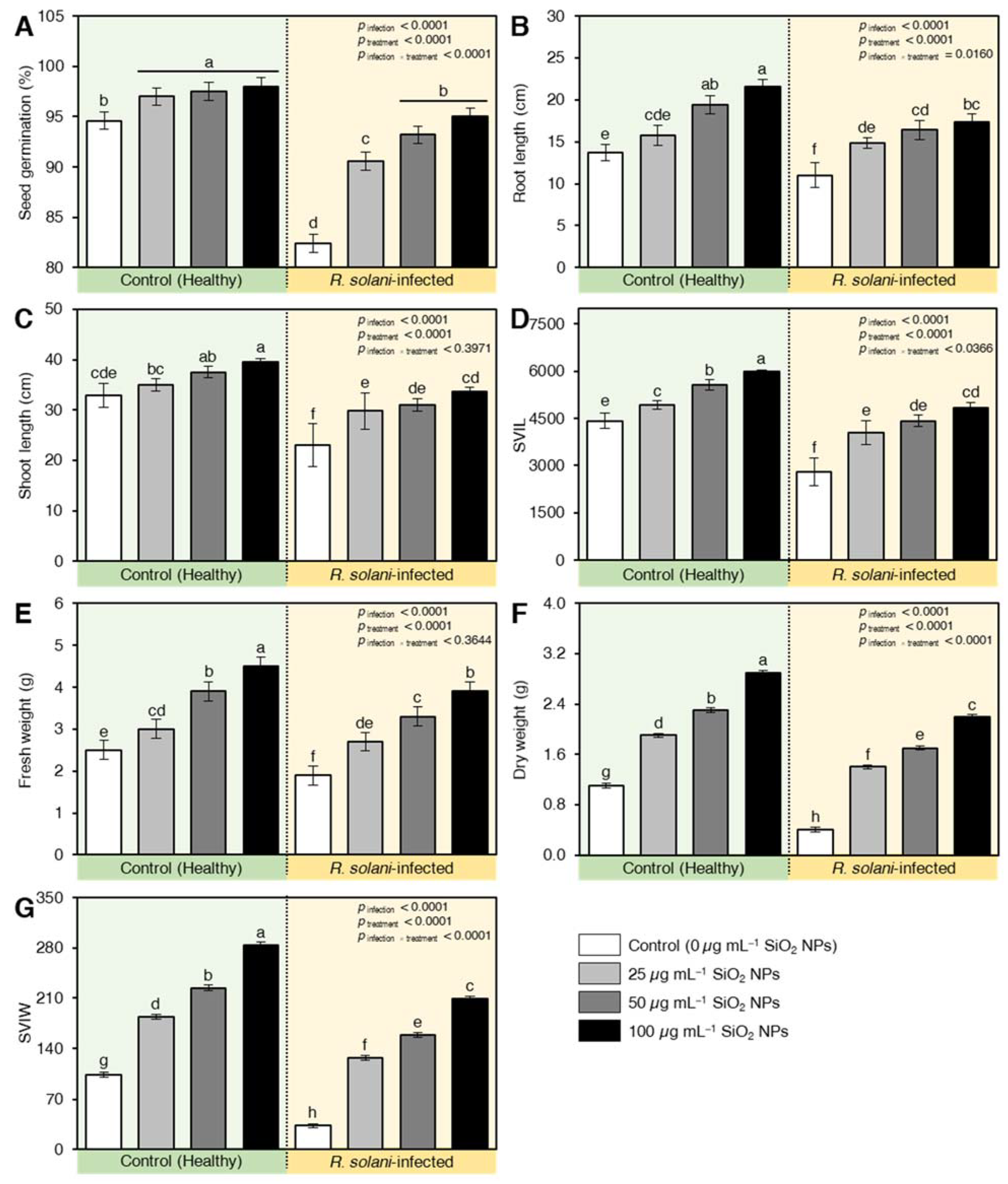
2.5. SiO2 NPs Enhanced the Germination, Root, and Shoot Length, and Vigor Indexes of Wheat Plants in Soil Inoculation with R. solani
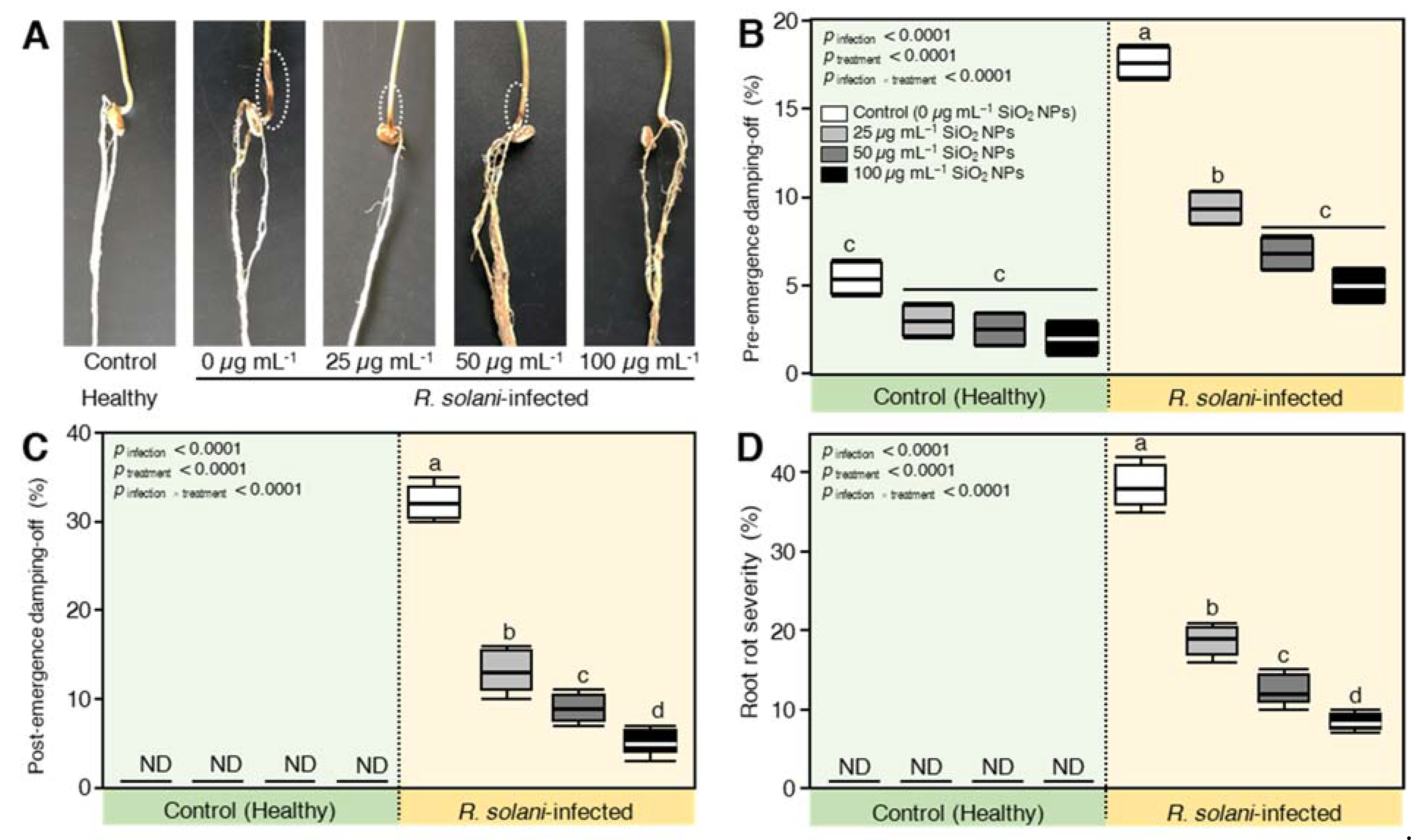
2.6. SiO2 NPs Application Reduced the Disease Parameters of R. solani—Inoculated Wheat
2.7. Effect of SiO2 NPs on the Biochemical Traits of Wheat Seedlings Inoculated with R. solani
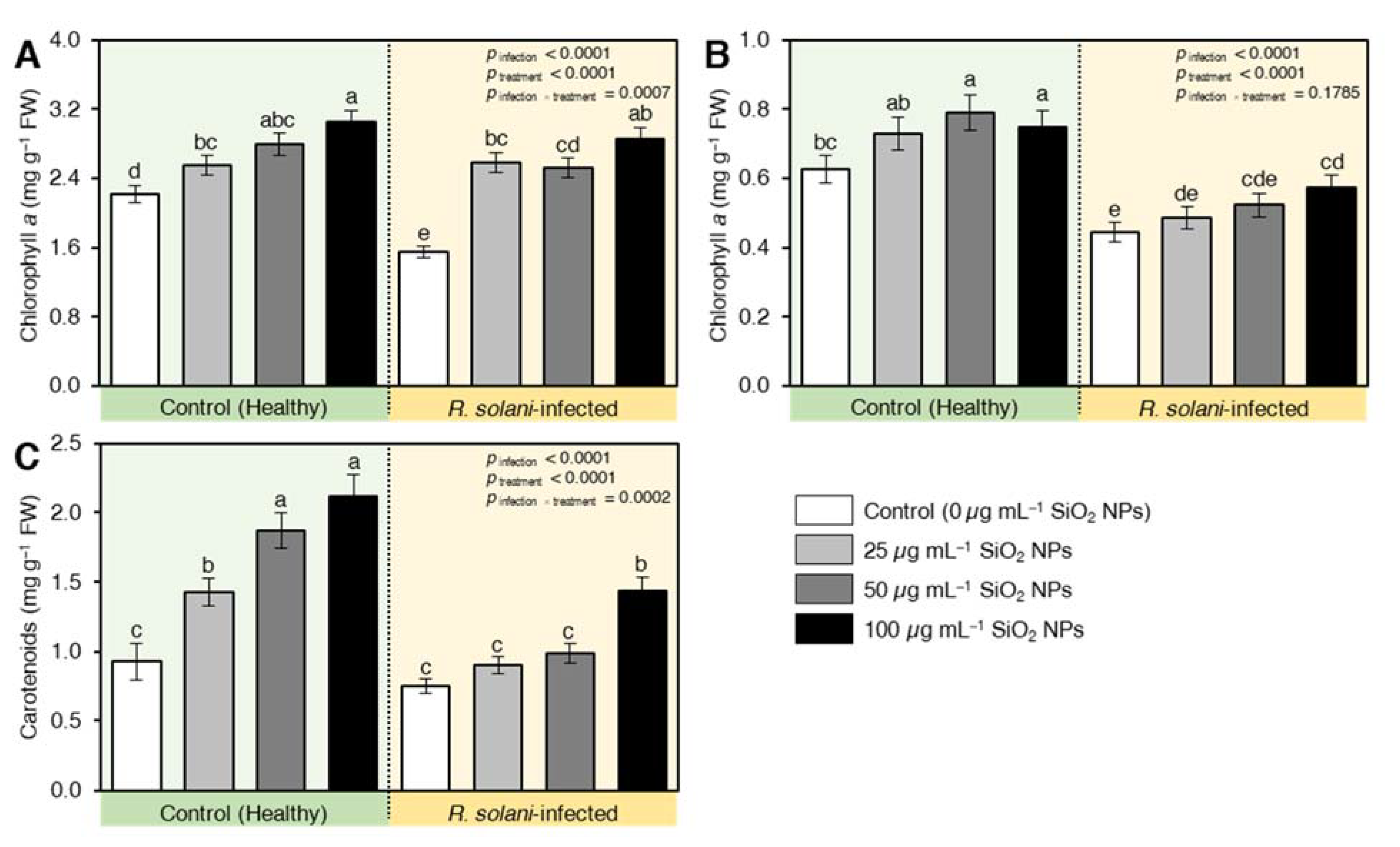
2.7.1. SiO2 NPs Enhanced the Content of the Photosynthetic Pigment in R. solani-Infected Plants
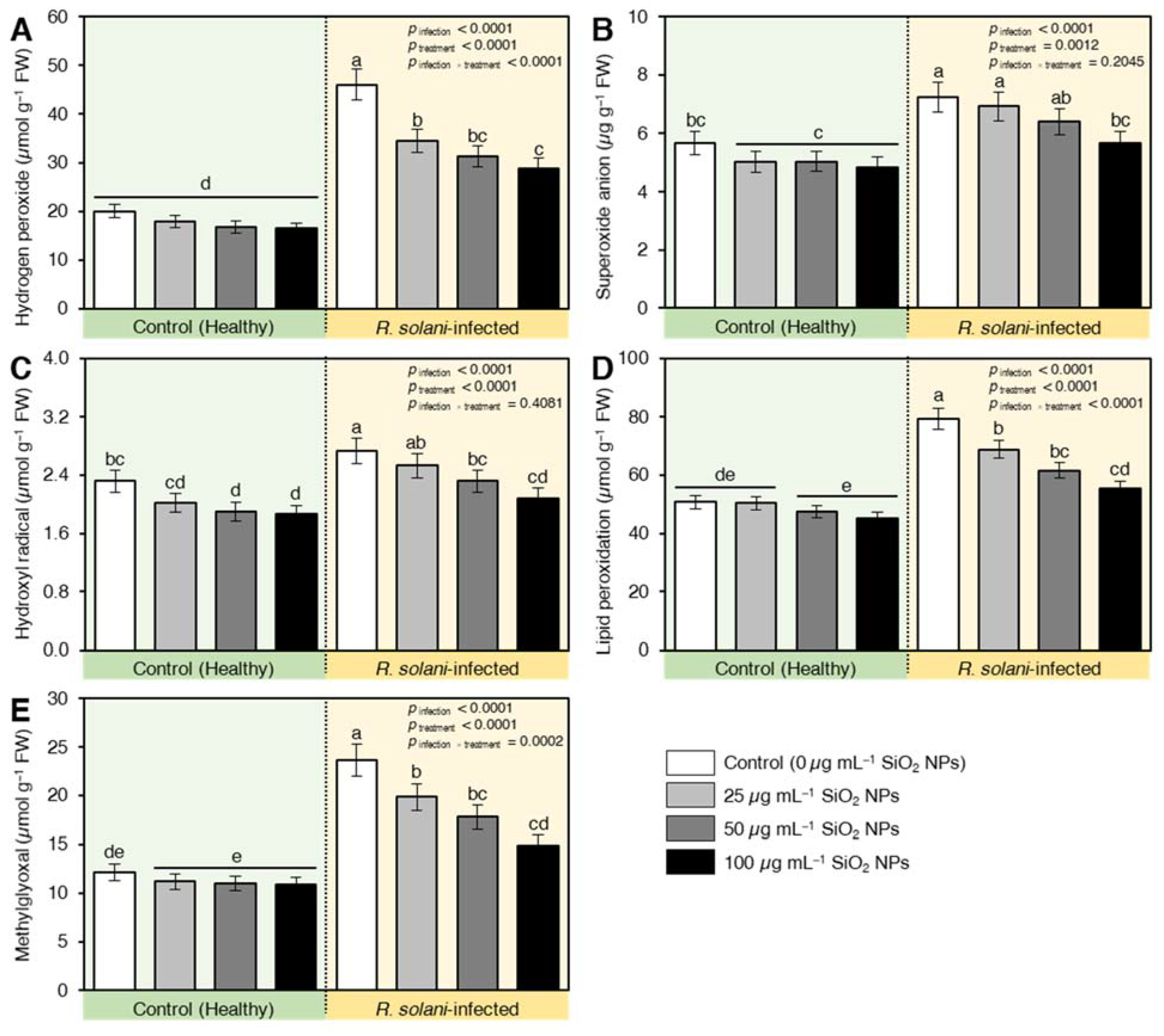
2.7.2. Treatment with SiO2 NPs Alleviated the Oxidative Stress in R. solani-Infected Plants
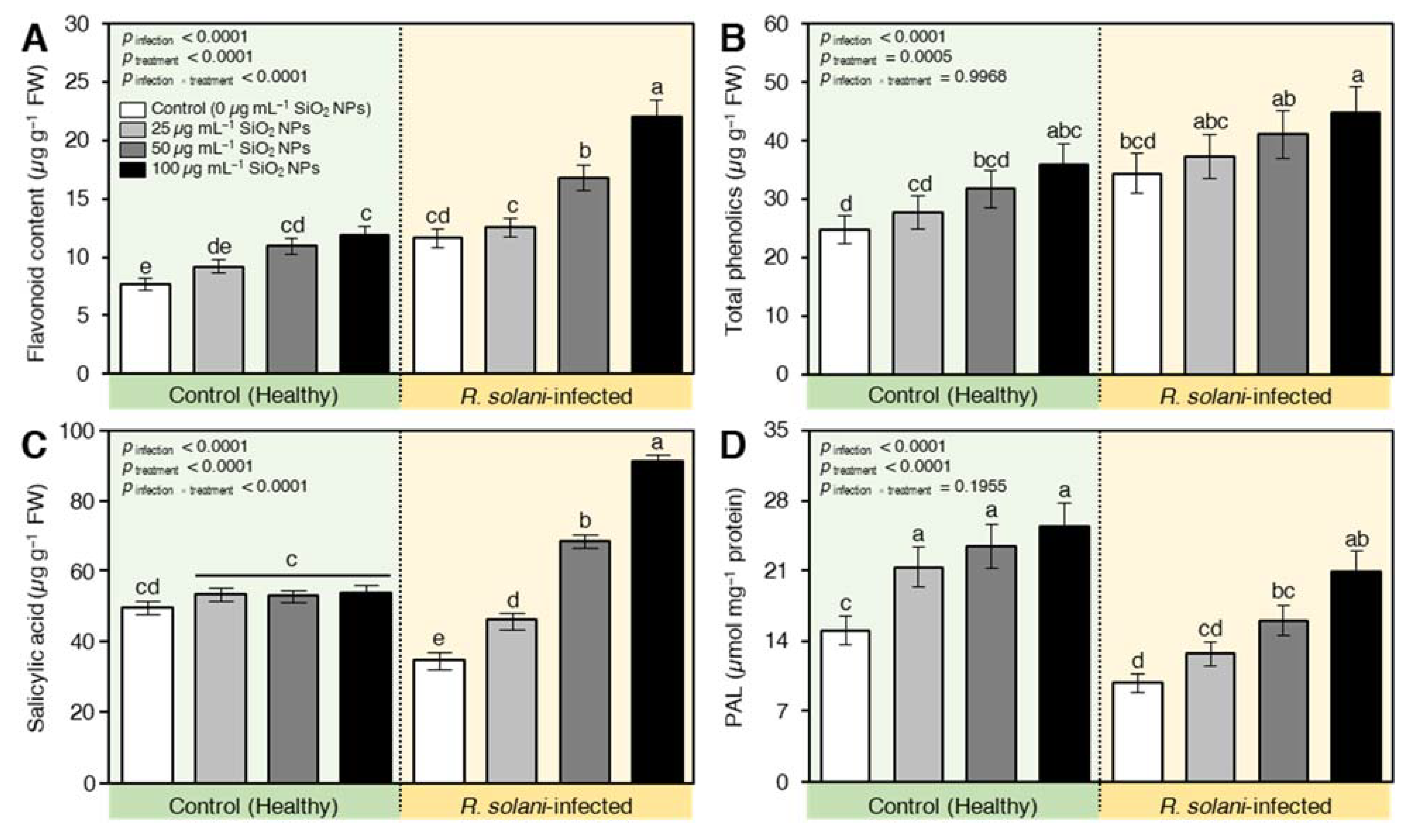
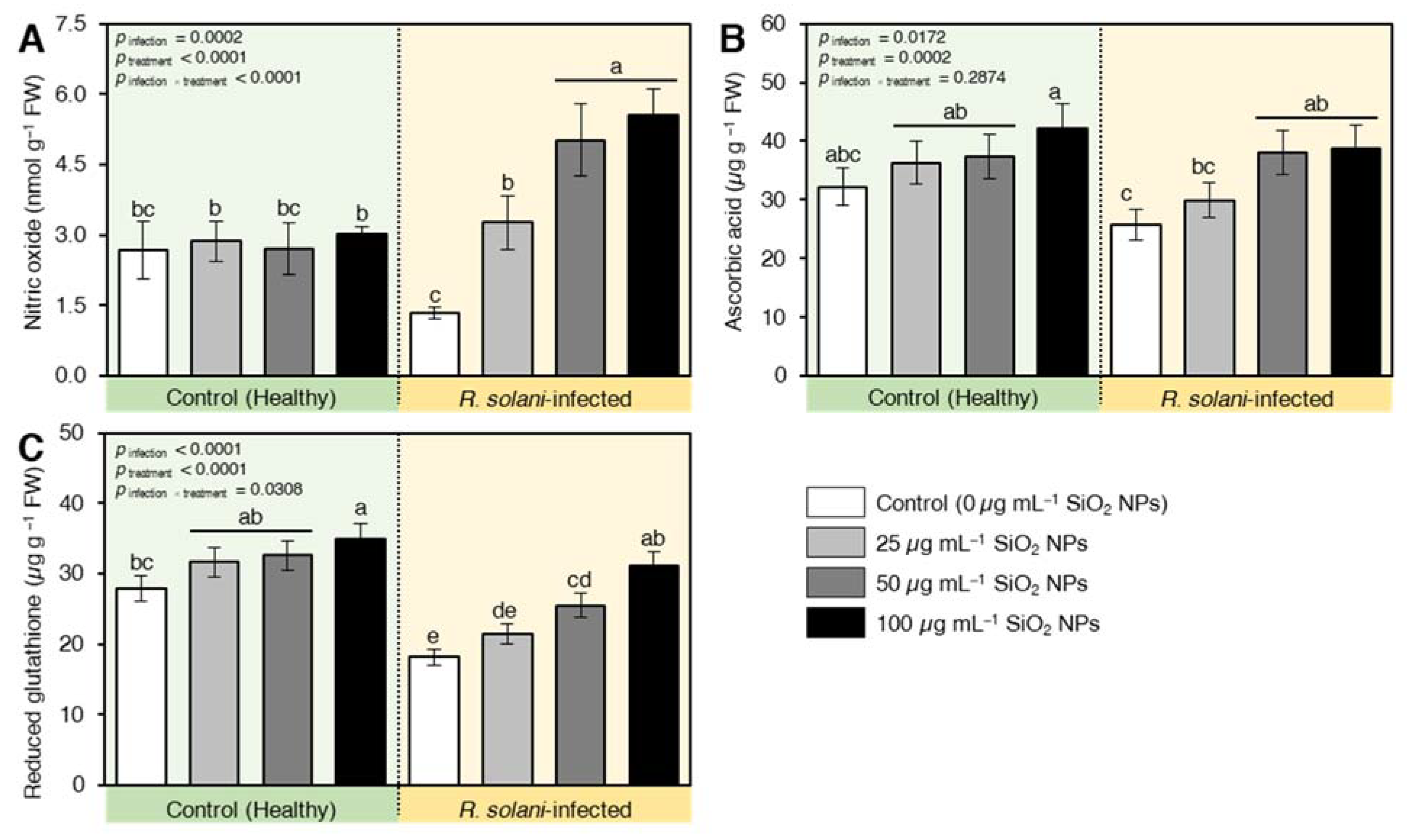
2.7.3. SiO2 NPs Induced the Accumulation of Defense-Related Compounds in R. solani-Infected Seedlings
2.7.4. SiO2 NPs Stimulated the Non-Enzymatic Antioxidant Machinery of R. solani-Infected Seedlings
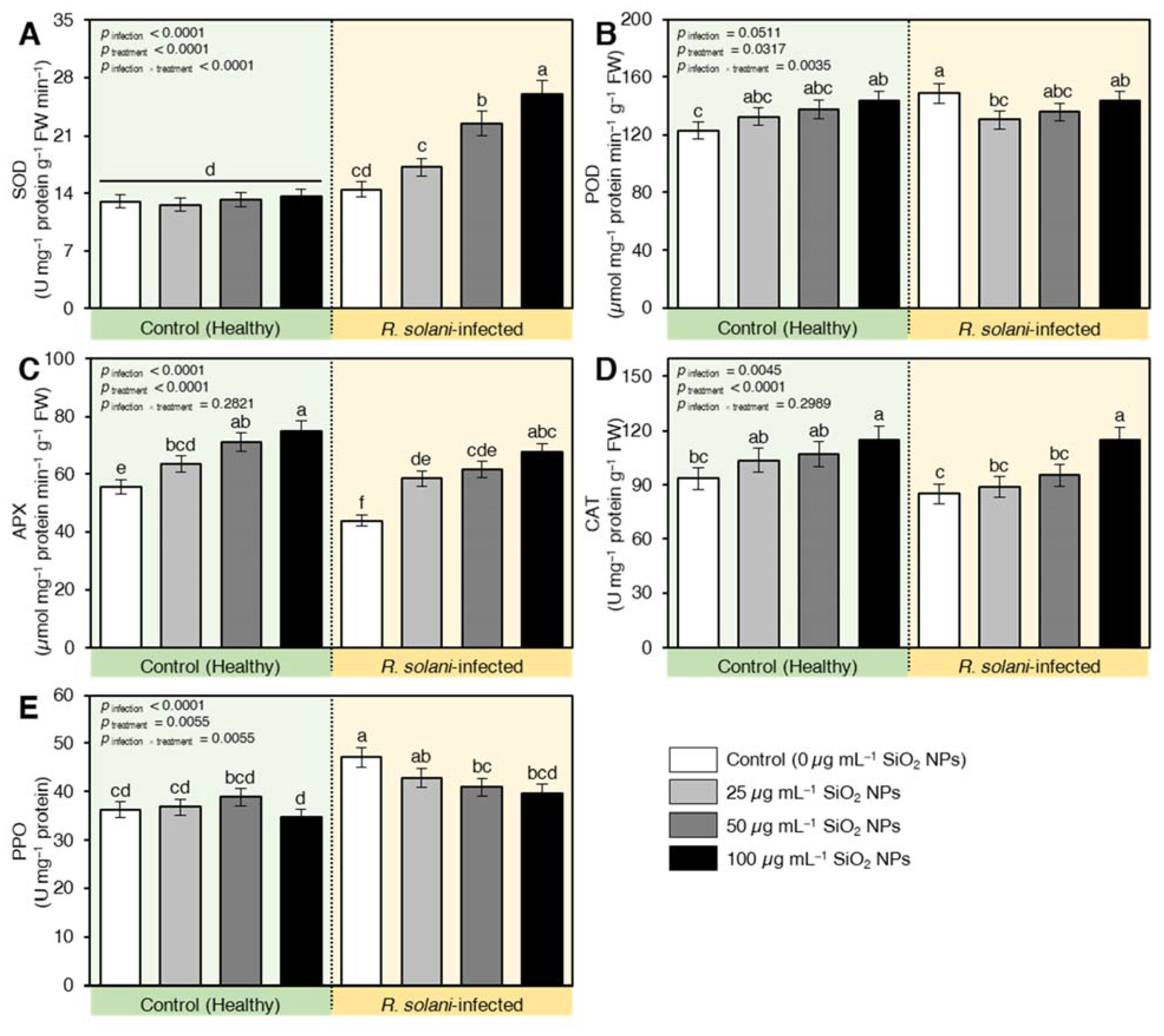
2.7.5. SiO2 NPs Induced the Enzymatic Antioxidant Machinery of R. solani-Infected Seedlings
3. Discussion
4. Materials and Methods
4.1. Biosynthesis of Silicon Dioxide (SiO2) Nanoparticles
4.1.1. Preparation of Aqueous Saffron Extraction
4.1.2. Synthesis of SiO2 Nanoparticles
4.1.3. Characterization of SiO2 Nanoparticles
Analysis of Ultraviolet Spectra
Fourier Transform Infrared Spectroscopy (FTIR)
X-ray Diffraction (XRD)
Transmission Electron Microscopy (TEM)
4.2. Pathogen Isolate and Pathogenicity Test
4.3. In Vitro Antifungal Activity of SiO2
4.3.1. Effect of SiO2 NPs on the Linear Growth of R. solani
4.3.2. Effect of SiO2 NPs on the Mycelium Fresh and Dry Weight of R. solani
4.3.3. Extracellular Conductivity
4.4. Effect of SiO2 NPs on Germination, Root and Shoot Length, and Vigor Indexes of R. solani-Infected Wheat Seedlings
4.5. Effect of SiO2 on Pre- and Post-Emergence Damping-Off and Root Rot of Wheat under Inoculation with R. solani
4.6. Stress-Associated Biochemical and Physiological Assessments
4.6.1. Pigments Content
4.6.2. Reactive Oxygen Species
4.6.3. Membrane Damage Criteria
4.6.4. Resistance-Related Compounds and Enzymes
Salicylic Acid (SA)
Phenolic Compounds
Nitric Oxide Content (NO)
Phenylalanine Ammonia-Lyase (PAL) and Polyphenol Oxidase (PPO) Activities
Flavonoids Content
4.6.5. Non-Enzymatic and Enzymatic Antioxidants
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 13 February 2020).
- González García, V.; Portal Onco, M.A.; Rubio Susan, V. Review. Biology and systematics of the form genus Rhizoctonia. Span. J. Agric. Res. 2006, 4, 55–79. [Google Scholar] [CrossRef] [Green Version]
- Hamada, M.; Yin, Y.; Chen, H.; Ma, Z. The escalating threat of Rhizoctonia cerealis, the causal agent of sharp eyespot in wheat. Pest Manag. Sci. 2011, 67, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Sneh, B.; Burpee, L.; Ogoshi, A. Identification of Rhizoctonia Species; Aps Press: St. Paul, MI, USA, 1991. [Google Scholar]
- Carling, D.E.; Kuninaga, S. DNA base sequence homology in Rhizoctonia solani Kühn: Inter- and intragroup relatedness of anastomosis group-9. Phytopathology 1990, 80, 1362. [Google Scholar] [CrossRef] [Green Version]
