
GIS-Facilitated Seed Germination and Multifaceted Evaluation of the Endangered Abies marocana Trab. (Pinaceae) Enabling Conservation and Sustainable Exploitation

 , , , , , , ,
, , , , , , , 
Abstract
:1. Introduction
2. Results
2.1. Overview of the Potential of Abies marocana in Economic Sectors
2.1.1. Ornamental-Horticultural Potential
2.1.2. Medicinal-Cosmetic and Agro-Alimentary Potential
2.1.3. Feasibility and Readiness Assessments for Sustainable Exploitation
2.2. Ecological Profiling of Abies marocana
2.2.1. Temperature-Related Attributes
2.2.2. Precipitation-Related Attributes
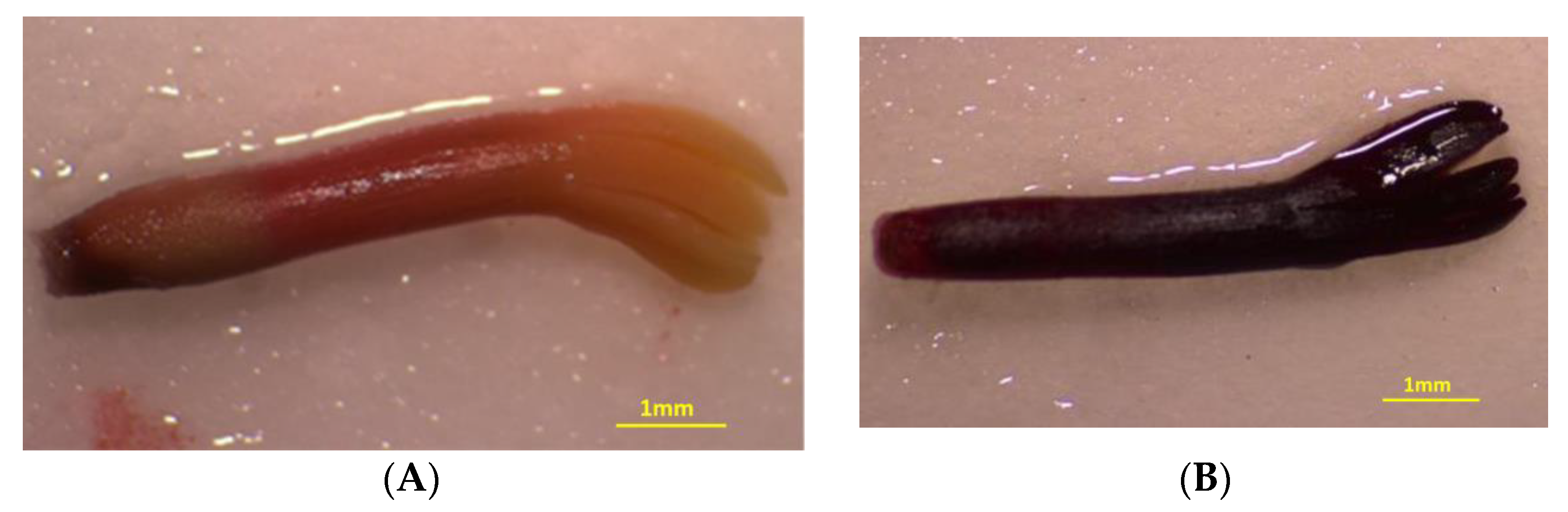
2.3. Seed Germination Tests
3. Discussion
3.1. Potential, Feasibility, and Readiness for Value Chain Creation
3.2. Seed Germination Requirements for Conservation and Sustainable Exploitation
4. Materials and Methods
4.1. Multifaceted Evaluation in Economic Sectors
4.2. Distribution Mapping and GIS Ecological Profiling
4.3. Seed Collection and Storage
4.4. Seed Treatment
4.5. Germination Tests
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vidakovic, V. Abies Mill.—Fir—Pinaceae. In Conifers: Morphology and Variation; Brekalo, B., Ed.; Graficki Zadov: Hrvatske, Yugoslavia, 1991; pp. 53–116. [Google Scholar]
- Kohli, R.K.; Batish, D.R.; Singh, H.P. Important tree species: Forests and Forest Plants. In Encyclopedia of Life Support System (EOLSS); Owens, J.N., Lund, H.G., Eds.; EOLSS Pub.: Oxford, UK, 2003; pp. 1–39. [Google Scholar]
- Redžić, S.S. The ecological aspect of ethnobotany and ethnopharmacology of population in Bosnia and Herzegovina. Coll. Antropol. 2007, 31, 869–890. [Google Scholar] [PubMed]
- Aussenac, G. Ecology and ecophysiology of circum-Mediterranean firs in the context of climate change. Ann. For. Sci. 2002, 59, 823–832. [Google Scholar] [CrossRef]
- Alaoui, M.L.; Knees, S.; Gardner, M. Abies pinsapo var. marocana. The IUCN Red List of Threatened Species 2011; e.T34126A9841418. 2011. Available online: https://www.iucnredlist.org/species/34126/9841418 (accessed on 10 November 2021).
- Baumer, M. Le sapin du Maroc. Biol. Forêt 1977, 24, 343–354. Available online: http://documents.irevues.inist.fr/bitstream/handle/2042/21153/RFF_1977_5_343.pdf (accessed on 6 July 2021). [CrossRef] [Green Version]
- Hampe, A.; Jump, A. Climate relicts: Past, present, future. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Esteban, L.G.; De Palacios, P. Pinsapo forests: Past, present and future. Bois Forêt Trop. 2007, 292, 39–47. [Google Scholar] [CrossRef]
- Alaoui, A.; Laaribya, S.; Ayan, S.; Ghallab, A.; López-Tirado, J. Modelling spatial distribution of endemic Moroccan fir (Abies marocana Trabut) in Talassemtane National Park, Morocco. Austrian J. For. Sci. 2021, 138, 73–94. [Google Scholar]
