
Morphological and Ultrastructural Features of Formation of the Skin of Wheat (Triticum aestivum L.) Kernel

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
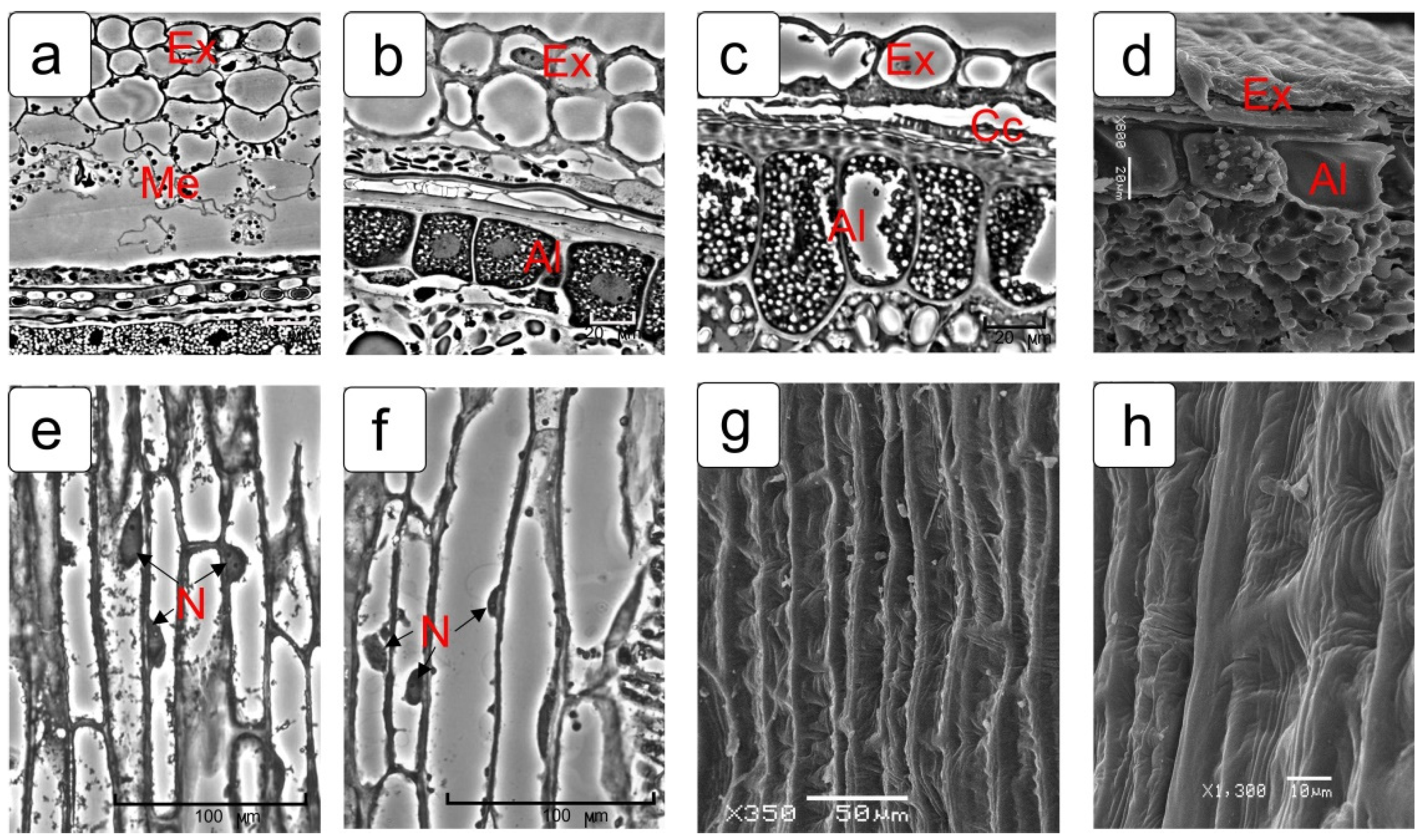
2.1. Pericarp
2.2. Cross Cells (Chlorophyll-Bearing Layer)
2.3. Endocarp (Tube Cells)
2.4. Integuments of Ovules
2.4.1. Outer Integument of Ovule
2.4.2. Inner Integument of Ovule
2.5. Epidermis of Nucellus
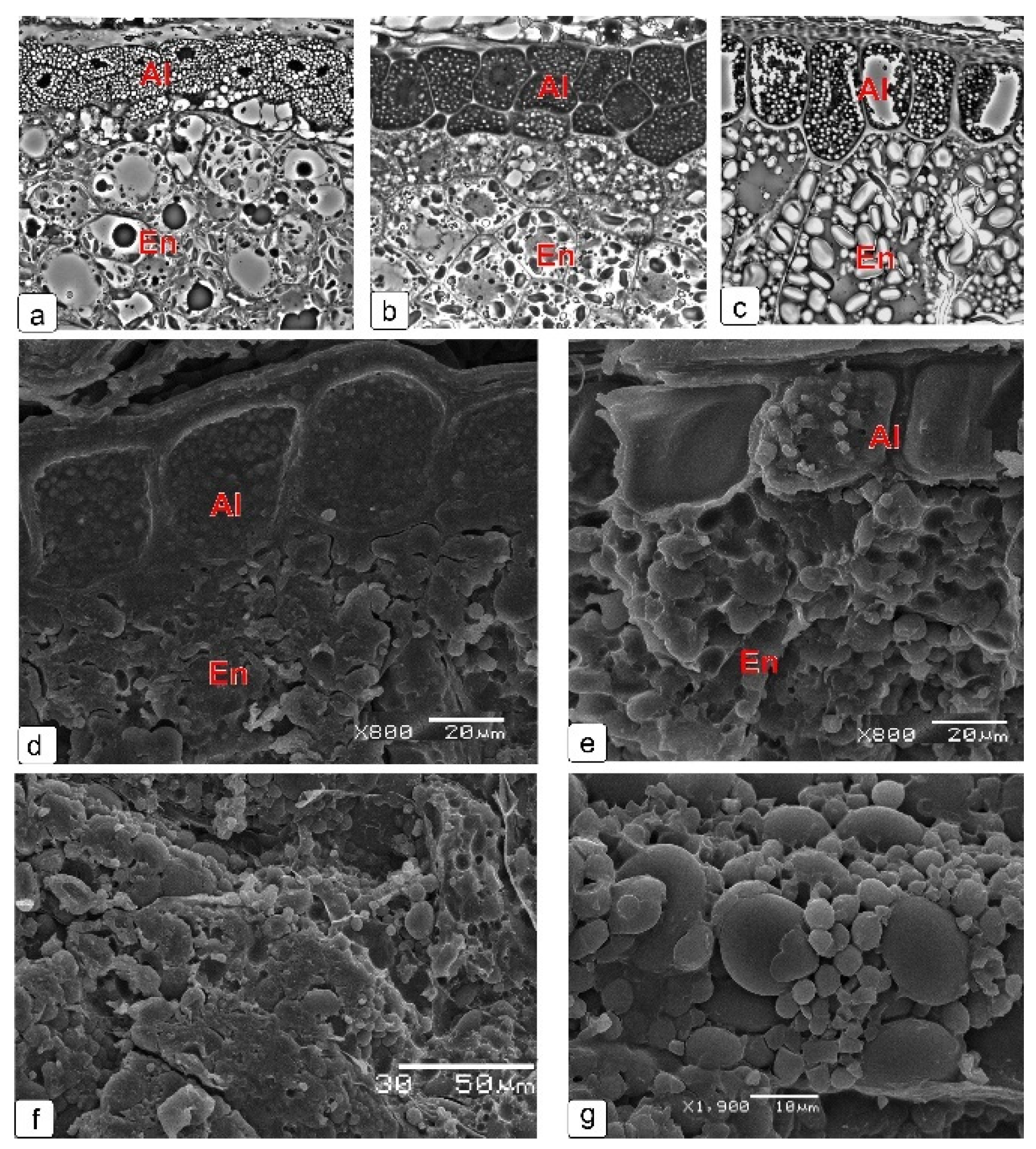
2.6. Endosperm
2.7. Cell Structure of All Layers of Kernel Skin at Final Stage of Development
3. Discussion
3.1. Pericarp
3.2. Outer Integument
3.3. Inner Integument
3.4. Nucellus
3.5. Epidermis of Nucellus
3.6. Endosperm
4. Materials and Methods
4.1. Plant Material
4.2. Light and Transmission Electron Microscopy
4.3. Scanning Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagel, M.; Börner, A. The longevity of crop seeds stored under ambient conditions. Seed Sci. Res. 2010, 20, 1. [Google Scholar] [CrossRef]
- Antoine, C.; Peyron, S.; Mabille, F.; Lapierre, C.; Bouchet, B.; Abecassis, J.; Rouau, X. Individual contribution of grain outer layers and their cell wall structure to the mechanical properties of wheat bran. J. Agric. Food Chem. 2003, 51, 2026–2033. [Google Scholar] [CrossRef] [PubMed]
- Baranova, E.N.; Levinskikh, M.A.; Gulevich, A.A. Wheat space odyssey: “From seed to seed”. Kernel morphology. Life 2019, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, F.; Yu, X.; Zhou, L.; Wang, F.; Xiong, A.S. Structural and physiological characterization during wheat pericarp development. Plant Cell Rep. 2013, 32, 1309–1320. [Google Scholar] [CrossRef]
- Yu, X.; Chen, X.; Zhou, L.; Zhang, J.; Yu, H.; Shao, S.; Xiong, F.; Wang, Z. Structural development of wheat nutrient transfer tissues and their relationships with filial tissues development. Protoplasma 2015, 252, 605–617. [Google Scholar]
- Earp, C.F.; McDonough, C.M.; Rooney, L.W. Microscopy of pericarp development in the caryopsis of Sorghum bicolor (L.). Moench. J. Cereal Sci. 2004, 39, 21–27. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, L.; Li, J.; Song, X.; Yang, C. Study on programmed cell death and dynamic changes of starch accumulation in pericarp cells of Triticum aestivum L. Protoplasma 2009, 236, 49–58. [Google Scholar] [CrossRef]