- Ogoshi, A.; Cook, R.J.; Bassett, E.N. Rhizoctonia species and anastomosis groups causing root rot of wheat and barley in the Pacific Northwest. Phytopathology 1990, 80. [Google Scholar] [CrossRef]
- Weller, J.L.; Terry, M.J.; Rameau, C.; Reid, J.B.; Kendrick, R.E. The phytochrome-deficient pcd1 mutant of pea is unable to convert heme to biliverdin IX[alpha]. Plant Cell 1996, 8, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Pumphrey, F.V. Influence of tillage and nitrogen fertilizer on Rhizoctonia root rot (bare patch) of winter wheat. Plant Dis. 1987, 71, 125. [Google Scholar] [CrossRef]
- Smiley, R.W. Impact of fungicide seed treatments on Rhizoctonia root rot, take-all, eyespot, and growth of winter wheat. Plant Dis. 1990, 74, 782. [Google Scholar] [CrossRef]
- Shu, C.; Sun, S.; Chen, J.; Chen, J.; Zhou, E. Comparison of different methods for total RNA extraction from sclerotia of Rhizoctonia solani. Electron. J. Biotechnol. 2014, 17, 50–54. [Google Scholar] [CrossRef] [Green Version]
- James Cook, R. Wheat root health management and environmental concern. Can. J. Plant Pathol. 1992, 14. [Google Scholar] [CrossRef]
- Ray, R.V.; Jenkinson, P.; Edwards, S.G. Effects of fungicides on eyespot, caused predominantly by Oculimacula acuformis, and yield of early-drilled winter wheat. Crop. Prot. 2004, 23, 1199–1207. [Google Scholar] [CrossRef]
- Kataria, H.R.; Hugelshofer, U.; Gisi, U. Sensitivity of Rhizoctonia species to different fungicides. Plant Pathol. 1991, 40, 203–211. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner, F.T. Germination response of loblolly Pine to temperature differentials on two-way thermogradient plante. J. Seed Technol. 1983, 8, 6–14. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria1. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabaghnia, N.; Janmohammadi, M. Graphic analysis of nano-silicon by salinity stress interaction on germination properties of lentil using the biplot method. Agric. For. 2014, 60, 24–40. [Google Scholar]
- Sabaghnia, N.; Janmohammadi, M. Effect of nano-silicon particles application on salinity tolerance in early growth of some lentil genotypes/Wpływ nanocząstek krzemionki na tolerancję zasolenia we wczesnym rozwoju niektórych genotypów soczewicy. Ann. UMCS Biol. 2015, 69. [Google Scholar] [CrossRef] [Green Version]
- Rawat, P.S.; Kumar, R.; Ram, P.; Pandey, P. Effect of nanoparticles on wheat seed germination and seedling growth. Int. J. Agric. Biosyst. Eng. 2018, 12, 13–16. [Google Scholar]
- Li, R.; He, J.; Xie, H.; Wang, W.; Bose, S.K.; Sun, Y.; Hu, J.; Yin, H. Effects of chitosan nanoparticles on seed germination and seedling growth of wheat (Triticum aestivum L.). Int. J. Biol. Macromol. 2019, 126, 91–100. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; YG, Z.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Curtis, F.; De Cicco, V.; Lima, G. Efficacy of biocontrol yeasts combined with calcium silicate or sulphur for controlling durum wheat powdery mildew and increasing grain yield components. Field Crop Res. 2012, 134. [Google Scholar] [CrossRef]
