
Application of Native or Exotic Arbuscular Mycorrhizal Fungi Complexes and Monospecific Isolates from Saline Semi-Arid Mediterranean Ecosystems Improved Phoenix dactylifera’s Growth and Mitigated Salt Stress Negative Effects

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Mycorrhizal Fungi Propagules
2.2. Preparation of the Mycorrhizal Inoculum
2.3. Single-Species Cultures and Microscopy Identification
2.4. Inoculation and Experimental Design
2.5. Mycorrhizal Traits, Growth, Mineral Nutrition, Physiological and Biochemical Parameters
2.5.1. Root Mycorrhizal Colonization
2.5.2. Growth Parameters
2.5.3. Mineral Contents Analysis
2.5.4. Chlorophyll Contents
2.5.5. Protein Contents
2.5.6. Malondialdehyde Contents
2.5.7. Antioxidant Enzyme Assay
2.6. Statistical Analysis
3. Results
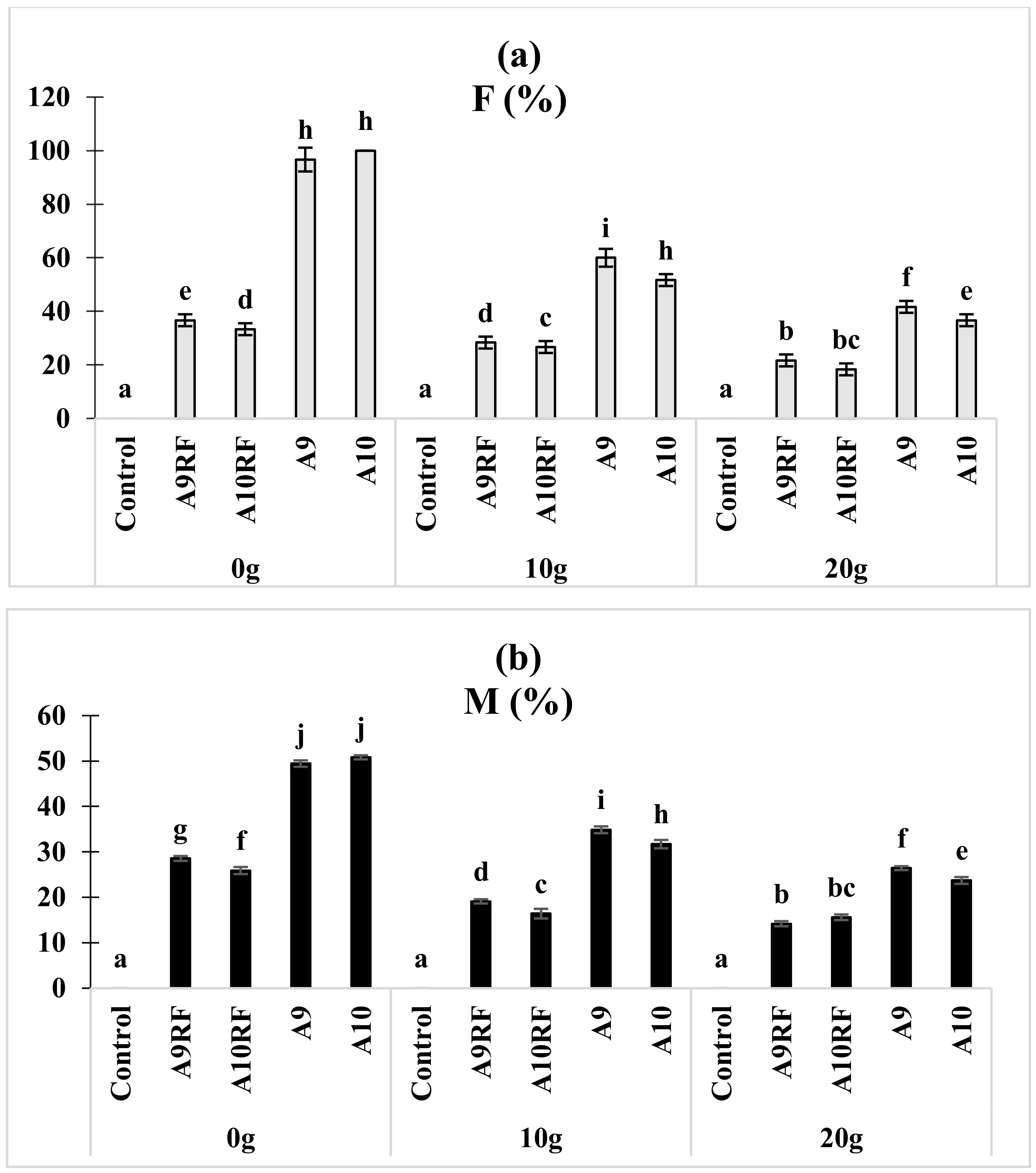
3.1. Effects of Salt Stress on Mycorrhizal Infection of Date Palm Seedlings
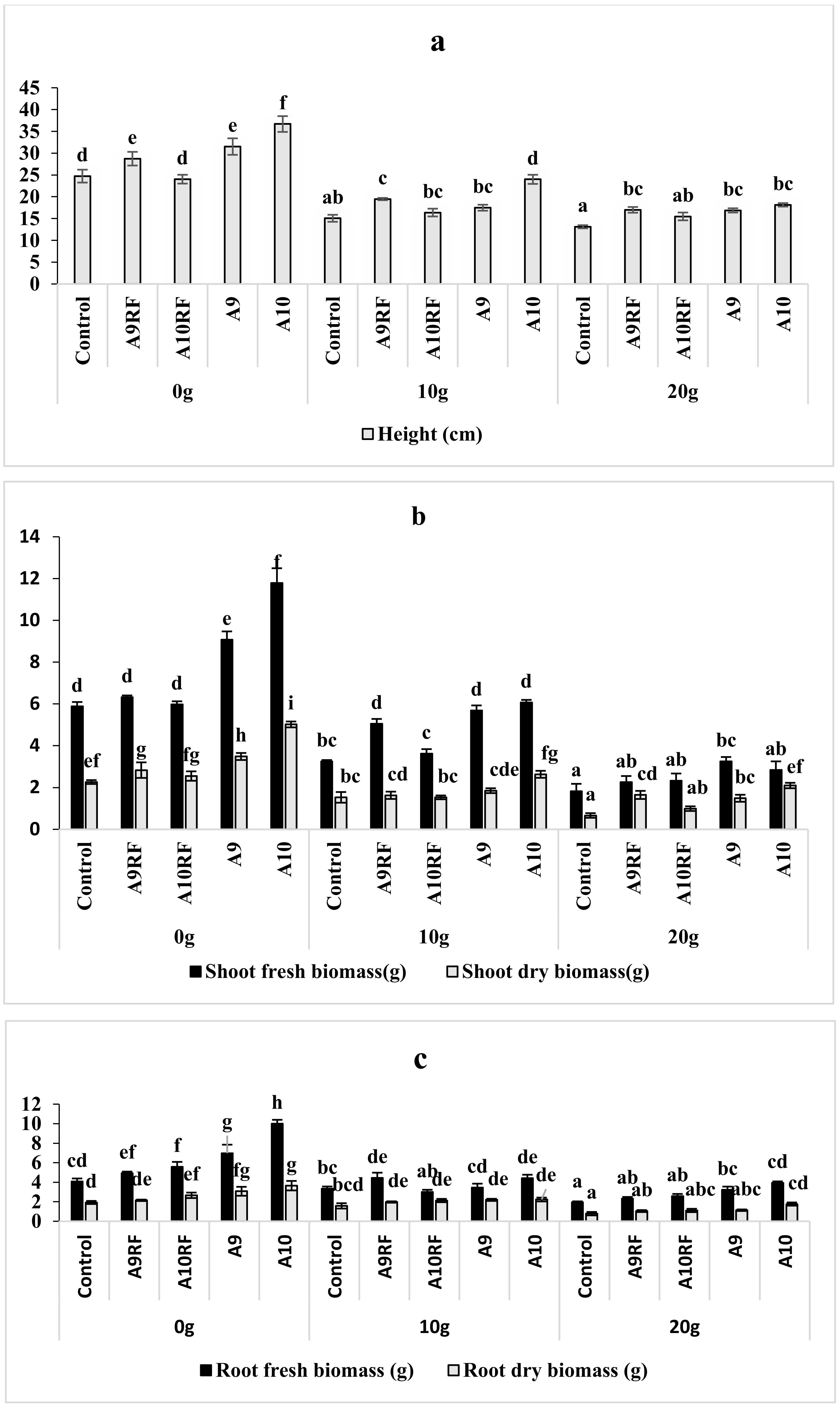
3.2. Effects of Different Inocula on the Growth of Date Palm Seedling under Salt Stress
3.3. Nutrient Contents
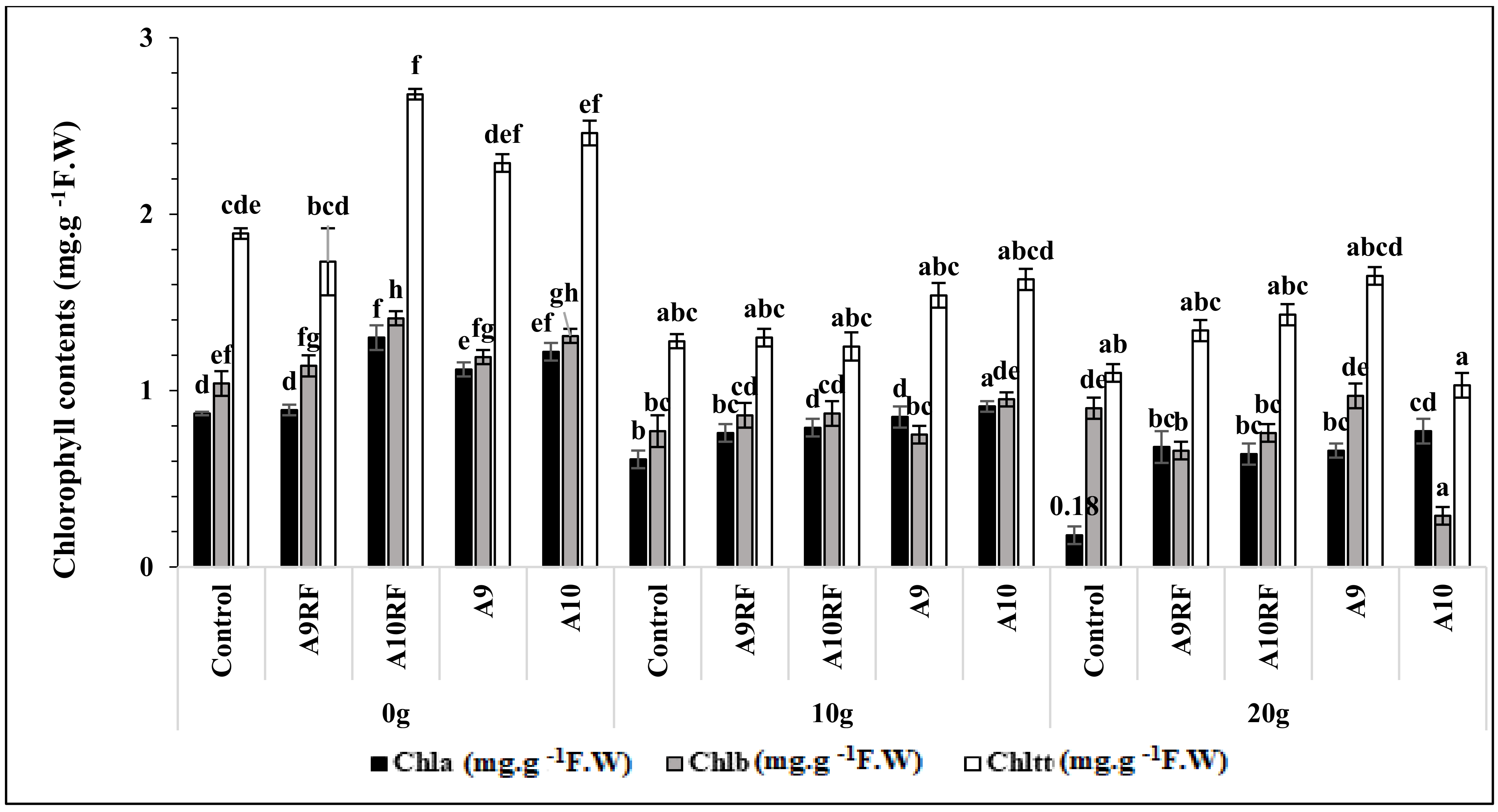
3.4. Chlorophyll Contents
3.5. Malondialdehyde and Protein Contents
3.6. Antioxidant Enzyme Activities
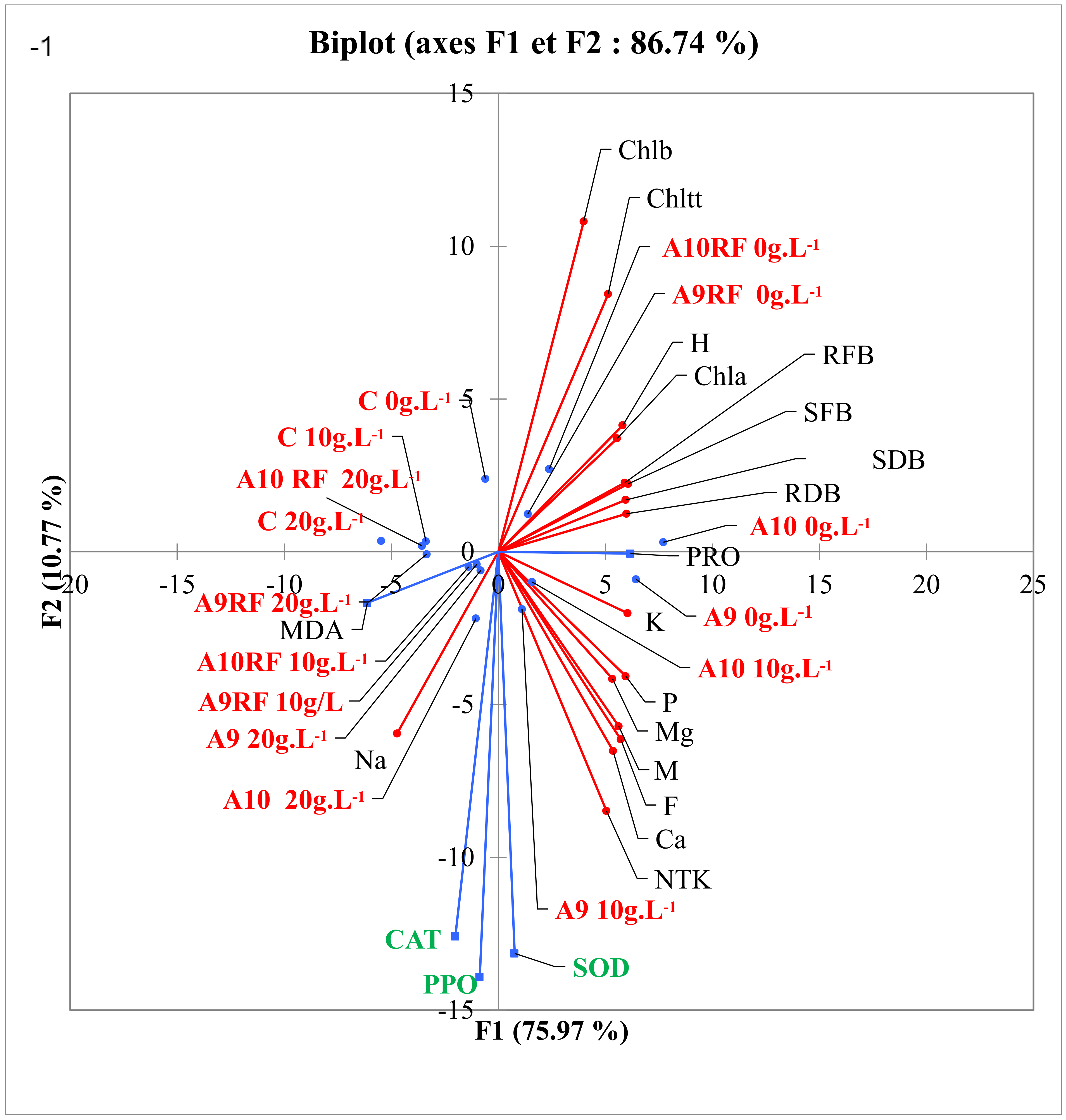
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Setia, R.; Gottschalk, P.; Smith, P.; Marschner, P.; Baldock, J.; Setia, D.; Smith, J. Soil salinity decreases global soil organic carbon stocks. Sci. Total Environ. 2013, 465, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Hussain, S.; Shaukat, M.; Ashraf, M.; Zhu, C.; Jin, Q.; Zhang, J. Salinity Stress in Arid and Semi-Arid Climates: Effects and Management in Field Crops. Clim. Chang. Agric. 2019, 13. [Google Scholar] [CrossRef] [Green Version]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gros-Balthazard, M.; Flowers, J.M. The Date Palm Genome; A brief history of the origin of domesticated date palms; Springer: New York, NY, USA, 2021; Volume 1, pp. 55–74. [Google Scholar]
- Tengberg, M. Beginnings and early history of date palm garden cultivation in the Middle East. J. Arid Environ. 2012, 86, 139–147. [Google Scholar] [CrossRef]
- El Hadrami, A.; Al-Khayri, J.M. Socioeconomic and traditional importance of date palm. Emir. J. Food Agric. 2012, 24, 371–385. [Google Scholar]
