
Phylogeny of Maleae (Rosaceae) Based on Complete Chloroplast Genomes Supports the Distinction of Aria, Chamaemespilus and Torminalis as Separate Genera, Different from Sorbus sp.


Abstract
:1. Introduction
2. Results
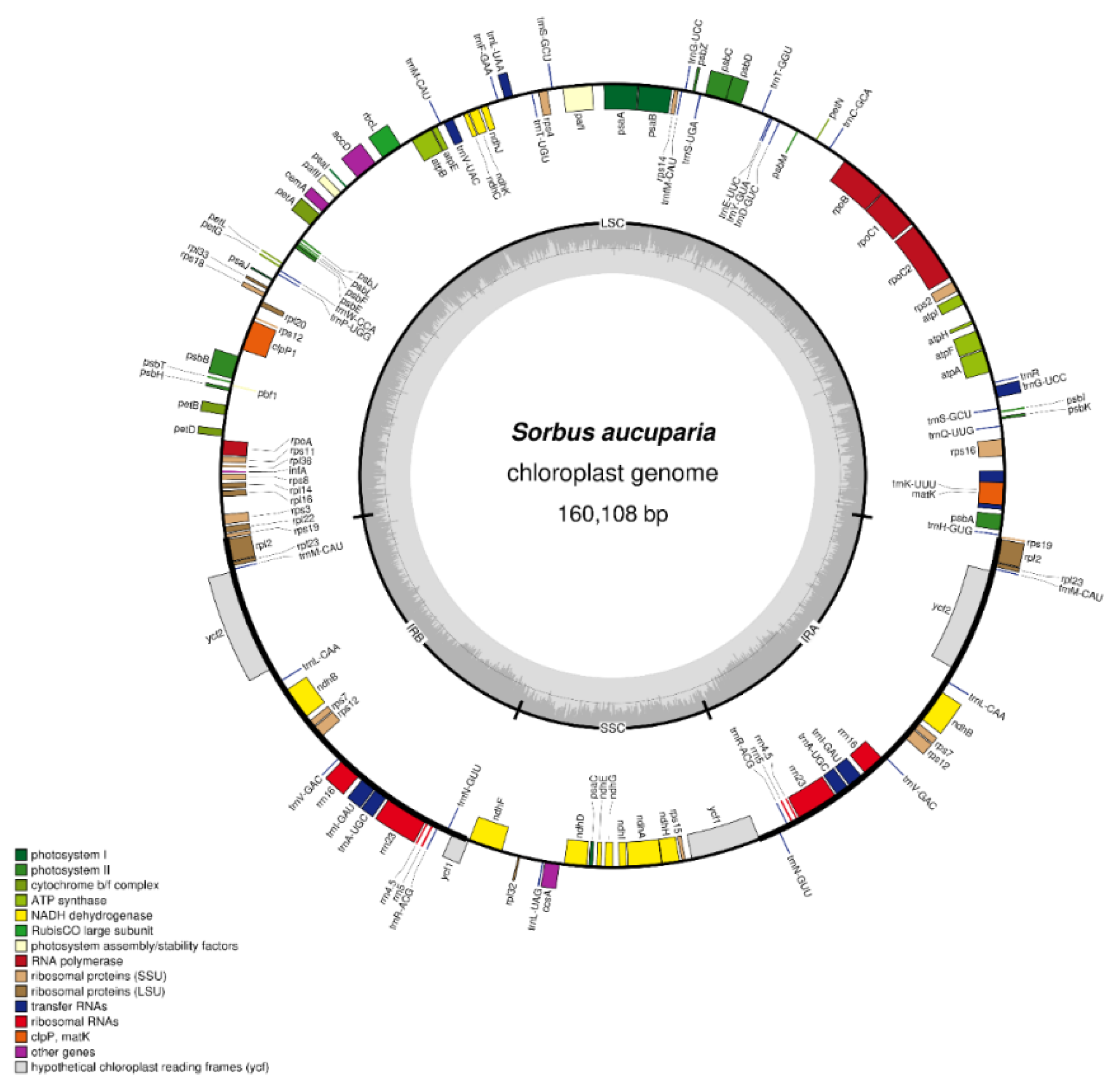
2.1. Chloroplast Genome of Sorbus aucuparia
2.2. Phylogeny of Maleae Tribe Based on Complete Chloroplast Genomes
2.3. Detailed Comparative Analysis of Four Chloroplast Genomes
3. Discussion
4. Materials and Methods
4.1. De Novo Assembly of the Chloroplast Genome of Sorbus aucuparia
4.2. Phylogeny of the Maleae Tribe Based on Complete Chloroplast Genomes
4.3. Detailed Comparative Analysis of Four Chloroplast Genomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruggiero, M.A.; Gordon, D.P.; Orrell, T.M.; Bailly, N.; Bourgoin, T.; Brusca, R.C.; Cavalier-Smith, T.; Guiry, M.D.; Kirk, P.M. A Higher Level Classification of All Living Organisms. PLoS ONE 2015, 10, e0119248. [Google Scholar] [CrossRef] [Green Version]
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Robertson, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Zhang, S.-D.; Jin, J.-J.; Chen, S.-Y.; Chase, M.W.; Soltis, D.E.; Li, H.-T.; Yang, J.-B.; Li, D.-Z.; Yi, T.-S. Diversification of Rosaceae since the Late Cretaceous based on plastid phylogenomics. New Phytol. 2017, 214, 1355–1367. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Shi, S.; Li, J.; Yu, J.; Wang, L.; Yang, X.; Guo, L.; Zhou, S. Phylogeny of Maleae (Rosaceae) Based on Multiple Chloroplast Regions: Implications to Genera Circumscription. BioMed Res. Int. 2018, 2018, 7627191. [Google Scholar] [CrossRef]
- Robertson, K.R.; Phipps, J.B.; Rohrer, J.R.; Smith, P.G. A Synopsis of Genera in Maloideae (Rosaceae). Syst. Bot. 1991, 16, 376–394. [Google Scholar] [CrossRef]
- Sennikov, A.N.; Kurtto, A. A phylogenetic checklist of Sorbus sl (Rosaceae) in Europe. Memo. Soc. Pro Fauna Flora Fenn. 2017, 93, 1–78. [Google Scholar]
- Rushforth, K. The Whitebeam problem, and a solution. Phytologia 2018, 100, 222–246. [Google Scholar]
- Liu, B.-B.; Campbell, C.S.; Hong, D.-Y.; Wen, J. Phylogenetic relationships and chloroplast capture in the Amelanchier-Malacomeles-Peraphyllum clade (Maleae, Rosaceae): Evidence from chloroplast genome and nuclear ribosomal DNA data using genome skimming. Mol. Phylogenet. Evol. 2020, 147, 106784. [Google Scholar] [CrossRef]
- Campbell, C.S.; Evans, R.C.; Morgan, D.R.; Dickinson, T.A.; Arsenault, M.P. Phylogeny of subtribe Pyrinae (formerly the Maloideae, Rosaceae): Limited resolution of a complex evolutionary history. Plant Syst. Evol. 2007, 266, 119–145. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Soltis, D. Phylogenetic consequences of cytoplasmic gene flow in plants. Evol. Trends Plants 1991, 5, 65–84. [Google Scholar]
- Stegemann, S.; Keuthe, M.; Greiner, S.; Bock, R. Horizontal transfer of chloroplast genomes between plant species. Proc. Natl. Acad. Sci. USA 2012, 109, 2434–2438. [Google Scholar] [CrossRef] [Green Version]