- Greenwood, J.S.; Helm, M.; Gietl, C. Ricinosomes and endosperm transfer cell structure in programmed cell death of the nucellus during Ricinus seed development. Proc. Natl. Acad. Sci. USA 2005, 102, 2238–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Gu, Y.J.; Zheng, Y.K.; Wang, H.H. Structure observation of rice endosperm cell development and its mineral element analysis. Chin. J. Rice Sci. 2012, 26, 693–705. [Google Scholar]
- Radchuk, V.; Weier, D.; Radchuk, R.; Weschke, W.; Weber, H. Development of maternal seed tissue in barley is mediated by regulated cell expansion and cell disintegration and coordinated with endosperm growth. J. Exp. Bot. 2011, 62, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Liu, J.; Li, D.; Liu, C.M. Rice caryopsis development I: Dynamic changes in different cell layers. J. Integr. Plant Biol. 2016, 58, 772–785. [Google Scholar] [CrossRef] [Green Version]
- Evers, T.; Millar, S. Cereal grain structure and development: Some implications for quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Krishnan, S.; Dayanandan, P. Structural and histochemical studies on grain-filling in the caryopsis of rice (Oryza sativa L.). J. Biosci. 2003, 28, 455–469. [Google Scholar] [CrossRef]
- Radchuk, V.; Borisjuk, L. Physical, metabolic and developmental functions of the seed coat. Front. Plant Sci. 2014, 5, 510. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.B.; Liu, J.X.; Li, D.Q.; Liu, C.M. Rice caryopsis development II: Dynamic changes in the endosperm. J. Integr. Plant Biol. 2016, 58, 786–798. [Google Scholar] [CrossRef]
- Ingram, G.C. Dying to live: Cell elimination as a developmental strategy in angiosperm seeds. J. Exp. Bot. 2017, 68, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez, F.; Cejudo, F.J. Programmed cell death (PCD): An essential process of cereal seed development and germination. Front. Plant Sci. 2014, 5, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daneva, A.; Gao, Z.; Van Durme, M.; Nowack, M.K. Functions and regulation of programmed cell death in plant development. Annu. Rev. Cell Dev. Biol. 2016, 32, 441–468. [Google Scholar] [CrossRef]
- Cubría-Radío, M.; Nowack, M.K. Transcriptional networks orchestrating programmed cell death during plant development. Curr. Top. Dev. Biol. 2019, 131, 161–184. [Google Scholar]