- Chao, C.C.T.; Krueger, R.R. The date palm (Phoenix dactylifera L.): Overview of biology, uses, and cultivation. HortScience 2007, 42, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Sedra, M.H. Date Palm Status and Perspective in Morocco. In Date Palm Genetic Resources and Utilization: Africa and the Americas; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Dordrecht, The Netherland, 2015; Volume 1, pp. 257–323. ISBN 978-94-017-9694-1. [Google Scholar]
- Yaish, M.W.; Kumar, P.P. Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 2015, 6, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Al Kharusi, L.; Al Yahyai, R.; Yaish, M.W. Antioxidant response to salinity in salt-tolerant and salt-susceptible cultivars of date palm. Agriculture 2019, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Yaish, M.W. Proline accumulation is a general response to abiotic stress in the date palm tree (Phoenix dactylifera L.). Genet. Mol. Res. 2015, 14, 9943–9950. [Google Scholar] [CrossRef]
- Awasthi, J.P.; Chandra, T.; Mishra, S.; Parmar, S.; Shaw, B.P.; Nilawe, P.D.; Chauhan, N.K.; Sahoo, S.; Panda, S.K. Identification and characterization of drought responsive miRNAs in a drought tolerant upland rice cultivar KMJ 1-12-3. Plant Physiol. Biochem. 2019, 137, 62–74. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Thioye, B.; Sanguin, H.; Kane, A.; de Faria, S.M.; Fall, D.; Prin, Y.; Sanogo, D.; Ndiaye, C.; Duponnois, R.; Sylla, S.N.; et al. Impact of mycorrhiza-based inoculation strategies on Ziziphus mauritiana Lam. and its native mycorrhizal communities on the route of the Great Green Wall (Senegal). Ecol. Eng. 2019, 128, 66–76. [Google Scholar] [CrossRef]
- Sally, E.; Smith, D.J.R. Mycorrhizal Symbiosis; Elsevier: New York, NY, USA, 2008; Volume 137, ISBN 9780123705266. [Google Scholar]
- Baslam, M.; Esteban, R.; García-Plazaola, J.I.; Goicoechea, N. Effectiveness of arbuscular mycorrhizal fungi (AMF) for inducing the accumulation of major carotenoids, chlorophylls and tocopherol in green and red leaf lettuces. Appl. Microbiol. Biotechnol. 2013, 97, 3119–3128. [Google Scholar] [CrossRef]
- Priyadharsini, P.; Muthukumar, T. Arbuscular mycorrhizal fungus influence maize root growth and architecture in rock phosphate amended tropical soil. An. Biol. 2017, 39, 211–222. [Google Scholar] [CrossRef]
- Miransari, M. Contribution of arbuscular mycorrhizal symbiosis to plant growth under different types of soil stress. Plant Biol. 2010, 12, 563–569. [Google Scholar] [CrossRef]
