
Beneficial Effect of Root or Foliar Silicon Applied to Cucumber Plants under Different Zinc Nutritional Statuses


Abstract
:1. Introduction
2. Results
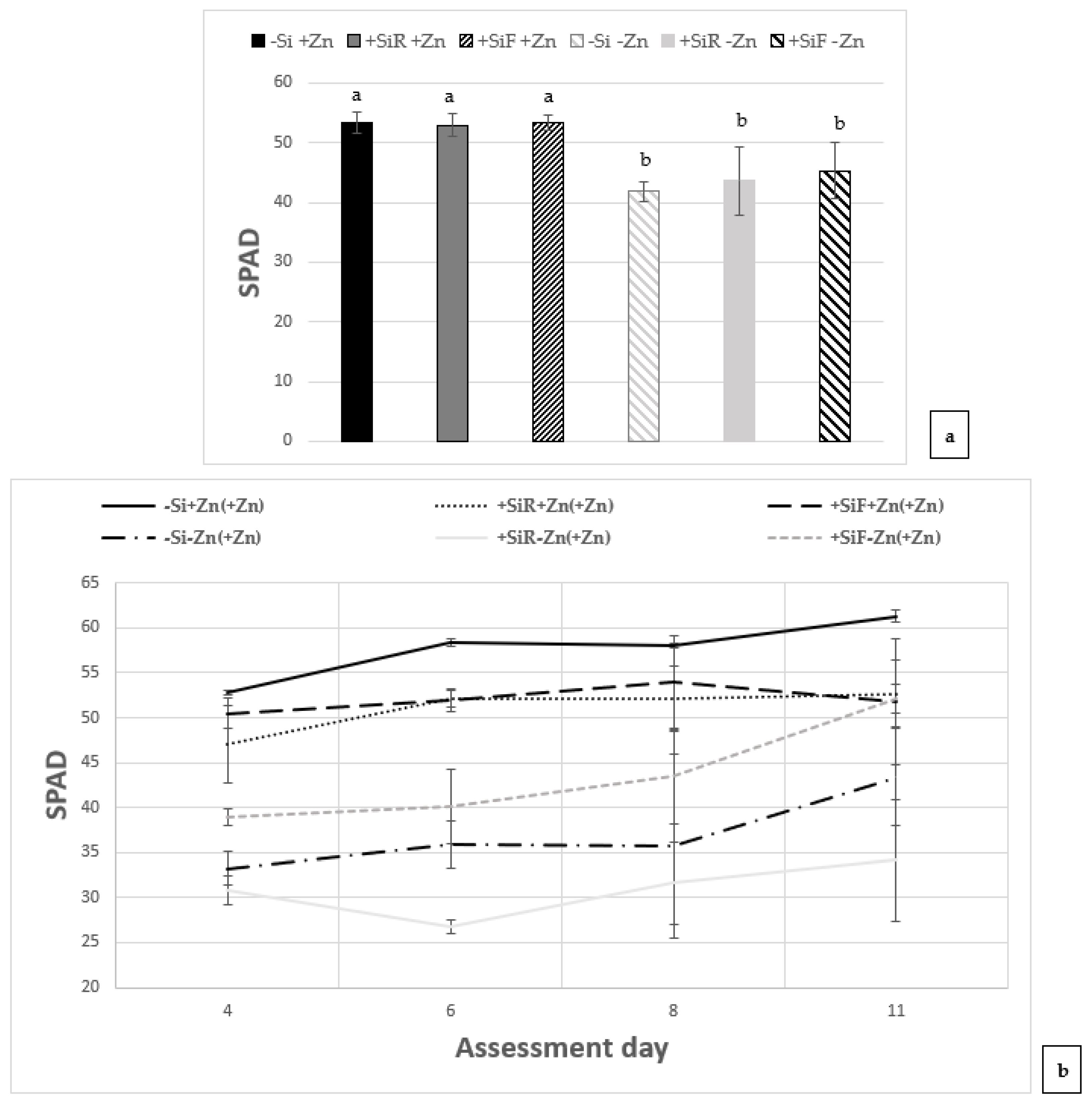
2.1. Effect of Silicon on SPAD Index and Biomass under Different Zn Nutrient Statuses
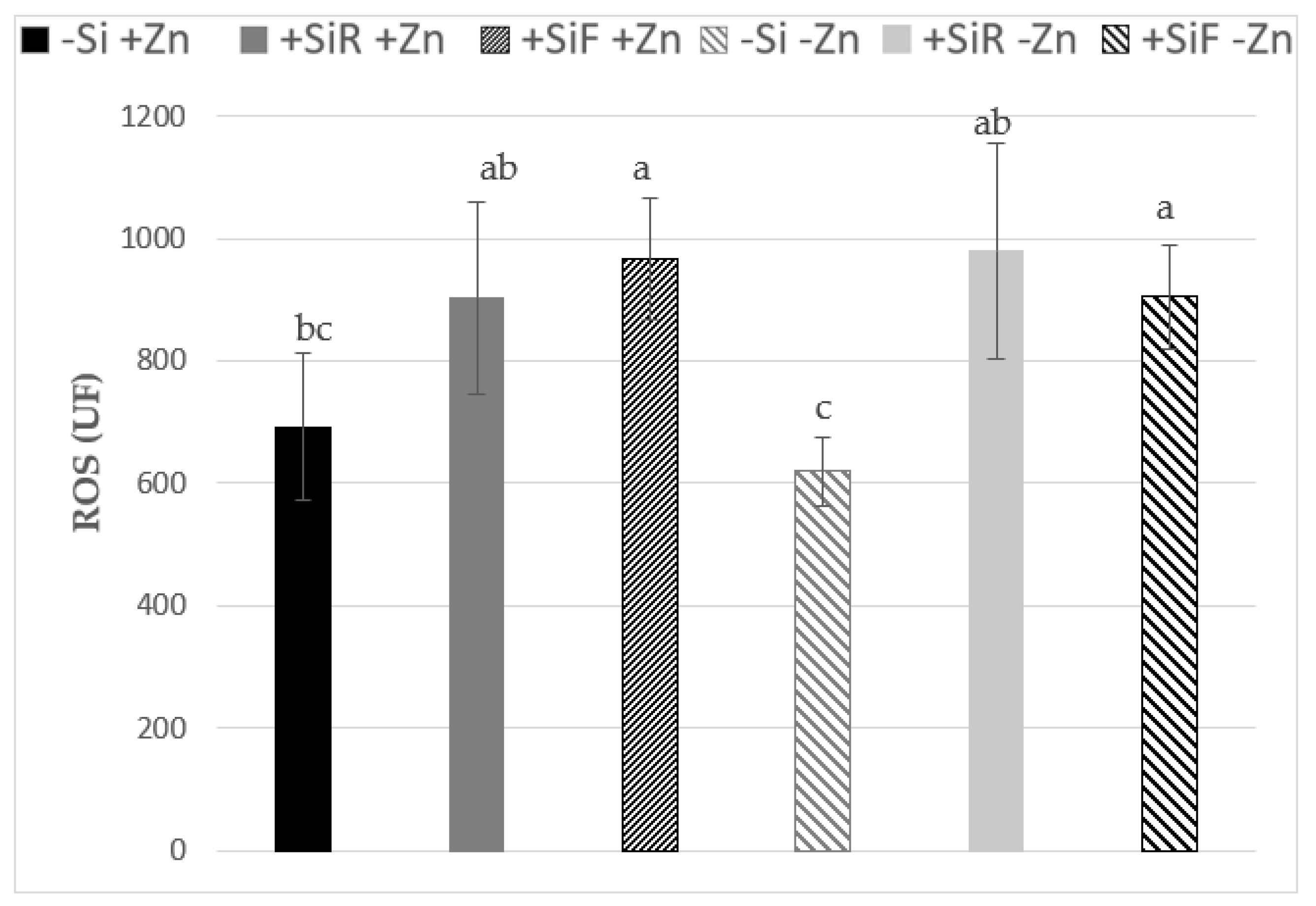
2.2. Effect of Silicon on ROS under Different Zn Nutrient Statuses
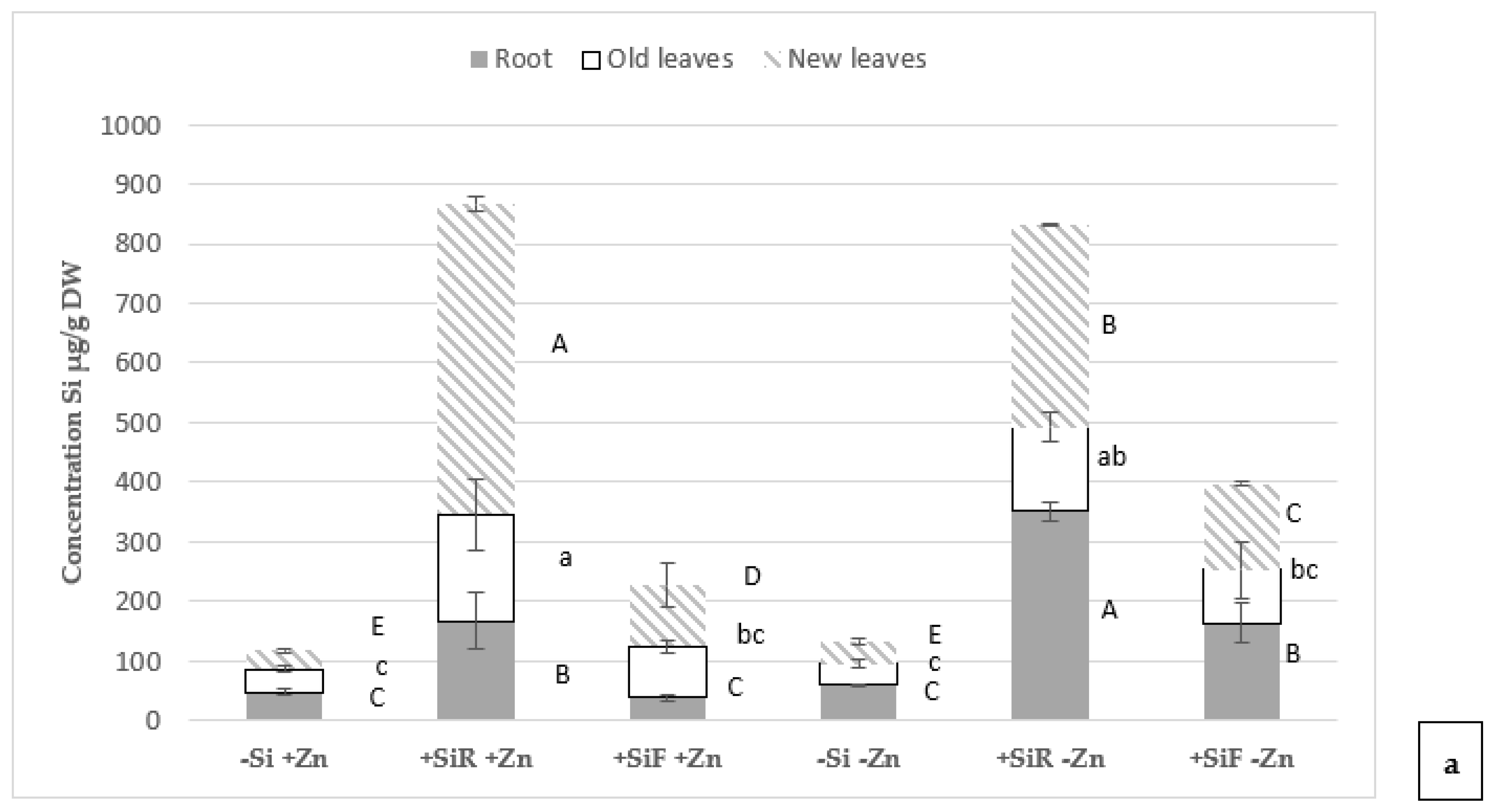
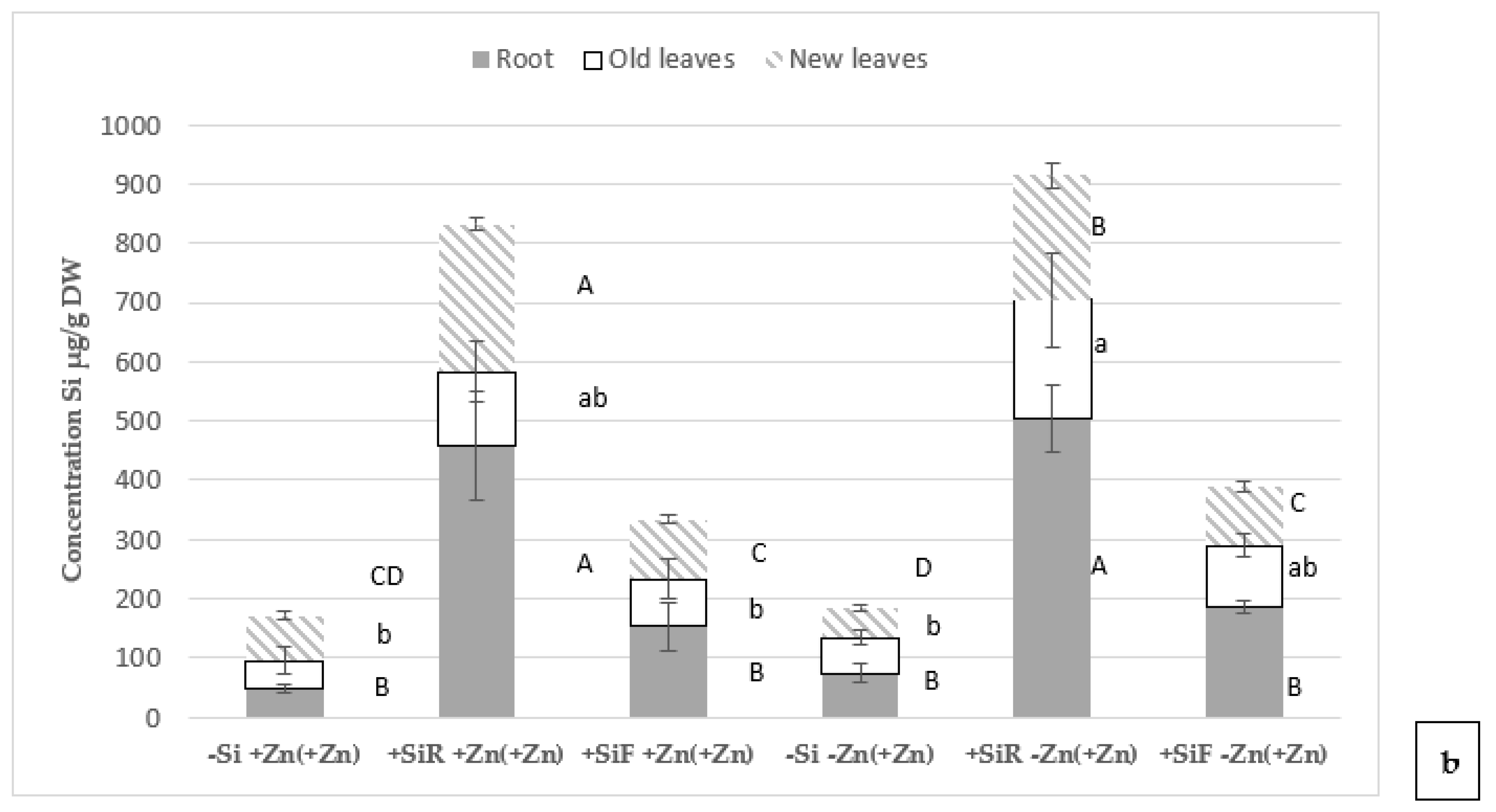
2.3. Effect of Silicon on Mineral Concentration in Plant Tissues under Different Zn Nutrient Statuses
3. Discussion
3.1. Effect of Silicon on SPAD Index and Biomass under Different Zn Nutrient Statuses
3.2. Effect of Silicon on ROS under Different Zn Nutritional Statuses
3.3. Effect of Silicon on Mineral Concentration in Plant Tissues under Different Zn Nutritional Statuses
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Determinations
4.3. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Hernandez-Apaolaza, L. Can Silicon Partially Alleviate Micronutrient Deficiency in Plants? A Review. Planta 2014, 240, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. The Anomaly of Silicon in Plant Biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellanos González, L.; de Mello Prado, R.; Silva Campos, C.N. El Silicio En La Resistencia de Los Cultivos. Cultiv. Trop. 2015, 36, 16–24. [Google Scholar]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The Controversies of Silicon’s Role in Plant Biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of Silicon with Essential and Beneficial Elements in Plants. Front. Plant Sci. 2021, 12, 1224. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Gil, S.; Rodríguez-Menéndez, S.; Fernández, B.; Pereiro, R.; de la Fuente, V.; Hernandez-Apaolaza, L. Silicon Induced Fe Deficiency Affects Fe, Mn, Cu and Zn Distribution in Rice (Oryza sativa L.) Growth in Calcareous Conditions. Plant Physiol. Biochem. 