- Lo, E.Y.Y.; Donoghue, M.J. Expanded phylogenetic and dating analyses of the apples and their relatives (Pyreae, Rosaceae). Mol. Phylogenet. Evol. 2012, 63, 230–243. [Google Scholar] [CrossRef]
- Zhang, X.; Rong, C.; Qin, L.; Mo, C.; Fan, L.; Yan, J.; Zhang, M. Complete Chloroplast Genome Sequence of Malus hupehensis: Genome Structure, Comparative Analysis, and Phylogenetic Relationships. Molecules 2018, 23, 2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulaszewski, B.; Sandurska, E.; Sztupecka, E.; Burczyk, J. The complete chloroplast genome sequence of wild service tree Sorbus torminalis (L.) Crantz. Conserv. Genet. Resour. 2017, 9, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Niu, Z.; Li, J.; Zhu, K.; Chen, X. The complete chloroplast genome sequence of the Chinese endemic species Sorbus setschwanensis (Rosaceae) and its phylogenetic analysis. Nord. J. Bot. 2020, 38. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Y.; Liu, Y.; Xu, C.; Yuan, Q.; Guo, L.; Huang, L. Evolutionary and phylogenetic aspects of the chloroplast genome of Chaenomeles species. Sci. Rep. 2020, 10, 11466. [Google Scholar] [CrossRef]
- Liu, B.-B.; Hong, D.-Y.; Zhou, S.-L.; Xu, C.; Dong, W.-P.; Johnson, G.; Wen, J. Phylogenomic analyses of the Photinia complex support the recognition of a new genus Phippsiomeles and the resurrection of a redefined Stranvaesia in Maleae (Rosaceae). J. Syst. Evol. 2019, 57, 678–694. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shi, C.; Tang, H.; He, C.; Duan, A.; Gong, H. The complete chloroplast genome sequence of Docynia indica (Wall.) Decne. Mitochondrial DNA Part B 2019, 4, 3046–3048. [Google Scholar] [CrossRef]
- Sokołowska, J.; Fuchs, H.; Celiński, K. New Insight into Taxonomy of European Mountain Pines, Pinus mugo Complex, Based on Complete Chloroplast Genomes Sequencing. Plants 2021, 10, 1331. [Google Scholar] [CrossRef]
- Alzahrani, D.A.; Albokhari, E.J.; Yaradua, S.S.; Abba, A. Comparative Analysis of Chloroplast Genomes of Four Medicinal Capparaceae Species: Genome Structures, Phylogenetic Relationships and Adaptive Evolution. Plants 2021, 10, 1229. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.-H.; Woo, M.-A.; Jo, S.; Kim, Y.-K.; Kim, K.-J. The Chloroplast Phylogenomics and Systematics of Zoysia (Poaceae). Plants 2021, 10, 1517. [Google Scholar] [CrossRef]
- Liu, B.-B.; Liu, G.-N.; Hong, D.-Y.; Wen, J. Eriobotrya Belongs to Rhaphiolepis (Maleae, Rosaceae): Evidence From Chloroplast Genome and Nuclear Ribosomal DNA Data. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Holub, J. Reclassifications and new names in vascular plants 1. Preslia 1998, 70, 97–122. [Google Scholar]
- Li, M.; Ohi-Toma, T.; Gao, Y.-D.; Xu, B.; Zhu, Z.-M.; Ju, W.-B.; Gao, X.-F. Molecular phylogenetics and historical biogeography of Sorbus sensu stricto (Rosaceae). Mol. Phylogenet. Evol. 2017, 111, 76–86. [Google Scholar] [CrossRef]