- Filho, J.A.W.; Duarte, H.S.S.; Rodrigues, F.A. Effect of foliar application of potassium silicate and fungicide on the severity of leaf rust and yellow leaf spot in wheat. Rev. Ceres 2013, 60, 726–730. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, F.A.; Dallagnol, L.J.; Duarte, H.S.S.; Datnoff, L.E. Silicon control of foliar diseases in monocots and dicots. In Silicon and Plant Diseases; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Côté-Beaulieu, C.; Chain, F.; Menzies, J.G.; Kinrade, S.D.; Bélanger, R.R. Absorption of aqueous inorganic and organic silicon compounds by wheat and their effect on growth and powdery mildew control. Environ. Exp. Bot. 2009, 65, 155–161. [Google Scholar] [CrossRef]
- Rodgers-Gray, B.S.; Shaw, M.W. Substantial reductions in winter wheat diseases caused by addition of straw but not manure to soil. Plant Pathol. 2000, 49, 590–599. [Google Scholar] [CrossRef]
- Pagani, A.P.S.; Dianese, A.C.; Café-Filho, A.C. Management of wheat blast with synthetic fungicides, partial resistance and silicate and phosphite minerals. Phytoparasit 2014, 42, 609–617. [Google Scholar] [CrossRef]
- Rodgers-Gray, B.S.; Shaw, M.W. Effects of straw and silicon soil amendments on some foliar and stem-base diseases in pot-grown winter wheat. Plant Pathol. 2004, 53. [Google Scholar] [CrossRef]
- Domiciano, G.P.; Rodrigues, F.A.; Guerra, A.M.N.; Vale, F.X.R. Infection process of Bipolaris sorokiniana on wheat leaves is affected by silicon. Trop. Plant Pathol. 2013, 38, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Azizian-Shermeh, O.; Valizadeh, M.; Taherizadeh, M.; Beigomi, M. Phytochemical investigation and phytosynthesis of eco-friendly stable bioactive gold and silver nanoparticles using petal extract of saffron (Crocus sativus L.) and study of their antimicrobial activities. Appl. Nanosci. 2020, 10, 2907–2920. [Google Scholar] [CrossRef]
- Alhumaydhi, F.A.; Khan, I.; Rauf, A.; Qureshi, M.N.; Aljohani, A.S.M.; Khan, S.A.; Khalil, A.A.; El-Esawi, M.A.; Muhammad, N. Synthesis, characterization, biological activities, and catalytic applications of alcoholic extract of saffron (Crocus sativus) flower stigma-based gold nanoparticles. Green Process. Synth. 2021, 10, 230–245. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef]
- Shokri, B.; Firouzjah, M.A.; Hosseini, S.I. FTIR analysis of silicon dioxide thin film deposited by metal organic-based PECVD. In Proceedings of the 19th International Plasma Chemistry Symposium, Bochum, Germany, 27–31 July 2009; pp. 1–4. Available online: www.ispc-conference.org (accessed on 10 December 2021).
- Verma, J.; Bhattacharya, A. analysis on synthesis of silica nanoparticles and its effect on growth of T. harzianum & Rhizoctonia species. Biomed. J. Sci. Tech. Res. 2018, 10. [Google Scholar] [CrossRef]
- Saravanan, S.; Dubey, R.S. Synthesis of SiO2 nanoparticles by sol-gel method and their optical and structural properties. Rom. J. Inf. Sci. Technol. 2020, 23, 105–112. [Google Scholar]