2018, 125, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and Future Prospects on the Action Mechanisms in Alleviating Biotic and Abiotic Stresses in Plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Ma, J.F. Role of Silicon in Enhancing the Resistance of Plants to Biotic and Abiotic Stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Pavlovic, J.; Samardzic, J.; Kostic, L.; Laursen, K.H.; Natic, M.; Timotijevic, G.; Schjoerring, J.K.; Nikolic, M. Silicon Enhances Leaf Remobilization of Iron in Cucumber under Limited Iron Conditions. Ann. Bot. 2016, 118, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Peris-Felipo, F.J.; Benavent-Gil, Y.; Hernández-Apaolaza, L. Silicon Beneficial Effects on Yield, Fruit Quality and Shelf-Life of Strawberries Grown in Different Culture Substrates under Different Iron Status. Plant Physiol. Biochem. 2020, 152, 23–31. [Google Scholar] [CrossRef]
- Bityutskii, N.; Pavlovic, J.; Yakkonen, K.; Maksimović, V.; Nikolic, M. Contrasting Effect of Silicon on Iron, Zinc and Manganese Status and Accumulation of Metal-Mobilizing Compounds in Micronutrient-Deficient Cucumber. Plant Physiol. Biochem. 2014, 74, 205–211. [Google Scholar] [CrossRef]
- Martín-Esquinas, A.; Hernández-Apaolaza, L. Rice Responses to Silicon Addition at Different Fe Status and Growth PH. Evaluation of Ploidy Changes. Plant Physiol. Biochem. 2021, 163, 296–307. [Google Scholar] [CrossRef]
- Pavlovic, J.; Samardzic, J.; Maksimović, V.; Timotijevic, G.; Stevic, N.; Laursen, K.H.; Hansen, T.H.; Husted, S.; Schjoerring, J.K.; Liang, Y.; et al. Silicon Alleviates Iron Deficiency in Cucumber by Promoting Mobilization of Iron in the Root Apoplast. New Phytol. 2013, 198, 1096–1107. [Google Scholar] [CrossRef]