- Wang, Y.; Ye, H.; Bau, J.; Ren, F. The regulatory framework of developmentally programed cell death in floral organs: A review. Plant Physiol. Biochem. 2021, 158, 103–112. [Google Scholar] [CrossRef]
- Saulnier, L.; Guillon, F.; Chateigner-Boutin, A.L. Cell wall deposition and metabolism in wheat grain. J. Cereal Sci. 2012, 56, 91–108. [Google Scholar] [CrossRef]
- Tran, V.; Weier, D.; Radchuk, R.; Thiel, J.; Radchuk, V. Caspase-like activities accompany programmed cell death events in developing barley grains. PLoS ONE 2014, 10, e109426. [Google Scholar] [CrossRef]
- Płachno, B.; Kapusta, M.; Świątek, P.; Stolarczyk, P.; Kocki, J. Immunocytochemistry of the mucilage cells from the 3 ovules of Pilosella officinarum Vaill. and Taraxacum officinale Agg. (Asteraceae). Int. J. Mol. Sci. 2020, 21, 9642. [Google Scholar] [CrossRef]
- Radchuk, V.; Borisjuk, L.; Sreenivasulu, N.; Merx, K.; Mock, H.-P.; Rolletschek, H.; Wobus, U.; Weschke, W. Spatiotemporal profiling of starch biosynthesis and degradation in the development barley grain. Plant Physiol. 2009, 150, 190–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreenivasulu, N.; Borisjuk, L.; Junker, B.H.; Mock, H.P.; Rolletschek, H.; Seiffert, U.; Weschke, W.; Wobus, U. Barley grain development: Toward an integrative view. Int. Rev. Cell Mol. Biol. 2010, 281, 49–89. [Google Scholar] [PubMed]
- Yunjie, G.; Zhong, W.; Juan, C.; Guoyong, Z. The structure and function of pericarp in rice. Acta Agron. Sin. 2002, 28, 439–444. (In Chinese) [Google Scholar]
- Li, D.-L. Study of the development of Sorghum endosperm cells and the relationship with its maternal tissue. Sci. Sin. Agri. 2014, 47, 3336–3347. [Google Scholar]
- Dong, Y.; Huang, C.; Zhao, G. Research summary on pericarp thickness of maize. J. Maize Sci. 2016, 19, 146–147. (In Chinese) [Google Scholar]
- Van Lammeren, A.A.M.; Kieft, F.M.; Schel, J.H.N. Cell differentiation in the pericarp and endosperm of developing maize kernels. In Embryology of Flowering Plants, Terminology and Concepts. Volume 2: Seed; Batygina, T.B., Ed.; Science Publishers: Enfield, NH, USA, 2006; pp. 131–138. [Google Scholar]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Trivandrum: Kerala, India, 2006; pp. 23–67. [Google Scholar]