- Navarro-Cerrillo, R.M.; Manzanedo, R.D.; Rodriguez-Vallejo, C.; Gazol, A.; Palacios-Rodríguez, G.; Camarero, J.J. Competition modulates the response of growth to climate in pure and mixed Abies pinsapo subsp. maroccana forests in northern Morocco. For. Ecol. Manag. 2020, 459, 117847. [Google Scholar] [CrossRef]
- Ghallab, A. Modélisation de la Distribution Potentielle Actuelle des Espèces Forestières Majeures Dans le Rif Occidental du Maroc et Prédiction de L’impact du Changement Climatique. Doctoral Thesis, Université Abdelmalek Essaâdi, Tétouan, Marocco, 2018. [Google Scholar]
- Navarro-Cerrillo, R.M.; Camarero, J.J.; Manzanedo, R.D.; Sánchez-Cuesta, R.; Lopez Quintanilla, J.; Sánchez Salguero, R. Regeneration of Abies pinsapo within gaps created by Heterobasidion annosum-induced tree mortality in southern Spain. IForest 2014, 7, 209. [Google Scholar] [CrossRef] [Green Version]
- Boukil, A. Etude de l’évolution de la gestion de la sapinière de Talassemtane (Rif centro-occidental, Maroc). Méditerranéenne 1998, 20, 145–152. [Google Scholar]
- Dadach, M. Recherche des Conditions Optimales de la Germination des Graines de Quelques Labiées du Mont de Tessala (Ouest Algérien) et Perspectives de Conservation. Ph.D. Thesis, Université Djillali Liabes de Sidi Bel Abbes, Sidi Bel Abbes, Algérie, 2013. [Google Scholar]
- Belmehdi, O.; El Harsal, A.; Benmoussi, M.; Laghmouchi, Y.; Skali Senhaji, N.; Abrini, J. Effect of light, temperature, salt stress and pH on seed germination of medicinal plant Origanum elongatum (Bonnet) Emb. & Maire. Biocatal. Agric. Biotechnol. 2018, 16, 126–131. [Google Scholar] [CrossRef]
- Flores-Enríquez, V.; Castillo, G.; Collazo-Ortega, M. Experimental seed germination for ex situ conservation of Mexican Podostemaceae. Bot. Sci. 2019, 97, 413–422. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Sarropoulou, V.; Krigas, N.; Maloupa, E.; Tsoktouridis, G. GIS-facilitated effective propagation protocols of the Endangered local endemic of Crete Carlina diae (Rech. f.) Meusel and A. Kástner (Asteraceae): Serving ex-situ conservation needs and its future sustainable exploitation as an ornamental. Plants 2020, 9, 1465. [Google Scholar] [CrossRef] [PubMed]
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M. Exploring the potential of neglected local endemic plants of three Mediterranean regions in the ornamental sector: Value chain feasibility and readiness timescale for their sustainable exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Benzyane, M.; Aafi, A.; Sbay, H.; El Antry, S.; Yassine, M.; Ilmen, R.; Ghaïloule, D. Les Ecosystèmes Naturels Marocains et les Changements Climatiques: La Résilience Ecologique à L’épreuve; Collection Maroc Nature; Centre de Recherche Forestière: Rabat, Morocco, 2010; 88p. [Google Scholar]