- Ahmad, A.; Afzal, M.; Ahmad, A.; Tahir, M. Effect of Foliar Application of Silicon on Yield and Quality of Rice (Oryza sativa L.). Cercet. Agron. Mold. 2013, 46, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Apaolaza, L.; Escribano, L.; Zamarreño, Á.M.; García-Mina, J.M.; Cano, C.; Carrasco-Gil, S. Root Silicon Addition Induces Fe Deficiency in Cucumber Plants, but Facilitates Their Recovery After Fe Resupply. A Comparison with Si Foliar Sprays. Front. Plant Sci. 2020, 11, 1851. [Google Scholar] [CrossRef]
- Hussain, S.; Mumtaz, M.; Manzoor, S.; Shuxian, L.; Ahmed, I.; Skalicky, M.; Brestic, M.; Rastogi, A.; Ulhassan, Z.; Shafiq, I.; et al. Foliar Application of Silicon Improves Growth of Soybean by Enhancing Carbon Metabolism under Shading Conditions. Plant Physiol. Biochem. 2021, 159, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Rattan, R.K.; Goswami, N.N. Mineral Nutrition of Plants. Soil Science: An Introduction; Indian Society of Soil Science: New Delhi, India, 2015; pp. 499–539. [Google Scholar]
- Alloway, B.J. Zinc in Soils and Crop Nutrition; International Zinc Association Communications: Brussels, Belgium, 2004; p. 116. [Google Scholar]
- Bettger, W.J.; O’Dell, B.L. A Critical Physiological Role of Zinc in the Structure and Function of Biomembranes. Life Sci. 1981, 28, 1425–1438. [Google Scholar] [CrossRef]
- Malakouti, M.J. The Effect of Micronutrients in Ensuring Efficient Use of Macronutrients. Turk. J. Agric. For. 2008, 32, 215–220. [Google Scholar]
- Karthika, K.S.; Rashmi, I.; Parvathi, M.S. Biological Functions, Uptake and Transport of Essential Nutrients in Relation to Plant Growth. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 1–49. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of Oxidative Stress in Plants: From Classical Chemistry to Cell Biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Webb, M.J.; Loneragan, J.F. Effect of Zinc Deficiency on Growth, Phosphorus Concentration, and Phosphorus Toxicity of Wheat Plants. Soil Sci. Soc. Am. J. 1988, 52, 1676–1680. [Google Scholar] [CrossRef]
- Alloway, B.J. Micronutrients and Crop Production: An Introduction. In Micronutrients Deficiencies in Global Crop Production; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–39. [Google Scholar] [CrossRef]
- Mehrabanjoubani, P.; Abdolzadeh, A.; Sadeghipour, H.R.; Aghdasi, M. Impacts of Silicon Nutrition on Growth and Nutrient Status of Rice Plants Grown under Varying Zinc Regimes. Theor. Exp. Plant Physiol. 2015, 27, 19–29. [Google Scholar] [CrossRef]
- Pascual, M.B.; Echevarria, V.; Gonzalo, M.J.; Hernández-Apaolaza, L. Silicon Addition to Soybean (Glycine max L.) Plants Alleviate Zinc Deficiency. Plant Physiol. Biochem. 2016, 108, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Ngo, N.S.; Schenk, M.K.A. Silicon Reduces the Iron Uptake in Rice and Induces Iron Homeostasis Related Genes. Sci. Rep. 2020, 10, 5079. [Google Scholar] [CrossRef] [Green Version]
- Fleck, A.T.; Nye, T.; Repenning, C.; Stahl, F.; Zahn, M.; Schenk, M.K. Silicon Enhances Suberization and Lignification in Roots of Rice (Oryza sativa). J. Exp. Bot. 2011, 62, 2001–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinrichs, M.; Fleck, A.T.; Biedermann, E.; Ngo, N.S.; Schreiber, L.; Schenk, M.K. An ABC Transporter Is Involved in the Silicon-Induced Formation of Casparian Bands in the Exodermis of Rice. Front. Plant Sci. 2017, 8, 671. [Google Scholar] [CrossRef]
