1. Introduction
High yields in coffee crops have been achieved using superior genotypes developed over decades of genetic improvement [
1].
Coffea canephora Pierre ex A. Froehner is a highly heterozygous species, and distinct characteristics are expressed in the different genotypes [
2]. Among them, we can mention drought tolerance [
2], accumulation of biomass and nutrients [
3,
4], vegetative growth [
5], and absorption and efficiency of nutrient use [
6,
7]. This broad diversity enables the selection and introduction of superior genotypes into the production chain.
Highly productive genotypes have high nutritional requirements [
8], and they may or may not vary between strains. The length of the reproductive cycle [
3,
4], anatomy of the plant organ [
6], and phenological phases [
9] are some examples of mechanisms responsible for the nutritional variation in coffee genotypes [
10]. They can vary in concentration and have a greater requirement of one or more elements [
11,
12,
13].
In general, in
C. canephora plants, nitrogen (N), calcium (Ca), and potassium (K) are absorbed in greater amounts, while phosphorus (P), magnesium (Mg), and sulfur (S) are required less [
4,
14,
15]. In leaves, there is a decreased concentration of macronutrients considered mobile in the plant during the reproductive phase. This decrease results from the migration of photoassimilates and mineral elements to drains, such as flowers and fruits, after the degradation of cellular compounds [
16]. In several varieties of conilon coffee, Oliosi et al. [
10] and Covre et al. [
15] observed greater demand in the following order: N, P, K, Ca, S, Mg, B, Fe, Mn, Zn, Cu in the reproductive stages, in anthesis, pre-flowering, grain filling, and fruiting.
The plant’s nutritional imbalance can promote the excessive accumulation of elements [
10], triggering a series of undesirable metabolic processes, which can vary according to the type of organ in the plant and may have an influence on the quality of the beverage [
17]. The plant’s nutritional status is an essential tool in managing coffee production, determined by the nutritional content in different plant tissues. Although the evaluations of organs from coffee trees at different phenological stages are essential tools to determine the proper fertilization management, the existing reference values of the critical level and sufficiency range for nutritional diagnosis do not consider the genetic diversity among genotypes [
8].
Several plant organs can provide subsidies for adequate nutritional management. Flower analysis enables us to advance in nutritional corrections during vegetative growth, avoiding further losses [
18,
19]. The grains and straw analysis provides essential answers about calculating the nutrients responsible for crop productivity [