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Sharrock, S.; Krigas, N. Ex-situ conservation of single-country endemic plants of Tunisia and northern Morocco (Mediterranean coast and Rif region) in seed banks and botanic gardens worldwide. Kew Bull. 2020, 75, 46. [Google Scholar] [CrossRef]
- Khabbach, A.; Libiad, M.; El Haissoufi, M.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Menteli, V.; Vokou, D.; Tsoktouridis, G. Electronic commerce of the endemic plants of Northern Morocco (Rif region and Mediterranean coast) and Tunisia over the Internet. Bot. Sci. 2021, 100, 1–29, in press. [Google Scholar] [CrossRef]
- Belaqziz, R.; Romane, A.; Abbad, A. Salt stress effects on germination, growth and essential oil content of an endemic thyme species in Morocco (Thymus maroccanus Ball). J. Appl. Sci. Res. 2009, 5, 858–863. Available online: https://www.cabdirect.org/cabdirect/abstract/20093204278 (accessed on 15 September 2021).
- Abbad, A.; Belaqziz, R.; Bekkouche, K.; Markouk, M. Influence of temperature and water potential on laboratory germination of two Moroccan endemic thymes: Thymus maroccanus Ball. and Thymus broussonetii Boiss. Afr. J. Agric. Res. 2011, 6, 4740–4745. [Google Scholar] [CrossRef]
- Laghmouchi, Y.; Belmehdi, O.; Bouyahya, A.; Skali Senhaji, N.; Abrini, J. Effect of temperature, salt stress and pH on seed germination of medicinal plant Origanum compactum. Biocatal. Agric. Biotechnol. 2017, 10, 1–19. [Google Scholar] [CrossRef]
- Edwards, D.G.W. Breaking dormancy in tree seeds with special reference to firs (Abies species)—The 1.4x solution. In Proceedings of the ISTA Forest Tree and Shrub Seed Committee Workshop Prague—Pruhonice, Czech Republic; Prochazkova, Z., Gosling, P.G., Sutherland, J.R., Eds.; Forestry and Game Management Research Institute Jiloviste—Strnady, CR and Forestry Commission Research Agency: Průhonice, Prague, Czech Republic, 2003; pp. 18–23. [Google Scholar]
- Edwards, D.G.W. Pinaceae—Pine family: Abies P. Mill. fir. In Woody Plant Seed Manual. Agriculture Handbook No. 727; Bonner, F.T., Karrfalt, R.P., Eds.; USDA Forest Service: Washington, DC, USA, 2008; pp. 149–198. [Google Scholar]
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Anestis, I.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Greveniotis, V.; Tsiripidis, I.; et al. Agro-Alimentary Potential of the Neglected and Underutilized Local Endemic Plants of Crete (Greece), Rif-Mediterranean Coast of Morocco and Tunisia: Perspectives and Challenges. Plants 2021, 10, 1770. [Google Scholar] [CrossRef]