- Jadrane, I.; Al Feddy, M.N.; Dounas, H.; Kouisni, L.; Aziz, F.; Ouahmane, L. Inoculation with selected indigenous mycorrhizal complex improves Ceratonia siliqua’s growth and response to drought stress. Saudi J. Biol. Sci. 2021, 28, 825–832. [Google Scholar] [CrossRef]
- Diatta, I.L.D.; Kane, A.; Agbangba, C.E.; Sagna, M.; Diouf, D.; Aberlenc-Bertossi, F.; Sane, D. Inoculation with arbuscular mycorrhizal fungi improves seedlings growth of two sahelian date palm cultivars (Phoenix dactylifera L., cv. Nakhla hamra and cv. Tijib) under salinity stresses. Adv. Biosci. Biotechnol. 2014, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Symanczik, S.; Błaszkowski, J.; Koegel, S.; Boller, T.; Wiemken, A.; Al-Yahya’Ei, M.N. Isolation and identification of desert habituated arbuscular mycorrhizal fungi newly reported from the Arabian Peninsula. J. Arid Land 2014, 6, 488–497. [Google Scholar] [CrossRef] [Green Version]
- Sieverding, E. Vesicular-Arbuscular Mycorrhiza Management in Tropical Agrosystems dt; Deutsche Gesellschaft für Technische Zusammenarbeit Eschborn; GTZ: Bremen, Germany, 1991. [Google Scholar]
- Mosse, B. Advances in the Study of Vesicular-Arbuscular Mycorrhiza. Annu. Rev. Phytopathol. 1973, 11, 171–196. [Google Scholar] [CrossRef]
- Selvakumar, G.; Kim, K.; Walitang, D.; Chanratana, M.; Kang, Y.; Chung, B.; Sa, T. Trap Culture Technique for Propagation of Arbuscular Mycorrhizal Fungi Using Different Host Plants. Korean J. Soil Sci. Fertil. 2016, 49, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Aguilar, D.; Lara-Capistrán, L.; Maldonado-Mendoza, I.E.; Zulueta-Rodríguez, R.; Sangabriel-Conde, W.; Mancera-López, M.E.; Negrete-Yankelevich, S.; Barois, I. Loss of arbuscular mycorrhizal fungal diversity in trap cultures during long-term subculturing. IMA Fungus 2013, 4, 161–167. [Google Scholar] [CrossRef]
- Feldmann, F. The strain-inherent variability of arbuscular mycorrhizal effectiveness: ii. Effectiveness of single spores. Symbiosis 1998, 25, 131–143. [Google Scholar]