15]. The most used organ of the coffee plant in these analyses is the leaves. They are physiologically active organs, present in all phenological cycles of the crop, favoring the monitoring of the plant’s nutritional status [
18,
19].
Therefore, the objective of this study was to determine the genetic variability for the concentration of nutrients in different plant organs throughout the reproductive cycle of nine genotypes of conilon coffee (Coffea canephora).
3. Discussion
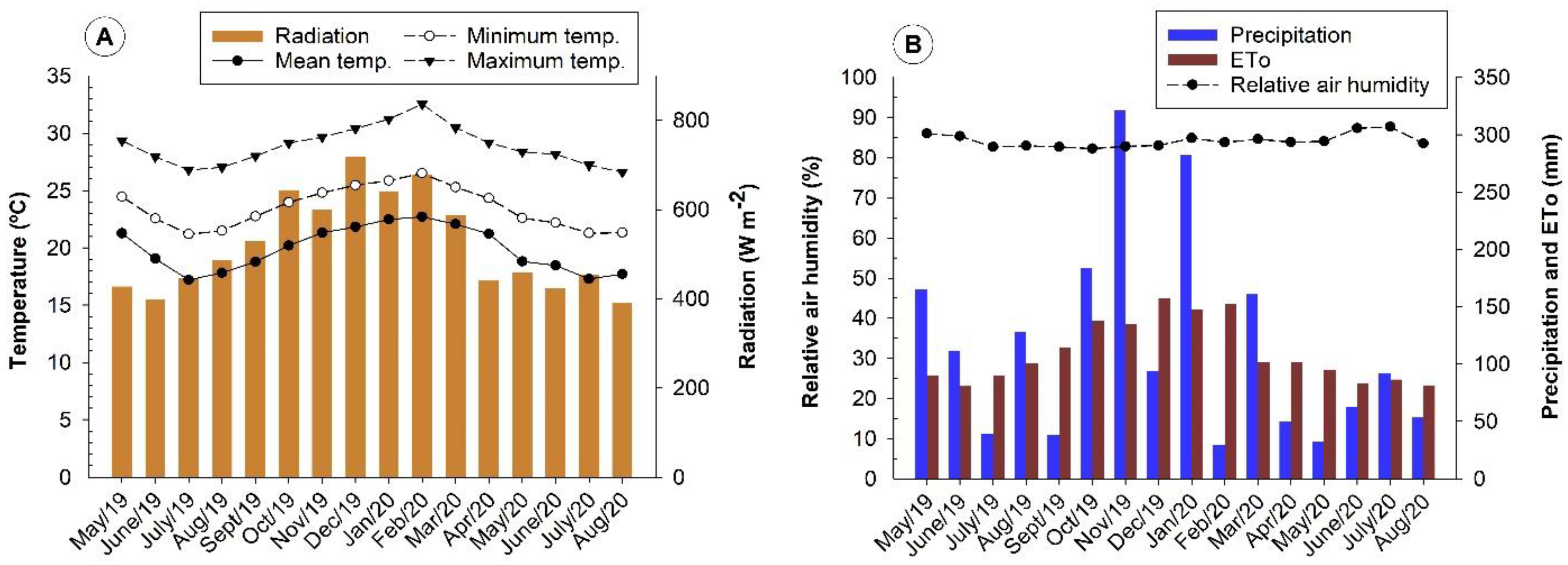
An occasional increase in nutrient concentrations may have been favored by the increase in precipitation and temperature in the experiment region after September (
Figure 5). Coffee plants can have their metabolism reduced in the dry period, due to mild temperatures. At the same time, growth rates are faster in the rainy season and with higher temperatures, due to increased metabolism, as noted by Amaral et al. [
21].
Genetic parameters, such as CVe, CVg, and H
2, are essential tools used in improvement programs, with which possible promising genotypes are identified, contributing to the selection process in genetic improvement [
22]. CVe, for example, aims to estimate how much the environment influences the expression of the evaluated characteristics of a given experiment [
23]. According to studies by Ferreira et al. [
24], CVg is a parameter that can allow inferences about the variability present in the population, and, thus, establishes levels of variability in specific genotypes. The higher the value, the more heterogeneous the evaluated genotypes [
22]. The highest CVg values in this study were observed for straw, followed by grains and flowers in some nutrients. However, for most nutrients, the variability was at levels considered low for genetic improvement research [
25].
A large part of the nutrients showed high H
2 values, above 80%. High heritability is associated with how much the phenotypic values represent the genotypic values of individuals [
26]. Heritability is one of the most important genetic parameters to verify the genetic control of traits and the efficiency of the selection process in plant improvement. In contrast to most results, the concentration of nutrients in the flowers showed a more significant influence from the environment, a result represented by the lower values of H
2 for the concentration of many nutrients in this plant organ.
C. canephora plants that are highly productive also have greater nutritional demands. These demands vary according to genetic characteristics and plant tissue [
4,
27]. Further, the nutrient concentration varies according to the edaphic conditions, season, plant age, and genotype maturation cycle [
4,
10], indicating that some genotypes can absorb or translocate more nutrients than others.