- Bourgou, S.; Jilani, I.B.H.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Khabbach, A.; Libiad, M.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; et al. Medicinal-cosmetic potential of the local endemic plants of Crete (Greece), Rif-Mediterranean Coast of Morocco and Tunisia: Priorities for sustainable exploitation. Biology 2021. under review. [Google Scholar]
- Krigas, N.; Mouflis, G.; Grigoriadou, K. Conservation of important plants from the Ionian Islands at the Balkan Botanic Garden of Kroussia, N Greece: Using GIS to link the in situ collection data with plant propagation and ex situ cultivation. Biodivers Conserv. 2010, 19, 3583–3603. [Google Scholar] [CrossRef]
- Krigas, N.; Papadimitriou, K.; Mazaris, A.D. GIS and ex-situ plant conservation. In Application of Geographic Information Systems; Alam, B.M., Ed.; InTechopen.com: Rijeka, Croatia, 2012. [Google Scholar]
- Padulosi, S.; Thompson, J.; Rudebjer, P. Fighting Poverty, Hunger and Malnutrition with Neglected and Underutilized Species (NUS): Needs, Challenges and the Way Forward; Bioversity International: Rome, Italy, 2013; p. 56. ISBN 978-92-9043-941-7. [Google Scholar]
- Yang, X.W.; Li, S.M.; Shen, Y.H.; Zhang, W.D. Phytochemical and biological studies of Abies species. Chem. Biodivers. 2008, 5, 56–81. [Google Scholar] [CrossRef]
- Wajs, A.; Urbańska, J.; Zaleśkiewicz, E.; Bonikowski, R. Composition of essential oil from seeds and cones of Abies alba. Nat. Prod. Commun. 2010, 5, 1291–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloukina, C.; Tomou, E.M.; Krigas, N.; Sarropoulou, V.; Madesis, P.; Maloupa, E.; Skaltsa, H. Non-polar secondary metabolites and essential oil of ex situ propagated and cultivated Sideritis syriaca L. subsp. syriaca (Lamiaceae) with consolidated identity (DNA Barcoding): Towards a potential new industrial crop. Ind. Crop. Prod. 2020, 158, 112957. [Google Scholar] [CrossRef]
- Macdonald, B. Practical Woody Plant Propagation for Nursery Growers; Timber Press Inc.: Portland, Oregon, 2006; p. 669. [Google Scholar]
- Stussi, I.; Henry, F.; Moser, P.; Danoux, L.; Jeanmaire, C.; Gillon, V.; Benoit, I.; Charrouf, Z.; Pauly, G. Argania spinosa—How ecological farming, fair trade and sustainability can drive the research for new cosmetic active ingredients. SÖFW-J. 2005, 131, 35–46. Available online: http://www.arganoils.com/articles/en/How%20Ecological%20Farming,%20Fair%20Trade%20and%20Sustainability%20Can%20Drive%20research%20for%20new%20cosmetic%20ingredients.pdf (accessed on 4 August 2021).