- Aziz, R.A.; Sopyan, I. Synthesis of TiO2-SiO2 powder and thin film photocatalysts by sol-gel method. Indian J. Chem. 2009, 48, 951–957. [Google Scholar]
- Arun Kumar, D.; Merline Shyla, J.; Xavier, F.P. Synthesis and characterization of TiO2/SiO2 nano composites for solar cell applications. Appl. Nanosci. 2012, 2, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Sooksaen, P.; Suttiruengwong, S.; Oniem, K.; Ngamlamiad, K.; Atireklapwarodom, J. Fabrication of porous bioactive glass-ceramics via decomposition of natural fibres. J. Met. Mater. Miner. 2017, 18, 85–91. [Google Scholar]
- Derbalah, A.; Shenashen, M.; Hamza, A.; Mohamed, A.; El Safty, S. Antifungal activity of fabricated mesoporous silica nanoparticles against early blight of tomato. Egypt. J. Basic Appl. Sci. 2018, 5, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Salem, H.F.; Eid, K.A.M.; Sharaf, M.A. Formulation and evaluation of silver nanoparticles as antibacterial and antifungal agents with a minimal cytotoxic effect. Int. J. Drug Deliv. 2011, 3, 293–304. [Google Scholar]
- Capeletti, L.B.; de Oliveira, L.F.; Gonçalves, K.d.A.; de Oliveira, J.F.A.; Saito, Â.; Kobarg, J.; dos Santos, J.H.Z.; Cardoso, M.B. Tailored silica–antibiotic nanoparticles: Overcoming bacterial resistance with low cytotoxicity. Langmuir 2014, 30, 7456–7464. [Google Scholar] [CrossRef]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of silicon on plant–pathogen interactions. Front. Plant Sci. 2017, 8, 701. [Google Scholar] [CrossRef] [Green Version]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s role in abiotic and biotic plant stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [Green Version]
- Weinhold, A.R.; Sinclair, J.B. Rhizoctonia solani: Penetration, colonization, and host response. Rhizoctonia Species Taxon. Mol. Biol. Ecol. Pathol. Dis. Control 1996, 163–174. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Yang, Y. Mechanisms of silicon-induced fungal disease resistance in plants. Plant Physiol. Biochem. 2021, 165, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Dorneles, K.R.; Dallagnol, L.J.; Pazdiora, P.C.; Rodrigues, F.A.; Deuner, S. Silicon potentiates biochemical defense responses of wheat against tan spot. Physiol. Mol. Plant Pathol. 2017, 97, 69–78. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Taher, M.A. Silicon induces resistance to postharvest rot of carrot caused by Sclerotinia sclerotiorum and the possible of defense mechanisms. Postharvest Biol. Technol. 2018, 140, 11–17. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Bhat, J.A.; Siddiqui, M.H.; Rinklebe, J.; Ahmad, P. Integration of silicon and secondary metabolites in plants: A significant association in stress tolerance. J. Exp. Bot. 2020, 71, 6758–6774. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Tahjib-Ul-arif, M.; Al Mamun Sohag, A.; Abdel Latef, A.A.H.; Ragaey, M.M. Mechanistic insight of allantoin in protecting tomato plants against ultraviolet c stress. Plants 2021, 10, 11. [Google Scholar] [CrossRef]