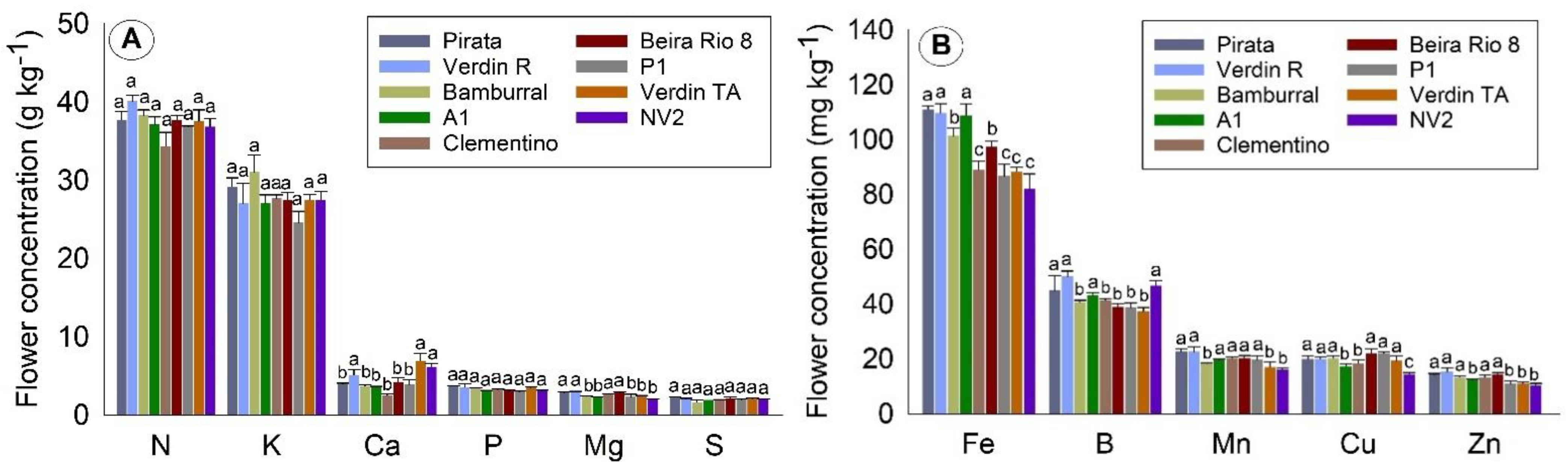
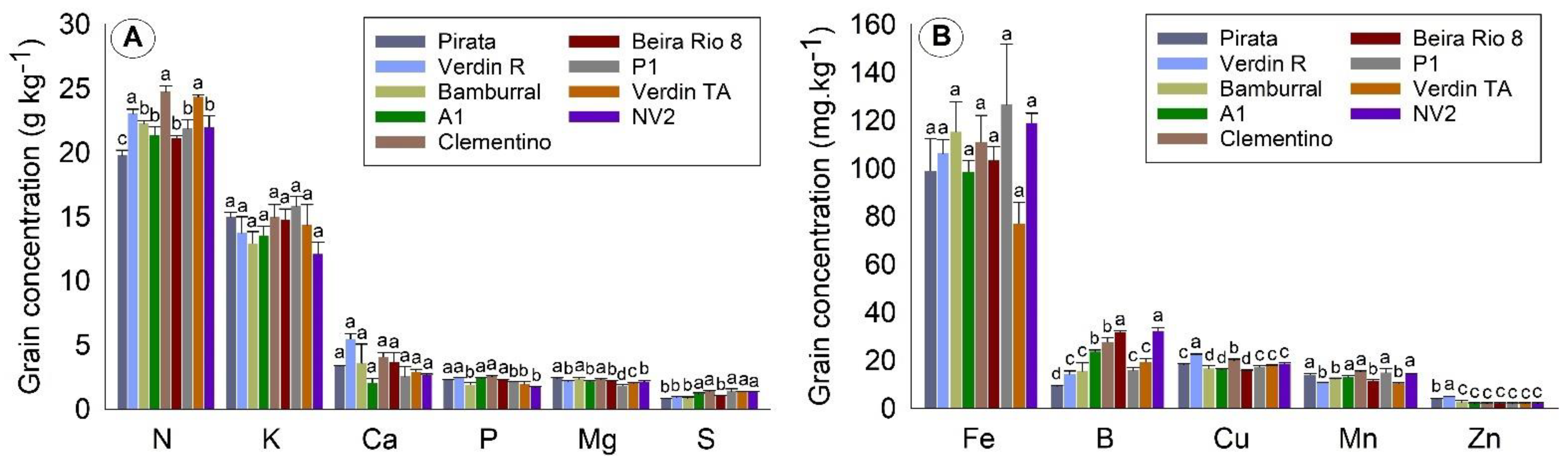
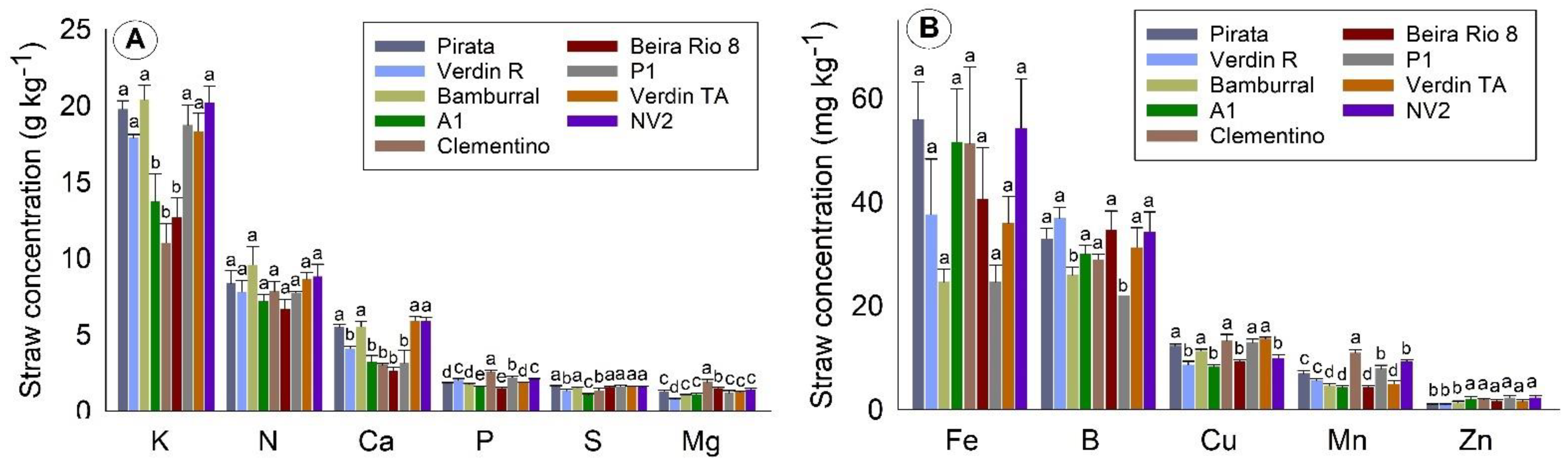
Considering all tissues, the orders of concentration of the macronutrients and micronutrients found in this work were N > K > Ca > P = Mg = S and Fe > B > Mn > Cu > Zn, respectively. N was the most required nutrient during the evaluation period, around 40 g kg−1 in flowers, and 20 g kg−1 in leaves and grains. The exception was straw, in which the most commonly found nutrient was K (about 12 g kg−1).
In flowers, N and K were found in higher concentrations (about 35 and 30 g kg
−¹, respectively) than the other macronutrients that, together, concentrated less than 20 g kg
−¹. While P is not the most required nutrient during the production cycle, it plays a vital role in this phase, and plays essential roles in plants, acting in the functions of energy metabolism in the form of ATP, which is essential in the growth of plant organs [
16].
In the present study, no differences were observed between genotypes for the N, K, P, and S concentration in flowers. Supporting the results, Laviola et al. [
28] also observed similar values for these macronutrients among cultivars of
C. arabica. As for the micronutrients, mainly Fe, differences between genotypes are observed; four genotypes (Clementino, P1, Verdin TA, and NV2) had about 20% less Fe than the genotypes Pirata, Verdin R, and A1.
The development of flower buds occurs concomitantly with the vegetative growth of plants. They can compete for water, nutrients and carbohydrates, and can be used as indicators for nutritional diagnosis, as they are a substantial drain within the plant [
28]. There are nutrients, such as Ca and B, that follow the transpiration flow; however, concerning the functionality of the conducting vessels, they are much smaller in flowers than in leaves, making transport difficult. Thus, during the formation of flower buds, it is interesting that the plant has already received and absorbed the necessary amount of nutrients, since the absorption of nutrients starts before floral anthesis.