- Tchabi, A.; Burger, S.; Coyne, D.; Hountondji, F.; Lawouin, L.; Wiemken, A.; Oehl, F. Promiscuous arbuscular mycorrhizal symbiosis of yam (Dioscorea spp.), a key staple crop in West Africa. Mycorrhiza 2009, 19, 375–392. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V.; Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de methodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae =Aspects Physiologiques et Genetiques des Mycorhizes, Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, 1–5 July 1985; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; Institut National de la Recherche Agronomique Press: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Olsen, S.R.; Dean, L.A. Phosphorus. In Chemical and Microbiological Properties; Methods of Soil Analysis, Part 2; ACSESS Digital Library: Madison, WI, USA, 1965; Volume 9, pp. 1035–1048. [Google Scholar]
- Raimbault, P.; Lantoine, F.; Neveux, J. Rapid measurement of chlorophyll a and of phaeopigments a by fluorimetry after extraction with methanol. Comparison with the classic acetone extraction method. Océanis 2004, 30, 189–205. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. D032_Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Oxygen Radicals in Biological Systems. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.M.; Flurkey, W.H. Sodium dodecyl sulfate activation of a plant polyphenoloxidase. Effect of sodium dodecyl sulfate on enzymatic and physical characteristics of purified broad bean polyphenoloxidase. J. Biol. Chem. 1990, 265, 4982–4988. [Google Scholar] [CrossRef]
- Yarahmadi, M.A.; Shahsavani, S.; Akhyani, A.; Dorostkar, V. Pomegranate growth affected by arbuscular mycorrhizae, phosphorus fertilizer, and irrigation water salinity. Commun. Soil Sci. Plant Anal. 2018, 49, 478–488. [Google Scholar] [CrossRef]
- Elhindi, K.M.; El-Din, A.S.; Elgorban, A.M. The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J. Biol. Sci. 2017, 24, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Kavroulakis, N.; Tsiknia, M.; Ipsilantis, I.; Kavadia, A.; Stedel, C.; Psarras, G.; Tzerakis, C.; Doupis, G.; Karpouzas, D.G.; Papadopoulou, K.K.; et al. Arbuscular mycorrhizal fungus inocula from coastal sand dunes arrest olive cutting growth under salinity stress. Mycorrhiza 2020, 30, 475–489. [Google Scholar] [CrossRef]
- Tigka, T.; Ipsilantis, I. Effects of sand dune, desert and field arbuscular mycorrhizae on lettuce (Lactuca sativa, L.) growth in a natural saline soil. Sci. Hortic. 2020, 264, 109191. [Google Scholar] [CrossRef]