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- ISTA (International Seed Testing Association). International Rules for Seed Testing; Seed Science and Technology: Wallisellen, Switzerland, 1999; p. 333. [Google Scholar]
- Edwards, D.G.W. The stratification-redry technique with special reference to true fir seeds. In National Proceedings, Forest and Conservation Nursery Associations; General Technical Report, PNW-GTR-389; Landis, T.D., South, D.B., Eds.; U.S.D.A. Forest Service, Pacific Northwest Research Station: Portland, Oregon, 1996; pp. 172–182. [Google Scholar]
- Politi, P.I.; Georghiou, K.; Arianoutsou, M. Reproductive biology of Abies cephalonica Loudon in Mount Aenos National Park, Cephalonia, Greece. Trees 2011, 25, 655–668. [Google Scholar] [CrossRef]
- Varsamis, G.K.; Takos, I.A.; Merou, T.P.; Galatsidas, S.A.; Dimopoulos, P.D. Germination characteristics of Abies × borisii-regis. Seed Technol. 2014, 36, 51–59. [Google Scholar]
- Skrzyszewska, K.; Chlanda, J. A study on the variation of morphological characteristics of silver fir (Abies alba Mill.) seeds and their internal structure determined by X-ray radiography in the Beskid Sadecki and Beskid Niski mountain ranges of the Carpathians (southern Poland). J. For. Sci. 2009, 55, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Rawat, B.S.; Khanduri, V.P.; Sharma, C.M. Beneficial effects of cold-moist stratification on seed germination behaviors of Abies pindrow and Picea smithiana. J. For. Res. 2008, 19, 125–130. [Google Scholar] [CrossRef]
- Leadem, C.L. Stratification of Abies amabilis seeds. Can. J. For. Res. 1986, 16, 755–760. [Google Scholar] [CrossRef]
- Daskalakou, E.N.; Koutsovoulou, K.; Mavroeidi, L.; Tsiamitas, C.; Kafali, E.; Radaiou, P.E.; Ganatsas, P.; Thanos, C.A. Interannual variability of germination and cone/seed morphometric characteristics in the endemic Grecian fir (Abies cephalonica) over an 8-year-long study. Seed Sci. Res. 2018, 28, 24–33. [Google Scholar] [CrossRef]
- Jones, S.K.; Samuel, Y.K.; Gosling, P.G. The effect of soaking and prechilling on the germination of noble fir seeds. Seed Sci. Technol. 1991, 19, 287–294. [Google Scholar]
- Blazich, F.A.; Hinesley, L.E. Low temperature germination of Fraser fir seed. Can. J. For. Res. 1984, 14, 948–949. [Google Scholar] [CrossRef]
- Adkins, C.R.; Hinesley, L.E.; Blazich, F.A. Role of stratification, temperature, and light in Fraser fir germination. Can. J. For. Res. 1984, 14, 88–93. [Google Scholar] [CrossRef]
- Pipinis, E.; Milios, E.; Kiamos, N.; Mavrokordopoulou, O.; Smiris, P. Effects of stratification and pre-treatment with gibberellic acid on seed germination of two Carpinus species. Seed Sci. Technol. 2012, 40, 21–31. [Google Scholar] [CrossRef]
- Pipinis, E.; Milios, E.; Mavrokordopoulou, O.; Smiris, P. Effect of sowing date on seedling emergence of species with seeds enclosed in a stony endocarp. J. Sustain. For. 2018, 37, 375–388. [Google Scholar] [CrossRef]
- Pipinis, E.; Stampoulidis, A.; Milios, E.; Kitikidou, K.; Akritidou, S.; Theodoridou, S.; Radoglou, K. Effects of seed moisture content, stratification and sowing date on the germination of Corylus avellana seeds. J. For. Res. 2020, 31, 743–749. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005; p. 250. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Br. 2019, 12, 662–666. [Google Scholar] [CrossRef]