- Hoque, T.S.; Hossain, M.A.; Mostofa, M.G.; Burritt, D.J.; Fujita, M.; Tran, L.-S.P. Methylglyoxal: An Emerging Signaling Molecule in Plant Abiotic Stress Responses and Tolerance. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Vásquez, A.; Salinas, P.; Holuigue, L. Salicylic acid and reactive oxygen species interplay in the transcriptional control of defense genes expression. Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Abdelrhim, A.S.; Dawood, M.F.A.; Galal, A.A. Hydrogen peroxide-mixed compounds and/or microwave radiation as alternative control means against onion seed associated pathogens, Aspergillus niger and Fusarium oxysporum. J. Plant Pathol. 2021, 1, 1–15. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M.; Tomar, N.S.; Shrivastava, M. Potassium induces positive changes in nitrogen metabolism and antioxidant system of oat (Avena sativa L cultivar Kent). J. Plant Interact. 2015, 10, 211–223. [Google Scholar] [CrossRef]
- Bagy, H.M.M.K.; Hassan, E.A.; Nafady, N.A.; Dawood, M.F.A. Efficacy of arbuscular mycorrhizal fungi and endophytic strain Epicoccum nigrum ASU11 as biocontrol agents against blackleg disease of potato caused by bacterial strain Pectobacterium carotovora subsp. atrosepticum PHY7. Biol. Control 2019, 134, 103–113. [Google Scholar] [CrossRef]
- Rodrigues, F.Á.; Benhamou, N.; Datnoff, L.E.; Jones, J.B.; Bélanger, R.R. Ultrastructural and cytochemical aspects of silicon-mediated rice blast resistance. Phytopathology 2003, 93. [Google Scholar] [CrossRef] [Green Version]
- Suriyaprabha, R.; Karunakaran, G.; Kavitha, K.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Application of silica nanoparticles in maize to enhance fungal resistance. IET Nanobiotechnol. 2014, 8, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Nehela, Y.; Hijaz, F.; Elzaawely, A.A.; El-Zahaby, H.M.; Killiny, N. Citrus phytohormonal response to Candidatus Liberibacter asiaticus and its vector Diaphorina citri. Physiol. Mol. Plant Pathol. 2018, 102, 24–35. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. Multiple phytohormonal signaling mediates citrus response to the bacterial pathogen Candidatus Liberibacter asiaticus. Phytopathology 2018, 108, 10. [Google Scholar]
- Nehela, Y.; Killiny, N. Melatonin is involved in citrus response to the pathogen huanglongbing via modulation of phytohormonal biosynthesis. Plant Physiol. 2020, 184, 2216–2239. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Azooz, M.M. Insights into the oxidative status and antioxidative responses of germinating broccoli (Brassica oleracea var. italica L.) seeds in tungstate contaminated water. Chemosphere 2020, 261, 127585. [Google Scholar] [CrossRef] [PubMed]