- Baslam, M.; Qaddoury, A.; Goicoechea, N. Role of native and exotic mycorrhizal symbiosis to develop morphological, physiological and biochemical responses coping with water drought of date palm, Phoenix dactylifera. Trees Struct. Funct. 2014, 28, 161–172. [Google Scholar] [CrossRef]
- Beltrano, J.; Ruscitti, M.; Arango, M.C.; Ronco, M. Effects of arbuscular mycorrhiza inoculation on plant growth, biological and physiological parameters and mineral nutrition in pepper grown under different salinity and p levels. J. Soil Sci. Plant Nutr. 2013, 13, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chaichi, M.R.; Keshavarz-Afshar, R.; Lu, B.; Rostamza, M. Growth and nutrient uptake of tomato in response to application of saline water, biological fertilizer, and surfactant. J. Plant Nutr. 2017, 40, 457–466. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Li, Y.; Wu, A.; Huang, J. Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Sallaku, G.; Sandén, H.; Babaj, I.; Kaciu, S.; Balliu, A.; Rewald, B. Specific nutrient absorption rates of transplanted cucumber seedlings are highly related to RGR and influenced by grafting method, AMF inoculation and salinity. Sci. Hortic. 2019, 243, 177–188. [Google Scholar] [CrossRef]
- Correia, P.J.; Gama, F.; Pestana, M.; Martins-Loução, M.A. Tolerance of young (Ceratonia siliqua L.) carob rootstock to NaCl. Agric. Water Manag. 2010, 97, 910–916. [Google Scholar] [CrossRef]
- Wahid, F.; Sharif, M.; Fahad, S.; Adnan, M.; Khan, I.A.; Aksoy, E.; Ali, A.; Sultan, T.; Alam, M.; Saeed, M.; et al. Arbuscular mycorrhizal fungi improve the growth and phosphorus uptake of mung bean plants fertilized with composted rock phosphate fed dung in alkaline soil environment. J. Plant Nutr. 2019, 42, 1760–1769. [Google Scholar] [CrossRef]
- Xu, H.; Lu, Y.; Tong, S. Effects of arbuscular mycorrhizal fungi on photosynthesis and chlorophyll fluorescence of maize seedlings under salt stress. Emirates J. Food Agric. 2018, 30, 199–204. [Google Scholar] [CrossRef]
- Liu, T.; Sheng, M.; Wang, C.Y.; Chen, H.; Li, Z.; Tang, M. Impact of arbuscular mycorrhizal fungi on the growth, water status, and photosynthesis of hybrid poplar under drought stress and recovery. Photosynthetica 2015, 53, 250–258. [Google Scholar] [CrossRef]
- Kumar, D.; Al Hassan, M.; Naranjo, M.A.; Agrawal, V.; Boscaiu, M.; Vicente, O. Effects of salinity and drought on growth, ionic relations, compatible solutes and activation of antioxidant systems in oleander (Nerium oleander L.). PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef]