- Ruijtenberg, S.; van den Heuvel, S. Coordinating cell proliferation and differentiation: Antagonism between cell cycle regulators and cell type-specific gene expression. Cell Cycle 2016, 15, 196–212. [Google Scholar] [CrossRef] [Green Version]
- Velappan, Y.; Signorelli, S.; Considine, M.J. Cell cycle arrest in plants: What distinguishes quiescence, dormancy and differentiated G1? Ann. Bot. 2017, 120, 495–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Magnani, E. Seed tissue and nutrient partitioning, a case for the nucellus. Plant Reprod. 2018, 31, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Płachno, B.J.; Świątek, P.; Kozieradzka-Kiszkurno, M.; Szeląg, Z.; Stolarczyk, P. Integument cell gelatinization—The fate of the integumentary cells in Hieracium and Pilosella (Asteraceae). Protoplasma 2017, 254, 2287–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolczyk, J.; Stolarczyk, P.; Płachno, B. Comparative anatomy of ovules in Galinsoga, Solidago and Ratibida (Asteraceae). Acta Biol. Crac. Ser. Bot. 2014, 56, 115–125. [Google Scholar] [CrossRef]
- Kobayashi, H.; Ikeda, T.M.; Nagata, K. Spatial and temporal progress of programmed cell death in the developing starchy endosperm of rice. Planta 2013, 237, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Young, T.E.; Gallie, D.R. Programmed cell death during endosperm development. Plant Mol. Biol. 2000, 44, 283–301. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaban, I.A.; Gulevich, A.A.; Smirnova, E.A.; Baranova, E.N. Morphological and Ultrastructural Features of Formation of the Skin of Wheat (Triticum aestivum L.) Kernel. Plants 2021, 10, 2538. https://doi.org/10.3390/plants10112538
Chaban IA, Gulevich AA, Smirnova EA, Baranova EN. Morphological and Ultrastructural Features of Formation of the Skin of Wheat (Triticum aestivum L.) Kernel. Plants. 2021; 10(11):2538. https://doi.org/10.3390/plants10112538
Chicago/Turabian StyleChaban, Inna A., Alexander A. Gulevich, Elena A. Smirnova, and Ekaterina N. Baranova. 2021. "Morphological and Ultrastructural Features of Formation of the Skin of Wheat (Triticum aestivum L.) Kernel" Plants 10, no. 11: 2538. https://doi.org/10.3390/plants10112538