- Kreszies, T.; Kreszies, V.; Ly, F.; Thangamani, P.D.; Shellakkutti, N.; Schreiber, L. Suberized Transport Barriers in Plant Roots: The Effect of Silicon. J. Exp. Bot. 2020, 71, 6799–6806. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Peterson, C.A. Current Insights into the Development, Structure, and Chemistry of the Endodermis and Exodermis of Roots. Can. J. Bot. 2003, 81, 405–421. [Google Scholar] [CrossRef]
- Li, B.; Kamiya, T.; Kalmbach, L.; Yamagami, M.; Yamaguchi, K.; Shigenobu, S.; Sawa, S.; Danku, J.M.C.; Salt, D.E.; Geldner, N.; et al. Role of LOTR1 in Nutrient Transport through Organization of Spatial Distribution of Root Endodermal Barriers. Curr. Biol. 2017, 27, 758–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Calvo-Polanco, M.; Reyt, G.; Barberon, M.; Champeyroux, C.; Santoni, V.; Maurel, C.; Franke, R.B.; Ljung, K.; Novak, O.; et al. Surveillance of Cell Wall Diffusion Barrier Integrity Modulates Water and Solute Transport in Plants. Sci. Rep. 2019, 9, 4227. [Google Scholar] [CrossRef] [Green Version]
- de Farias Guedes, V.H.; de Mello Prado, R.; Frazão, J.J.; Oliveira, K.S.; Cazetta, J.O. Foliar-Applied Silicon in Sorghum (Sorghum bicolor L.) Alleviate Zinc Deficiency. Silicon 2020. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Suresh Kumar, J.; Suprasanna, P. Seed ‘Primeomics’: Plants Memorize Their Germination under Stress. Biol. Rev. 2021, 96, 1723–1743. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple Exposures to Drought “train” Transcriptional Responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef]
- Mehrabanjoubani, P.; Abdolzadeh, A.; Sadeghipour, H.R.; Aghdasi, M. Silicon Affects Transcellular and Apoplastic Uptake of Some Nutrients in Plants. Pedosphere 2015, 25, 192–201. [Google Scholar] [CrossRef]
- World Vegetable Center. The Possibility of Silicon as an Essential Element for Higher Plants. Available online: https://worldveg.tind.io/record/19607/ (accessed on 18 October 2021).