- Al Kharusi, L.; Assaha, D.V.M.; Al-Yahyai, R.; Yaish, M.W. Screening of date palm (Phoenix dactylifera L.) cultivars for salinity tolerance. Forests 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Wu, N.; Li, Z.; Wu, F.; Tang, M. Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant Salinity Tolerance Conferred by Arbuscular Mycorrhizal Fungi and Associated Mechanisms: A Meta-Analysis. Front. Plant Sci. 2020, 11, 1–22. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- Ahmad, P.; Prasad, M.N.V.; Azooz, M.M. Salt Stress in Plants: Signalling, Omics and Adaptations; Springer: New York, USA, 2014; ISBN 9781461461081. [Google Scholar]
- Chenchouni, H.; Mekahlia, M.N.; Beddiar, A. Effect of inoculation with native and commercial arbuscular mycorrhizal fungi on growth and mycorrhizal colonization of olive (Olea europaea L.). Sci. Hortic. 2020, 261, 108969. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Barea, J.M.; Ruiz-Lozano, J.M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. 2013, 201–202, 42–51. [Google Scholar] [CrossRef]
- Abbaspour, H.; Saeidi-Sar, S.; Afshari, H.; Abdel-Wahhab, M.A. Tolerance of Mycorrhiza infected Pistachio (Pistacia vera L.) seedling to drought stress under glasshouse conditions. J. Plant Physiol. 2012, 169, 704–709. [Google Scholar] [CrossRef]
- Samuel, A.D.; Brejea, R.; Domuta, C.; Bungau, S.; Cenusa, N.; Tit, D.M. Enzymatic indicators of soil quality. J. Environ. Prot. Ecol. 2017, 18, 871–878. [Google Scholar]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long-term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 28, 1–23. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Soil Origin | Location | pH | Conductivity (ms·cm−1) | TOC (%) | TNK (%) | P (mg·kg−1) | Total Number of Spores 100 g−1 Soil |
|---|---|---|---|---|---|---|---|---|
| Saline site Marrakesh (A9) | Date palm tree | 31.57 ± 7.67 | 7.05 ± 0.05 | 118.03 ± 1.97 | 3.67 ± 0.43 | 0.46 ± 0.14 | 12.24 ± 1.11 | 425 ± 70 |
| Palm grove Zagora (A10) | Date palm tree | 30.38 ± 6.21 | 8.23 ± 0.12 | 27.75 ± 0.22 | 0.96 ± 0.01 | 0.32 ± 0.21 | 4.45 ± 0.89 | 520 ± 15 |
| NaCl Treatment | Inoculum | P (mg·g−1) | NTK (%) | k (%) | Mg (%) | Ca (%) | Na (%) |
|---|---|---|---|---|---|---|---|