- Bittner, N.; Trauer-Kizilelma, U.; Hilker, M. Early plant defence against insect attack: Involvement of reactive oxygen species in plant responses to insect egg deposition. Planta 2017, 245, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Fleck, A.T.; Nye, T.; Repenning, C.; Stahl, F.; Zahn, M.; Schenk, M.K. Silicon enhances suberization and lignification in roots of rice (Oryza sativa). J. Exp. Bot. 2011, 62, 2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asgari, F.; Majd, A.; Jonoubi, P.; Najafi, F. Effects of silicon nanoparticles on molecular, chemical, structural and ultrastructural characteristics of oat (Avena sativa L.). Plant Physiol. Biochem. PPB 2018, 127, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Gao, D.; Luo, S.; Zeng, R.; Yang, J.; Zhu, X. Physiological and cytological mechanisms of silicon-induced resistance in rice against blast disease. Physiol. Plant. 2008, 134, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Si, J.; Römheld, V. Silicon uptake and transport is an active process in Cucumis sativus. New Phytol. 2005, 167, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Sallam, N.M.A.; AbdElfatah, H.A.S.; Dawood, M.F.A.; Hassan, E.A.; Mohamed, M.S.; Khalil Bagy, H.M.M. Physiological and histopathological assessments of the susceptibility of different tomato (Solanum lycopersicum) cultivars to early blight disease. Eur. J. Plant Pathol. 2021, 160, 541–556. [Google Scholar] [CrossRef]
- Constabel, C.P.; Barbehenn, R. Defensive roles of polyphenol oxidase in plants. Induc. Plant Resist. Herbiv. 2008, 253–270. [Google Scholar] [CrossRef]
- Nagaonkar, D.; Gaikwad, S.; Rai, M. Catharanthus roseus leaf extract-synthesized chitosan nanoparticles for controlled in vitro release of chloramphenicol and ketoconazole. Colloid Polym. Sci. 2015, 293, 1465–1473. [Google Scholar] [CrossRef]
- Hossain, A.; Abdallah, Y.; Ali, M.A.; Masum, M.M.I.; Li, B.; Sun, G.; Meng, Y.; Wang, Y.; An, Q. Lemon-fruit-based green synthesis of zinc oxide nanoparticles and titanium dioxide nanoparticles against soft rot bacterial pathogen Dickeya dadantii. Biomolecules 2019, 9, 863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masum, M.I.; Siddiqa, M.M.; Ali, K.A.; Zhang, Y.; Abdallah, Y.; Ibrahim, E.; Qiu, W.; Yan, C.; Li, B. Biogenic synthesis of silver nanoparticles using Phyllanthus emblica fruit extract and its inhibitory action against the pathogen Acidovorax oryzae strain RS-2 of rice bacterial brown stripe. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Ogunyemi, S.O.; Abdallah, Y.; Zhang, M.; Fouad, H.; Hong, X.; Ibrahim, E.; Masum, M.M.I.; Hossain, A.; Mo, J.; Li, B. Green synthesis of zinc oxide nanoparticles using different plant extracts and their antibacterial activity against Xanthomonas oryzae pv. oryzae. Artif. Cells Nanomed. Biotechnol. 2019, 47, 341–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alejandre, A.; Medina, F.; Salagre, P.; Fabregat, A.; Sueiras, J.E. Characterization and activity of copper and nickel catalysts for the oxidation of phenol aqueous solutions. Appl. Catal. B Environ. 1998, 18, 307–315. [Google Scholar] [CrossRef]
- Atallah, O.O.; Osman, A.; Ali, M.A.S.; Sitohy, M. Soybean β-conglycinin and catfish cutaneous mucous p22 glycoproteins deteriorate sporangial cell walls of Pseudoperonospora cubensis and suppress cucumber downy mildew. Pest Manag. Sci. 2021, 77, 3313–3324. [Google Scholar] [CrossRef] [PubMed]