- Ma, J.F.; Miyake, Y.; Takahashi, E. Chapter 2 Silicon as a Beneficial Element for Crop Plants. In Studies in Plant Science; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Silicon in Agriculture Series; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 17–39. [Google Scholar] [CrossRef]
- Miyake, Y.; Takahashi, E. Effect of Silicon on the Growth of Solution-Cultured Cucumber Plant. Soil Sci. Plant Nutr. 1983, 29, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and Functional Characterization of Silicon Transporters in Soybean Using Comparative Genomics of Major Intrinsic Proteins in Arabidopsis and Rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and Antioxidant Signalling in Plants: A Re-Evaluation of the Concept of Oxidative Stress in a Physiological Context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Martínez-Hernández, G.B.; del Rodríguez, S.C.; Cao, C.-M.; Cisneros-Zevallos, L. Plants as Biofactories: Physiological Role of Reactive Oxygen Species on the Accumulation of Phenolic Antioxidants in Carrot Tissue under Wounding and Hyperoxia Stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar] [CrossRef]
- Surjadinata, B.B.; Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Physiological Role of Reactive Oxygen Species, Ethylene, and Jasmonic Acid on UV Light Induced Phenolic Biosynthesis in Wounded Carrot Tissue. Postharvest Biol. Technol. 2021, 172, 111388. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon Nanoparticles More Effectively Alleviated UV-B Stress than Silicon in Wheat (Triticum aestivum) Seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and Plants: Current Knowledge and Technological Perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, S.; Ali, S.; Ahmad, R.; Ercisli, S.; Anjum, M.A. Foliar Application of Silicon Enhances Growth, Flower Yield, Quality and Postharvest Life of Tuberose (Polianthes tuberosa L.) under Saline Conditions by Improving Antioxidant Defense Mechanism. Silicon 2021. [Google Scholar] [CrossRef]
- El-Hady, N.A.A.A.; ElSayed, A.I.; El-saadany, S.S.; Deligios, P.A.; Ledda, L. Exogenous Application of Foliar Salicylic Acid and Propolis Enhances Antioxidant Defenses and Growth Parameters in Tomato Plants. Plants 2021, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, H.; Esmaiel Pour, B.; Rizwan, M. Foliar Application of Silicon Nanoparticles Affected the Growth, Vitamin C, Flavonoid, and Antioxidant Enzyme Activities of Coriander (Coriandrum sativum L.) Plants Grown in Lead (Pb)-Spiked Soil. Environ. Sci. Pollut. Res. 2021, 28, 1417–1425. [Google Scholar] [CrossRef]
- Hacisalihoglu, G.; Hart, J.J.; Vallejos, C.E.; Kochian, L.V. The role of shoot-localized processes in the mechanism of Zn efficiency in common bean. Planta 2004, 218, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Bosnić, D.; Bosnić, P.; Nikolić, D.; Nikolić, M.; Samardžić, J. Silicon and Iron Differently Alleviate Copper Toxicity in Cucumber Leaves. Plants 2019, 8, 554. [Google Scholar] [CrossRef] [Green Version]
- Pilon, M.; Ravet, K.; Tapken, W. The Biogenesis and Physiological Function of Chloroplast Superoxide Dismutases. Biochim. Biophys. Acta BBA Bioenerg. 2011, 1807, 989–998. [Google Scholar] [CrossRef] [Green Version]
- Bosnić, D.; Nikolić, D.; Timotijević, G.; Pavlović, J.; Vaculík, M.; Samardžić, J.; Nikolić, M. Silicon Alleviates Copper (Cu) Toxicity in Cucumber by Increased Cu-Binding Capacity. Plant Soil 2019, 441, 629–641. [Google Scholar] [CrossRef]
- Sun, B.; Jing, Y.; Chen, K.; Song, L.; Chen, F.; Zhang, L. Protective Effect of Nitric Oxide on Iron Deficiency-Induced Oxidative Stress in Maize (Zea mays). J. Plant Physiol. 2007, 164, 536–543. [Google Scholar] [CrossRef]
- M’sehli, W.; Houmani, H.; Donnini, S.; Zocchi, G.; Abdelly, C.; Gharsalli, M. Iron Deficiency Tolerance at Leaf Level in Medicago ciliaris Plants. Am. J. Plant Sci. 2014, 05, 2541–2553. [Google Scholar] [CrossRef] [Green Version]
- Pilon, C.; Soratto, R.P.; Moreno, L.A. Effects of Soil and Foliar Application of Soluble Silicon on Mineral Nutrition, Gas Exchange, and Growth of Potato Plants. Crop Sci. 2013, 53, 1605–1614. [Google Scholar] [CrossRef]
- Hacisalihoglu, G. Zinc (Zn): The Last Nutrient in the Alphabet and Shedding Light on Zn Eficiency for the Future of Crop Production under Suboptimal Zn. Plants 2020, 9, 1471. [Google Scholar] [CrossRef] [PubMed]