| 0 g·L−1 | Control | 1.9 ± 0.89 ab | 0.66 ± 0.07 ab | 2.06 ± 0.07 c | 2.12 ± 0.18 d | 2.67 ± 0.19 bc | 2.14 ± 0.14 a |
| A9RF | 4.27 ± 0.14 c | 0.91 ± 0.04 c | 2.79 ± 0.09 fg | 2.56 ± 0.1 e | 3.04 ± 0.56 cd | 2.43 ± 0.1 a | |
| A10RF | 5.79 ± 0.12 d | 0.8 ± 0.04 bc | 3.08 ± 0.05 gh | 1.35 ± 0.1 b | 3.98 ± 0.11 ef | 2.29 ± 0.13 a | |
| A9 | 9.68 ± 0.24 f | 2.63 ± 0.07 f | 5.28 ± 0.05 j | 3.57 ± 0.09 g | 5.34 ± 0.07 h | 2.42 ± 0.15 a | |
| A10 | 9.83 ± 0.11 f | 2.63 ± 0.1 f | 4.23 ± 0.09 i | 3.12 ± 0.05 f | 4.85 ± 0.08 gh | 2.56 ± 0.01 ab | |
| 10 g·L−1 | Control | 1.51 ± 0.34 a | 0.58 ± 0.12 ab | 1.62 ± 0.1 b | 1.83 ± 0.09 c | 2.13 ± 0.09 ab | 5 ± 0.21 ef |
| A9RF | 2.73 ± 0.29 b | 0.75 ± 0.07 bc | 2.62 ± 0.15 ef | 1.91 ± 0.06 cd | 3.92 ± 0.03 e | 4.73 ± 0.58 def | |
| A10RF | 2.55 ± 0.27 b | 0.66 ± 0.07 abc | 2.58 ± 0.1 ef | 2.11 ± 0.11 cd | 4.04 ± 0.08 ef | 4.38 ± 0.01 de | |
| A9 | 7.43 ± 0.23 e | 1.98 ± 0.09 de | 2.79 ± 0.09 fg | 2.56 ± 0.1 e | 4.5 ± 0 fg | 3.43 ± 0.1 c | |
| A10 | 6.81 ± 0.27 e | 2.11 ± 0.07 e | 3.08 ± 0.06 h | 2.17 ± 0.05 d | 4.18 ± 0.09 ef | 4.05 ± 0.66 cd | |
| 20 g·L−1 | Control | 0.94 ± 0.17 a | 0.44 ± 0.05 a | 0.96 ± 0.13 a | 0.72 ± 0.09 a | 1.67 ± 0.32 a | 5.21 ± 0.01 f |
| A9RF | 1.77 ± 0.38 ab | 0.58 ± 0.09 ab | 1.12 ± 0.11 a | 0.87 ± 0.1 a | 2.86 ± 0.07 cd | 4.38 ± 0.06 de | |
| A10RF | 1.4 ± 0.15 a | 0.56 ± 0.09 ab | 1.17 ± 0.06 a | 0.82 ± 0.04 a | 2.58 ± 0.09 bc | 4.44 ± 0.22 de | |
| A9 | 4.47 ± 0.36 c | 1.86 ± 0.12 de | 2.38 ± 0.15 de | 1.89 ± 0.06 cd | 3.35 ± 0.09 d | 3.29 ± 0.16 bc | |
| A10 | 3.96 ± 0.21 c | 1.75 ± 0.14 d | 2.22 ± 0.06 cd | 2.05 ± 0.1 cd | 3.93 ± 0.06 e | 3.48 ± 0.01 c |
| Na Cl Treatment | Inoculum | MDA Level in (μM·g−1 of FM) | Protein (mg·g−1 of FM) | PPO (μmol de Catechol. min−1·mg−1 Prot) | SOD Activity (U·mg−1 of Protein) | CAT (U·mg−1 of Protein) |
|---|---|---|---|---|---|---|
| 0 g·L−1 | Control | 26.07 ± 0.67 c | 11.7 ± 0.7 f | 0.64 ± 0.09 a | 1.14 ± 0.11 a | 1.47 ± 0.25 a |
| A9RF | 24.09 ± 0.16 b | 14.12 ± 0.59 g | 0.94 ± 0.06 ab | 1.52 ± 0.45 a | 1.78 ± 0.11 ab | |
| A10RF | 23.53 ± 1.05 b | 13.95 ± 0.15 g | 0.76 ± 0.08 a | 1.38 ± 0.3 a | 1.66 ± 0.18 ab | |
| A9 | 19.37 ± 0.44 a | 17.85 ± 0.71 h | 1.45 ± 0.09 def | 2.54 ± 0.27 c | 2.72 ± 0.15 ced | |
| A10 | 18.35 ± 0.13 a | 16.97 ± 0.34 h | 1.41 ± 0.06 cde | 2.62 ± 0.08 c | 2.41 ± 0.1 bc | |