- Xiang, Y.; Huang, C.-H.; Hu, Y.; Wen, J.; Li, S.; Yi, T.; Chen, H.; Xiang, J.; Ma, H. Evolution of Rosaceae Fruit Types Based on Nuclear Phylogeny in the Context of Geological Times and Genome Duplication. Mol. Biol. Evol. 2017, 34, 262–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, K.-K.; Chen, S.-F.; Xu, K.-W.; Zhou, R.-C.; Li, M.-W.; Dhamala, M.K.; Liao, W.-B.; Fan, Q. Phylogenomic analyses based on genome-skimming data reveal cyto-nuclear discordance in the evolutionary history of Cotoneaster (Rosaceae). Mol. Phylogenet. Evol. 2021, 158, 107083. [Google Scholar] [CrossRef]
- Wang, N.; Thomson, M.; Bodles, W.J.; Crawford, R.M.; Hunt, H.V.; Featherstone, A.W.; Pellicer, J.; Buggs, R.J. Genome sequence of dwarf birch (Betula nana) and cross-species RAD markers. Mol. Ecol. 2013, 22, 3098–3111. [Google Scholar] [CrossRef]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997v1. [Google Scholar]
- Milne, I.; Bayer, M.; Cardle, L.; Shaw, P.; Stephen, G.; Wright, F.; Marshall, D. Tablet--next generation sequence assembly visualization. Bioinformatics 2010, 26, 401–402. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq-versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Carelse, O.; Kempf, J.; Toussaint, J.L.; Chetsanga, C.J.; Mubumbila, M.V. The common bean chloroplast trnH (GUG) gene and its eukaryotic putative promoter elements. Nucleic Acids Res. 1992, 20, 5481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| S. aucuparia vs. | T. glaberrima vs. | A. edulis vs. | |||||
|---|---|---|---|---|---|---|---|
| T. glaberrima | A. edulis | C. alpina | A. edulis | C. alpina | C. alpina | ||
| Type | Total non-matching sites [bp] | 3004 | 2673 | 2788 | 1923 | 1855 | 560 |
| Indels [bp] | 2434 | 2128 | 2242 | 1706 | 1636 | 486 | |
| (81.0%) | (79.6%) | (80.4%) | (88.7%) | (88.2%) | (86.8%) | ||
| Substitutions [bp] | 570 | 545 | 546 | 217 | 219 | 74 | |
| (19.0%) | (20.4% | (19.6% | (11.3%) | (11.8%) | (13.2%) | ||
| Coding [bp] | 124 | 107 | 109 | 60 | 62 | 11 | |
| (4.1%) | (4.0%) | (3.9%) | (2.5%) | (2.8%) | (1.8%) | ||
| Location | Non-coding [bp] | 2880 | 2566 | 2679 | 1863 | 1793 | 549 |
| (95.9%) | (96.0%) | (96.1%) | (97.5%) | (97.2%) | (98.2%) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulaszewski, B.; Jankowska-Wróblewska, S.; Świło, K.; Burczyk, J. Phylogeny of Maleae (Rosaceae) Based on Complete Chloroplast Genomes Supports the Distinction of Aria, Chamaemespilus and Torminalis as Separate Genera, Different from Sorbus sp. Plants 2021, 10, 2534. https://doi.org/10.3390/plants10112534
Ulaszewski B, Jankowska-Wróblewska S, Świło K, Burczyk J. Phylogeny of Maleae (Rosaceae) Based on Complete Chloroplast Genomes Supports the Distinction of Aria, Chamaemespilus and Torminalis as Separate Genera, Different from Sorbus sp. Plants. 2021; 10(11):2534. https://doi.org/10.3390/plants10112534
Chicago/Turabian StyleUlaszewski, Bartosz, Sandra Jankowska-Wróblewska, Katarzyna Świło, and Jarosław Burczyk. 2021. "Phylogeny of Maleae (Rosaceae) Based on Complete Chloroplast Genomes Supports the Distinction of Aria, Chamaemespilus and Torminalis as Separate Genera, Different from Sorbus sp." Plants 10, no. 11: 2534. https://doi.org/10.3390/plants10112534