- Atallah, O.; Yassin, S. Aspergillus spp. eliminate Sclerotinia sclerotiorum by imbalancing the ambient oxalic acid concentration and parasitizing its sclerotia. Environ. Microbiol. 2020, 22, 5265–5279. [Google Scholar] [CrossRef] [PubMed]
- Botha, A.; Denman, S.; Lamprecht, S.C.; Mazzola, M.; Crous, P.W. Characterisation and pathogenicity of Rhizoctonia isolates associated with black root rot of strawberries in the Western Cape Province, South Africa. Australas. Plant Pathol. 2003, 32, 195–201. [Google Scholar] [CrossRef]
- Raju, K.; MK, N. Survey and assessment for the post-harvest diseases of onion in North-Eastern Karnataka. Karnataka J. Agric. Sci. 2007, 20, 164–165. [Google Scholar]
- Beale, R.; Phillion, D.; Headrick, J.; O’reilly, P.; Cox, J. MON65500: A unique fungicide for the control of take-all in wheat. In Proceedings of the 1998 Brighton Conference, Farnham, UK.—Pests and Diseases, Brighton, UK, 16–19 November 1998; Farnham Survey, British Crop Protection Council: Farnham, UK, 1998; pp. 343–350. [Google Scholar]
- Joshi, S.M.; De Britto, S.; Jogaiah, S.; Ito, S. Mycogenic selenium nanoparticles as potential new generation broad spectrum antifungal molecules. Biomolecules 2019, 9, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EL-Nagar, A.; Elzaawely, A.A.; Taha, N.A.; Nehela, Y. The antifungal activity of gallic acid and its derivatives against Alternaria solani, the causal agent of tomato early blight. Agronomy 2020, 10, 1402. [Google Scholar] [CrossRef]
- Nehela, Y.; Taha, N.A.; Elzaawely, A.A.; Xuan, T.D.; Amin, M.A.; Ahmed, M.E.; El-Nagar, A. Benzoic acid and its hydroxylated derivatives suppress early blight of tomato (Alternaria solani) via the induction of salicylic acid biosynthesis and enzymatic and nonenzymatic antioxidant defense machinery. J. Fungi 2021, 7, 663. [Google Scholar] [CrossRef]
- Li, Q.; Jiang, Y.; Ning, P.; Zheng, L.; Huang, J.; Li, G.; Jiang, D.; Hsiang, T. Suppression of Magnaporthe oryzae by culture filtrates of Streptomyces globisporus JK-1. Biol. Control 2011, 58, 139–148. [Google Scholar] [CrossRef]
- Newell, S.Y.; Statzell-Tallman, A. Factors for conversion of fungal biovolume values to biomass, carbon and nitrogen: Variation with mycelial ages, growth conditions, and strains of fungi from a salt marsh. Oikos 1982, 39, 261. [Google Scholar] [CrossRef]
- Manhas, R.K.; Kaur, T. Biocontrol potential of Streptomyces hydrogenans strain DH16 toward Alternaria brassicicola to control damping off and black leaf spot of Raphanus sativus. Front. Plant Sci. 2016, 7, 1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Yang, H.; Wu, F.; Cheng, J. Reduced chilling injury in cucumber by nitric oxide and the antioxidant response. Food Chem. 2011, 127, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Madhava Rao, K.V.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Gilbert, R.P.; Brandt, R.B. Spectrophotometric determination of methyl glyoxal with 2,4-dinitrophenylhydrazine. Anal. Chem. 2002, 47, 2418–2422. [Google Scholar] [CrossRef] [PubMed]
- Warrier, R.R.; Paul, M.; Plant, M.V.V.-G. Estimation of salicylic acid in Eucalyptus leaves using spectrophotometric methods. Genet. Plant Physiol. 2013, 3, 90–97. [Google Scholar]