| 10 g·L−1 | Control | 31.99 ± 0.66 g | 9.18 ± 0.34 bcd | 1.11 ± 0.09 bc | 1.58 ± 0.07 ab | 2.35 ± 0.49 abc |
| A9RF | 30.19 ± 0.47 fg | 9.93 ± 0.21 de | 1.35 ± 0.12 cd | 1.78 ± 0.13 ab | 2.68 ± 0.27 cd | |
| A10RF | 29.54 ± 0.51 cd | 9.45 ± 0.62 cd | 1.39 ± 0.15 cde | 1.6 ± 0.18 ab | 2.47 ± 0.28 bc | |
| A9 | 26.15 ± 0.71 c | 13.43 ± 0.62 g | 1.74 ± 0.1 f | 3.54 ± 0.19 d | 3.61 ± 0.35 ef | |
| A10 | 26.68 ± 0.32 cd | 13.66 ± 0.36 g | 1.75 ± 0.12 f | 3.34 ± 0.3 d | 3.39 ± 0.34 def | |
| 20 g·L−1 | Control | 37.28 ± 0.76 i | 6.56 ± 0.42 a | 1.45 ± 0.12 def | 2.19 ± 0.11 bc | 3.46 ± 0.3 def |
| A9RF | 35.68 ± 0.41 hi | 8.38 ± 0.36 bc | 1.73 ± 0.1 f | 2.48 ± 0.23 c | 3.65 ± 0.37 f | |
| A10RF | 34.76 ± 0.51 h | 8.01 ± 0.16 b | 1.68 ± 0.1 ef | 2.46 ± 0.09 c | 3.51 ± 0.21 def | |
| A9 | 28.34 ± 0.78 de | 11.88 ± 0.16 f | 2.38 ± 0.1 g | 3.86 ± 0.1 d | 4.62 ± 0.47 g | |
| A10 | 28.88 ± 0.88 ef | 11.14 ± 0.63 ef | 2.28 ± 0.13 g | 3.47 ± 0.2 d | 3.98 ± 0.24 fg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Outamamat, E.; Bourhia, M.; Dounas, H.; Salamatullah, A.M.; Alzahrani, A.; Alyahya, H.K.; Albadr, N.A.; Al Feddy, M.N.; Mnasri, B.; Ouahmane, L. Application of Native or Exotic Arbuscular Mycorrhizal Fungi Complexes and Monospecific Isolates from Saline Semi-Arid Mediterranean Ecosystems Improved Phoenix dactylifera’s Growth and Mitigated Salt Stress Negative Effects. Plants 2021, 10, 2501. https://doi.org/10.3390/plants10112501
Outamamat E, Bourhia M, Dounas H, Salamatullah AM, Alzahrani A, Alyahya HK, Albadr NA, Al Feddy MN, Mnasri B, Ouahmane L. Application of Native or Exotic Arbuscular Mycorrhizal Fungi Complexes and Monospecific Isolates from Saline Semi-Arid Mediterranean Ecosystems Improved Phoenix dactylifera’s Growth and Mitigated Salt Stress Negative Effects. Plants. 2021; 10(11):2501. https://doi.org/10.3390/plants10112501
Chicago/Turabian StyleOutamamat, Elmostapha, Mohammed Bourhia, Hanane Dounas, Ahmad Mohammad Salamatullah, Abdulhakeem Alzahrani, Heba Khalil Alyahya, Nawal A. Albadr, Mohamed Najib Al Feddy, Bacem Mnasri, and Lahcen Ouahmane. 2021. "Application of Native or Exotic Arbuscular Mycorrhizal Fungi Complexes and Monospecific Isolates from Saline Semi-Arid Mediterranean Ecosystems Improved Phoenix dactylifera’s Growth and Mitigated Salt Stress Negative Effects" Plants 10, no. 11: 2501. https://doi.org/10.3390/plants10112501