C. canephora genotypes have high genetic variability due to factors such as allogamy, which makes nutrient absorption, or translocation, and flowering differentiated [
2,
19].
The highest nutrient values found in concentrations of flowers, grains, straw, and leaves can determine favorable conditions for identifying the diversity within these genotypes [
29]. The present study can be compared to that by Covre et al. [
15], who evaluated
C. canephora grains and straws, obtaining the macronutrients K, N, and Ca as the highest in straws, and N and K in grains. For micronutrients, the authors also found Fe and B to be the most accumulated in straws and grains. These higher levels of nutrients are believed to be due to the fertilization in previous months, contributing to seed filling. Moreover, in this production stage, fertilization has already been carried out according to the fertilization recommendation for the specific crop [
30].
The more significant amount of N in coffee grains than in straw, and the more significant amount of K in straw, may be due to several factors, such as being linked to the requirement of the plant itself for higher concentrations of N for production [
31]. The high concentration of K in straw is believed to be due to the pre-existing high content in leaves and fruits [
32]. Thus, coffee straw can be used in crops as soil organic matter, due to its high concentrations of K and N.
The micronutrient Fe had the highest concentration in
C. canephora grains and straw, which confirms the studies by Covre et al. [
15] and Partelli et al. [
5], which found that Fe is the most accumulated micronutrient in conilon coffee. When evaluating genotypes with different maturation cycles, Marré et al. [
3] also found a higher Fe concentration in
C. canephora genotypes. B was the second most accumulated, followed by Mn. These results are similar to those found by Covre et al. [
15] in the grains and straws of irrigated and non-irrigated coffee trees.
After flowering, and until grain maturation in coffee genotypes, a reduction in N and K concentrations in leaves is noted, reflecting the translocation of the photoassimilates needed to ensure fruit quality formation [
10].
Leaf N concentration was relatively within what is expected for the crop in the flowering and grain filling stages [
8]. According to Martins et al. [
7], there is a direct relationship between the absorption of nutrients and the plant cycle, considering that the maturation of the fruits requires more significant amounts of metabolites, which intensifies the transport of photoassimilates to the fruits.
Oscillations in the concentrations of N and K in the leaves were also observed between the genotypes Pirata and Bamburral, which are known to have late and medium/late maturation, with a decrease in leaf concentrations later compared to the other genotypes. These fluctuations can be attributed to the maturation conditions of the grains, conditioned to the genetic characteristics of each genotype. According to Partelli et al. [
4,
5], the highest nutrient absorption speed was verified in fruits with early maturation and higher growth rates. Genotypes may present nutritional variations throughout the year, with influence from the type of maturation (early, late, and medium) [
4,
10], climatic factors, or the phenological phase itself. Thus, different genotypes need nutrient concentrations at different periods, but the period is almost always similar to their phenological phase [
4,
30]. Oliosi et al. [
10] studied the seasonal variation relating to leaf concentrations, obtaining higher N concentrations in early and medium maturation genotypes in the first samplings, and lower in late-maturing genotypes, with a higher peak in November for the latter.
In leaves, the macronutrient P is required more in the periods before flowering, in June, July, and the grain filling stages, with a higher concentration at the end of January [
18], which is similar to the studies of the present work. In season 3 (February), there was an increase in the concentration of this nutrient. While P is the macronutrient required in the smallest amount [
14], it plays a vital role in photosynthesis and protein phosphorylation [
16].
As for the concentrations of Ca and Mg present in the leaves, both showed similarities in the seasons evaluated [
14]. Ca showed a reduction in the fruit set phases for leaf contents and Mg, which, although uncommon, may reflect the redistribution of nutrients to the fruits [
9]. Oliosi et al. [
10] stated, in their studies, that the highest concentrations of Ca were in October and November, and Mg decreased in September, followed by an increase.