- Kofalvi, S.A.; Nassuth, A. Influence of wheat streak mosaic virus infection on phenylpropanoid metabolism and the accumulation of phenolics and lignin in wheat. Physiol. Mol. Plant Pathol. 1995, 47, 365–377. [Google Scholar] [CrossRef]
- Ding, A.H.; Nathan, C.F.; Stuehr, D.J. Release of reactive nitrogen intermediates and reactive oxygen intermediates from mouse peritoneal macrophages. Comparison of activating cytokines and evidence for independent production. J Immunol. 1988, 141, 2407–2412. [Google Scholar]
- Hu, X.; Neill, S.J.; Cai, W.; Tang, Z. NO-mediated hypersensitive responses of rice suspension cultures induced by incompatible elicitor. Chin. Sci. Bull. 2003, 48, 358–363. [Google Scholar] [CrossRef]
- Havir, E.A.; Hanson, K.R. L-Phenylalanine ammonia-lyase (maize and potato). Evidence that the enzyme is composed of four subunits. Biochemistry 2002, 12, 1583–1591. [Google Scholar] [CrossRef]
- Kumar, K.B.; Khan, P.A. Peroxidase and polyphenol oxidase in excised ragi (Eleusine corocana cv PR 202) leaves during senescence. Indian J. Exp. Biol. 1982, 412–416. [Google Scholar]
- Zou, Y.; Lu, Y.; Wei, D. Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro. J. Agric. Food Chem. 2004, 52, 5032–5039. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Jagota, S.K.; Dani, H.M. A new colorimetric technique for the estimation of vitamin C using Folin phenol reagent. Anal. Biochem. 1982, 127. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. Oxidative stress and antioxidative systems: Recipes for successful data collection and interpretation. Plant. Cell Environ. 2016, 39. [Google Scholar] [CrossRef] [Green Version]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Silva, E.N.; Silveira, J.A.G.; Aragão, R.M.; Vieira, C.F.; Carvalho, F.E.L. Photosynthesis impairment and oxidative stress in Jatropha curcas exposed to drought are partially dependent on decreased catalase activity. Acta Physiol. Plant. 2018, 41, 1–12. [Google Scholar] [CrossRef]
- Zaharieva, T.; Yamashita, K.; Matsumoto, H. Iron deficiency induced changes in ascorbate content and enzyme activities related to ascorbate metabolism in cucumber roots. Plant Cell Physiol. 1999, 40, 273–280. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelrhim, A.S.; Mazrou, Y.S.A.; Nehela, Y.; Atallah, O.O.; El-Ashmony, R.M.; Dawood, M.F.A. Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat Against Rhizoctonia solani. Plants 2021, 10, 2758. https://doi.org/10.3390/plants10122758
Abdelrhim AS, Mazrou YSA, Nehela Y, Atallah OO, El-Ashmony RM, Dawood MFA. Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat Against Rhizoctonia solani. Plants. 2021; 10(12):2758. https://doi.org/10.3390/plants10122758
Chicago/Turabian StyleAbdelrhim, Abdelrazek S., Yasser S. A. Mazrou, Yasser Nehela, Osama O. Atallah, Ranya M. El-Ashmony, and Mona F. A. Dawood. 2021. "Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat Against Rhizoctonia solani" Plants 10, no. 12: 2758. https://doi.org/10.3390/plants10122758