- Kostic, L.; Nikolic, N.; Bosnic, D.; Samardzic, J.; Nikolic, M. Silicon Increases Phosphorus (P) Uptake by Wheat under Low P Acid Soil Conditions. Plant Soil 2017, 419, 447–455. [Google Scholar] [CrossRef]
- Soratto, R.P.; Fernandes, A.M.; Pilon, C.; Souza, M.R. Phosphorus and Silicon Effects on Growth, Yield, and Phosphorus Forms in Potato Plants. J. Plant Nutr. 2019, 42, 218–233. [Google Scholar] [CrossRef]
- Wang, Y.; Krogstad, T.; Clarke, J.L.; Hallama, M.; Øgaard, A.F.; Eich-Greatorex, S.; Kandeler, E.; Clarke, N. Rhizosphere Organic Anions Play a Minor Role in Improving Crop Species’ Ability to Take Up Residual Phosphorus (P) in Agricultural Soils Low in P Availability. Front. Plant Sci. 2016, 7, 1664. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Marschner, H. Mechanism of Phosphorus-Induced Zinc Deficiency in Cotton. I. Zinc Deficiency-Enhanced Uptake Rate of Phosphorus. Physiol. Plant. 1986, 68, 483–490. [Google Scholar] [CrossRef]
- Keller, C.; Rizwan, M.; Davidian, J.-C.; Pokrovsky, O.S.; Bovet, N.; Chaurand, P.; Meunier, J.-D. Effect of Silicon on Wheat Seedlings (Triticum turgidum L.) Grown in Hydroponics and Exposed to 0 to 30 ΜM Cu. Planta 2015, 241, 847–860. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Shen, Z. Interaction of Silicon and Boron in Oilseed Rape Plants. J. Plant Nutr. 1994, 17, 415–425. [Google Scholar] [CrossRef]
- Celikkol Akcay, U.; Erkan, I. Silicon Induced Antioxidative Responses and Expression of BOR2 and Two PIP Family Aquaporin Genes in Barley Grown Under Boron Toxicity. Plant Mol. Biol. Rep. 2016, 34, 318–326. [Google Scholar] [CrossRef]
- Gunes, A.; Inal, A.; Bagci, E.G.; Pilbeam, D.J. Silicon-Mediated Changes of Some Physiological and Enzymatic Parameters Symptomatic for Oxidative Stress in Spinach and Tomato Grown in Sodic-B Toxic Soil. Plant Soil 2007, 290, 103–114. [Google Scholar] [CrossRef]
- Soylemezoglu, G.; Demir, K.; Inal, A.; Gunes, A. Effect of Silicon on Antioxidant and Stomatal Response of Two Grapevine (Vitis vinifera L.) Rootstocks Grown in Boron Toxic, Saline and Boron Toxic-Saline Soil. Sci. Hortic. 2009, 123, 240–246. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Guneri, M.; Ashraf, M. Mitigation Effects of Silicon on Tomato Plants Bearing Fruit Grown at High Boron Levels. J. Plant Nutr. 2011, 34, 1985–1994. [Google Scholar] [CrossRef]
- Nozawa, S.; Sato, T.; Otake, T. Effect of Dissolved Silica on Immobilization of Boron by Magnesium Oxide. Minerals 2018, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Rogalla, H.; Römheld, V. Effects of Silicon on the Availability of Boron. In Boron in Plant and Animal Nutrition; Goldbach, H.E., Brown, P.H., Rerkasem, B., Thellier, M., Wimmer, M.A., Bell, R.W., Eds.; Springer: Boston, MA, USA, 2002; pp. 205–211. [Google Scholar] [CrossRef]
- Loomis, W.D.; Durst, R.W. Chemistry and Biology of Boron. BioFactors 1992, 3, 229–239. [Google Scholar]
- Camacho-Cristóbal, J.J.; Martín-Rejano, E.M.; Herrera-Rodríguez, M.B.; Navarro-Gochicoa, M.T.; Rexach, J.; González-Fontes, A. Boron Deficiency Inhibits Root Cell Elongation via an Ethylene/Auxin/ROS-Dependent Pathway in Arabidopsis Seedlings. J. Exp. Bot. 2015, 66, 3831–3840. [Google Scholar] [CrossRef]
- Gonzalo, M.J.; Lucena, J.J.; Hernández-Apaolaza, L. Effect of Silicon Addition on Soybean (Glycine max) and Cucumber (Cucumis sativus) Plants Grown under Iron Deficiency. Plant Physiol. Biochem. 2013, 70, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, M.; Nikolic, N.; Liang, Y.; Kirkby, E.A.; Römheld, V. Germanium-68 as an Adequate Tracer for Silicon Transport in Plants. Characterization of Silicon Uptake in Different Crop Species. Plant Physiol. 2007, 143, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Sparks, D.L. Methods of Soil Analysis Part 3: Chemical Methods; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fresh Weight Distribution in Plant Tissues (%) | Total FW (g) | ||||
|---|---|---|---|---|---|
| Root | Stem | Old Leaves | New Leaves | ||
| Sampling 1 | |||||
| −Si+Zn | 47.40 b | 9.52 ab | 31.23 a | 11.85 ab | 11.56 a |
| +SiR+Zn | 50.94 b | 8.32 b | 33.58 a | 7.17 b | 9.62 a |
| +SiF+Zn | 55.26 a | 7.33 ab | 25.44 a | 11.97 a | 15.96 a |
| −Si−Zn | 45.62 c | 15.67 c | 11.06 b | 27.65 b | 2.17 b |
| +SiR−Zn | 43.70 c | 14.96 c | 4.72 b | 36.61 b | 2.54 b |
| +SiF−Zn | 38.24 c | 15.69 c | 12.25 b | 33.82 b | 2.04 b |
| Sampling 2 | |||||
| −Si+Zn(+Zn) | 35.62 c | 25.12 b | 14.81 b | 24.46 c | 31.81 c |