The S contents in the leaves are found to have higher concentrations for grain filling and seed filling periods, as in the studies by Oliosi et al. [
10] and Silva et al. [
25]. The concentration of this macronutrient showed variations due to the maturation of the genotypes. In early, medium, late, and super late cycle conilon coffee, Partelli et al. [
4] and Oliosi et al. [
10] also obtained this variation, with higher concentrations of S in the leaf for the genotypes V8 and V12, which are considered to have an early maturation cycle. According to Tomaz et al. [
33], different genotypes may differ concerning the absorption, translocation, and efficiency of S. This effect is due to affinities in absorption by the root system, mobility in the xylem, and phloem vessels [
34].
Thus, the seasonality observed in the concentration of macronutrients in leaves varies according to the seasons and genotypes evaluated in this study. The hottest periods of the year and early spring coincide with the highest concentrations of the macronutrients N, P, and K. After September, there is an increase in precipitation (
Figure 5), favoring greater nutrient absorption, which justifies the higher concentrations found for the macronutrients Ca, Mg, and S. Furthermore, in this season, according to Marenco and Lopes [
35], the increase in temperature causes acceleration of the respiratory activities of plants and better metabolism of the root system.
The micronutrient Fe is considered to be the most accumulated by conilon coffee plants. According to Bragança et al. [
36], approximately 80% of Fe accumulates in chloroplasts, which have a vital role in photosynthesis and chlorophyll production. According to Covre et al. [
9], low micronutrient levels in coffee are observed at the end of the crop cycle. In this study, this situation occurs in season 4, the fruit maturation period. According to Oliosi et al. [
10], the Fe concentrations in leaves showed higher concentrations soon after it rained.
In studies with coffee trees in the Western Amazon, Dubberstein et al. [
37] noted that Cu showed slight variation and linear behavior, remaining constant since the first evaluation. This characteristic situation of Cu also occurred in the present study.
In studies with two evaluation periods, Marré et al. [
3] observed that the concentrations of Mn in the leaves were reduced, which could be due to the greater demand of the micronutrient in the development of the fruits during this period, the maturation period. Oliosi et al. [