- Ben-Said, M.; Ghallab, A.; Lamrhari, H.; Carreira, J.A.; Linares, J.C.; Taïqui, L. Characterizing spatial structure of Abies marocana forest through point pattern analysis. For. Syst. 2020, 29, e016. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Inter. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Plant Propagation: Principles and Practices, 6th ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1997; p. 770. [Google Scholar]
- Snedecor, G.W.; Cochran, W.C. Statistical Methods, 7th ed.; The Iowa State University Press: Ames, IA, USA, 1980; p. 507. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 7th ed.; John Wiley and Sons Publications Inc.: New York, NY, USA, 1984; p. 680. [Google Scholar]
- Klockars, A.; Sax, G. Multiple Comparisons; Sage Publications: Newbury Park, CA, USA, 1986; p. 87. [Google Scholar]
- Toothaker, L. Multiple Comparison Procedures; Sage Publications: Newbury Park, CA, USA, 1993; p. 96. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attributes | Factors | Sum of Squares | df | Mean Square | F | Sig. |
|---|---|---|---|---|---|---|
| Cold Stratification (CS) | 250.98 | 2 | 125.49 | 6.35 | 0.004 | |
| Germination Percentage (GP) | Temperature (T) | 367.82 | 3 | 122.60 | 6.20 | 0.002 |
| CS * T | 380.42 | 6 | 63.40 | 3.21 | 0.013 | |
| Cold Stratification (CS) | 14,770.65 | 2 | 7385.33 | 677.24 | 0.000 | |
| Mean Germination Time (MGT) | Temperature (T) | 2965.46 | 3 | 988.49 | 90.64 | 0.000 |
| CS * T | 563.41 | 6 | 93.90 | 8.61 | 0.000 |
| Cold Stratification(Months) | Incubation Temperature | |||
|---|---|---|---|---|
| 10 °C | 15 °C | 20 °C | 25 °C | |
| Germination percentage (% ± S.D.) | ||||
| 0 | 12.00 b 1 B 2 ± 5.66 | 35.00 a A ± 6.83 | 19.00 ab A ± 6.83 | 21.00 ab A ± 3.83 |
| 1 | 27.00 a A ± 6.83 | 32.00 a A ± 8.64 | 26.00 a A ± 6.93 | 23.00 a A ± 6.00 |
| 2 | 24.00 a AB ± 6.53 | 22.00 a A ± 5.16 | 16.00 a A ± 5.66 | 15.00 a A ± 6.83 |
| Mean Germination Time (days ± S.D.) | ||||
| 0 | 74.26 a 1 A 2 ± 4.50 | 50.15 b A ± 2.40 | 52.77 b A ± 3.70 | 48.18 b A ± 4.62 |
| 1 | 38.13 a B ± 5.99 | 17.72 b B ± 1.34 | 14.35 b B ± 1.01 | 14.75 b B ± 0.96 |
| 2 | 22.83 a C ± 3.55 | 18.35 ab B ± 2.48 | 15.46 b B ± 2.21 | 12.69 b B ± 2.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatzilazarou, S.; El Haissoufi, M.; Pipinis, E.; Kostas, S.; Libiad, M.; Khabbach, A.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; et al. GIS-Facilitated Seed Germination and Multifaceted Evaluation of the Endangered Abies marocana Trab. (Pinaceae) Enabling Conservation and Sustainable Exploitation. Plants 2021, 10, 2606. https://doi.org/10.3390/plants10122606
Hatzilazarou S, El Haissoufi M, Pipinis E, Kostas S, Libiad M, Khabbach A, Lamchouri F, Bourgou S, Megdiche-Ksouri W, Ghrabi-Gammar Z, et al. GIS-Facilitated Seed Germination and Multifaceted Evaluation of the Endangered Abies marocana Trab. (Pinaceae) Enabling Conservation and Sustainable Exploitation. Plants. 2021; 10(12):2606. https://doi.org/10.3390/plants10122606
Chicago/Turabian StyleHatzilazarou, Stefanos, Mohamed El Haissoufi, Elias Pipinis, Stefanos Kostas, Mohamed Libiad, Abdelmajid Khabbach, Fatima Lamchouri, Soumaya Bourgou, Wided Megdiche-Ksouri, Zeineb Ghrabi-Gammar, and et al. 2021. "GIS-Facilitated Seed Germination and Multifaceted Evaluation of the Endangered Abies marocana Trab. (Pinaceae) Enabling Conservation and Sustainable Exploitation" Plants 10, no. 12: 2606. https://doi.org/10.3390/plants10122606