| +SiR+Zn(+Zn) | 51.45 b | 19.51 a | 12.01 a | 17.04 d | 72.95 b |
| +SiF+Zn(+Zn) | 57.48 a | 15.25 a | 11.28 a | 15.98 d | 96.1 a |
| −Si−Zn(+Zn) | 37.15 c | 22.55 c | 7.95 c | 32.35 b | 5.41 d |
| +SiR−Zn(+Zn) | 41.90 c | 16.50 c | 7.83 c | 33.76 b | 13.15 d |
| +SiF−Zn(+Zn) | 34.74 c | 18.36 c | 4.49 c | 42.40 a | 7.57 d |
| ∆ROS (FU) | |
|---|---|
| Zn Sufficiency (Sampling 2-Sampling 1) | |
| −Si | −182.47 ± 44.06 e |
| +SiR | −223.23 ± 39.52 e |
| +SiF | −220.93 ± 26.77 e |
| Zn Resupply (Sampling 2)—Zn Deficiency (Sampling 1) | |
| −Si | 202.68 ± 13.42 a |
| +SiR | 114.54 ± 21.77 b |
| +SiF | −43.33 ± 8.45 c |
| Zn Distribution in Plant Tissues (%) | P Distribution in Plant Tissues (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Root | Old Leaves | New Leaves | Total (µg/g) | Root | Old Leaves | New Leaves | Total (mg/g) | |
| Sampling 1 | ||||||||
| −Si+Zn | 51.59 a | 21.28 n.s | 27.14 bc | 117.78 a | 33.49 cd | 20.36 c | 46.14 b | 4.18 b |
| +SiR+Zn | 45.97 bc | 18.13 | 35.90 abc | 94.48 ab | 32.381 b | 48.56 ab | 19.05 c | 6.75 ab |
| +SiF+Zn | 44.40 ab | 15.02 | 40.58 a | 118.57 a | 25.53 bcd | 31.99 bc | 42.46 c | 5.61 ab |
| −Si−Zn | 40.49 c | 29.50 | 30.00 c | 78.57 b | 34.60 d | 51.40 a | 13.99 c | 5.89 ab |
| +SiR−Zn | 42.65 c | 24.79 | 32.56 c | 80.37 b | 36.43 a | 47.75 abc | 15.82 c | 6.18 ab |
| +SiF−Zn | 40.06 bc | 20.67 | 39.28 ab | 110.75 a | 26.09 bc | 21.10 bc | 52.80 a | 9.45 a |
| Sampling 2 | ||||||||
| −Si+Zn(+Zn) | 26.02 b | 22.30 ab | 51.68 a | 145.64 b | 14.36 c | 64.07 c | 21.55 b | 1.96 b |
| +SiR+Zn(+Zn) | 47.30 a | 15.43 b | 37.27 a | 170.64 ab | 59.82 ab | 15.87 c | 24.30 b | 7.91 a |
| +SiF+Zn(+Zn) | 37.33 a | 23.51 a | 39.16 a | 183.89 a | 23.27 c | 50.79 bc | 25.93 b | 3.57 b |
| −Si−Zn(+Zn) | 33.45 c | 25.77 b | 40.77 b | 93.20 c | 36.03 bc | 48.68 a | 15.27 b | 7.71 a |
| +SiR−Zn(+Zn) | 46.78 a | 15.73 b | 37.49 a | 164.31 ab | 41.42 a | 43.99 ab | 14.57 b | 6.70 a |
| +SiF−Zn(+Zn) | 34.80 b | 18.01 b | 47.19 a | 128.68 bc | 22.46 c | 29.96 bc | 47.57 a | 8.00 a |
| Cu Distribution in Plant Tissues (%) | B Distribution in Plant Tissues (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Root | Old Leaves | New Leaves | Total (µg/g) | Root | Old Leaves | New Leaves | Total (µg/g) | |
| Sampling 1 | ||||||||
| −Si+Zn | 29.12 bc | 27.42 a | 43.46 b | 84.31 c | 17.40 b | 68.22 a | 14.39 c | 49.51 ab |
| +SiR+Zn | 28.90 bc | 11.83 c | 59.26 a | 152.56 b | 32.64 a | 64.50 a | 2.86 d | 55.47 a |
| +SiF+Zn | 26.79 c | 14.91 c | 58.30 a | 77.85 c | 15.71 b | 62.41 a | 21.88 bc | 58.23 a |
| −Si−Zn | 35.16 b | 25.05 a | 39.79 c | 259.78 a | 10.82 b | 52.17 b | 37.01 a | 44.55 b |
| +SiR−Zn | 39.73 b | 12.93 c | 47.34 b | 168.67 b | 12.73 b | 51.61 b | 35.67 a | 43.72 b |
| +SiF−Zn | 59.50 a | 20.12 b | 20.38 d | 274.10 a | 12.77 b | 48.56 b | 38.66 a | 56.11 a |
| Sampling 2 | ||||||||
| −Si+Zn(+Zn) | 33.01 d | 22.13 ab | 44.86 a | 81.78 c | 32.64 b | 46.31 b | 21.05 b | 70.29 b |
| +SiR+Zn(+Zn) | 48.91 c | 24.75 a | 26.34 c | 76.02 c | 12.77 c | 68.20 a | 19.03 b | 94.34 a |
| +SiF+Zn(+Zn) | 49.79 c | 22.79 ab | 27.43 c | 62.06 d | 28.79 b | 37.43 c | 33.78 a | 103.19 a |
| −Si−Zn(+Zn) | 60.67 b | 7.28 c | 32.05 b | 170.52 a | 26.84 c | 70.03 a | 3.13 c | 44.73 c |
| +SiR−Zn(+Zn) | 56.80 b | 18.51 b | 24.70 c | 86.43 c | 26.85 c | 36.95 c | 36.20 a | 57.93 c |
| +SiF−Zn(+Zn) | 71.57 a | 27.23 a | 1.20 d | 116.25 b | 38.78 a | 40.25 bc | 20.97 b | 108.87 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano-González, J.M.; Valverde, C.; Hernández, C.D.; Martin-Esquinas, A.; Hernández-Apaolaza, L. Beneficial Effect of Root or Foliar Silicon Applied to Cucumber Plants under Different Zinc Nutritional Statuses. Plants 2021, 10, 2602. https://doi.org/10.3390/plants10122602
Lozano-González JM, Valverde C, Hernández CD, Martin-Esquinas A, Hernández-Apaolaza L. Beneficial Effect of Root or Foliar Silicon Applied to Cucumber Plants under Different Zinc Nutritional Statuses. Plants. 2021; 10(12):2602. https://doi.org/10.3390/plants10122602
Chicago/Turabian StyleLozano-González, José María, Clara Valverde, Carlos David Hernández, Alexandra Martin-Esquinas, and Lourdes Hernández-Apaolaza. 2021. "Beneficial Effect of Root or Foliar Silicon Applied to Cucumber Plants under Different Zinc Nutritional Statuses" Plants 10, no. 12: 2602. https://doi.org/10.3390/plants10122602