10] observed a reduction in B in August, mainly due to flower formation.
In most micronutrients (Fe, Zn, B, and Mn), lower levels in the first seasons (fruit set phase) and a high range could be verified, characterized as expansion and seed filling of the fruits. This behavior is characteristic of conilon coffee trees in Brazil, as already observed in studies in the northern region of Espírito Santo [
3] and the Western Amazon [
37].
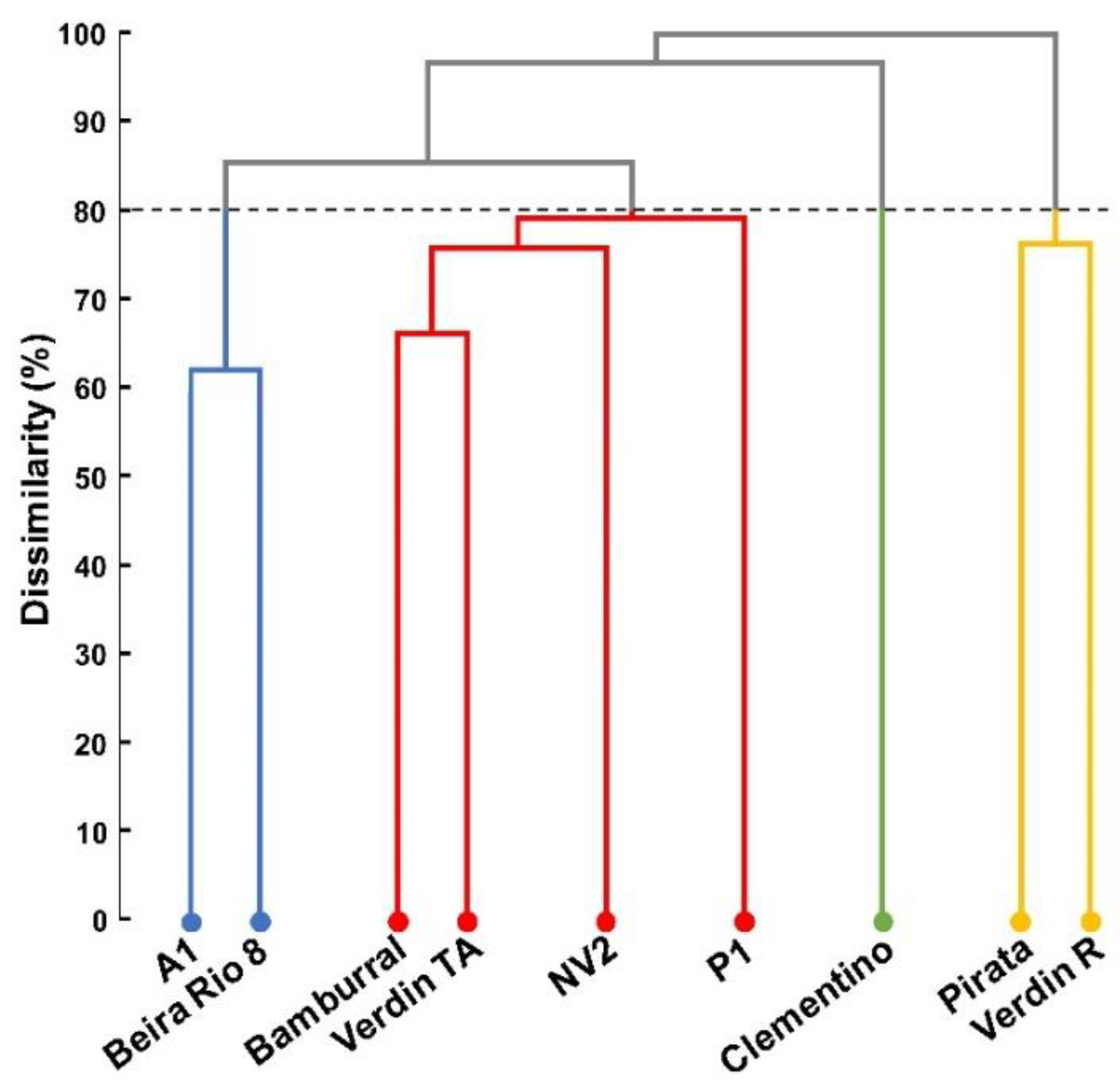
The formation of groups by the dendrogram indicates variability among the
C. canephora genotypes for nutritional contents of leaves, flowers, grains, and straw. Silva et al. [
25], Gomes et al. [
8], and Martins et al. [
2] also observed divergence between the conilon coffee genotypes for nutrient concentration. These studies are essential for improvement programs, being used to select genotypes with superior characteristics.
Of the four groups formed in the present study, the one composed of Pirata and Verdin R presented the highest concentrations of nutrients in the evaluated tissues, and the slightest oscillation in the nutritional dynamics in their leaves. Pirata and Verdin R have similar maturation cycles, early and medium/early, respectively. Two other genotypes, which form another group, also share the following similar maturation cycles: medium cycle, A1; medium/early cycle, Beira Rio 8.
Several correlations were observed for the concentration of nutrients between the evaluated plant organs. Most of the correlations were positive and involved the concentration of nutrients in the leaves and other plant organs. The concentration of nutrients in the leaves is commonly used for the nutritional diagnosis of coffee plants [
8]. Thus, an adequate concentration of nutrients in the coffee leaves must be maintained, especially during periods of greater nutritional demand, such as flowering and grain filling [
19,
28].
While this study has addressed the nutritional similarities in different organs of the plant, and has, thus, obtained dissimilarity-based groups, factors such as leaf morphology, and roots and plant architecture were not addressed. These morphological attributes act directly on nutrient absorption, photosynthetic rates, and radiation resistance, and can affect the nutritional status of each plant’s tissues [
38]. Thus, deepening the morphological parameters may bring greater understanding to the nutritional discussion of conilon coffee genotypes.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}