
Testing Virulence of Different Species of Insect Associated Fungi against Yellow Mealworm (Coleoptera: Tenebrionidae) and Their Potential Growth Stimulation to Maize


Abstract
:1. Introduction
2. Results
2.1. Virulence Bioassay
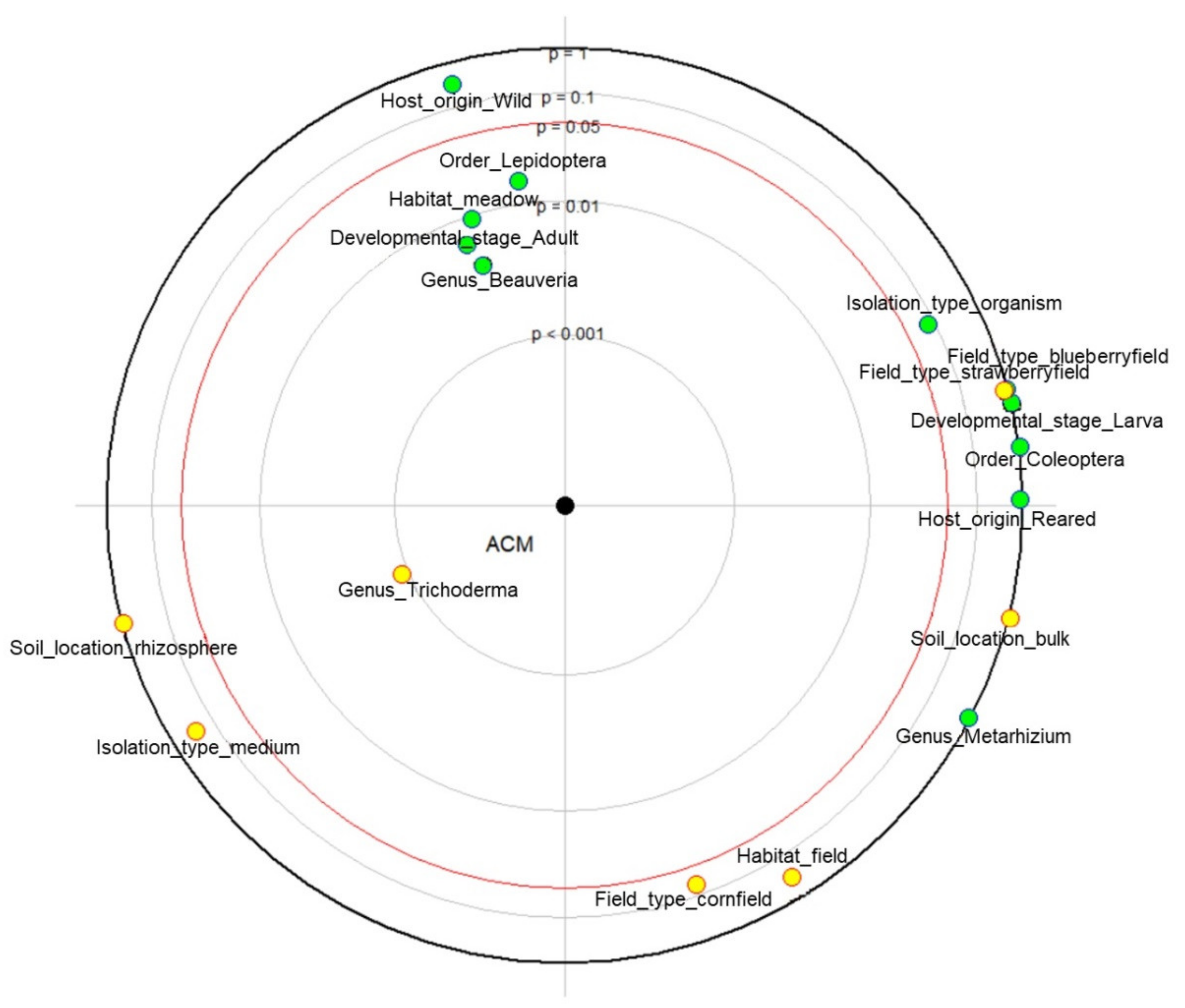
2.2. Influence of Fungal Origin and Isolation Method on Mortality Rate
2.3. Growth Stimulation Bioassay
2.4. Enhancement of Nutrient Utilization by Fungi in Maize
3. Discussion
4. Materials and Methods
4.1. Isolation of Fungi
4.2. Fungal Virulence toward Tenebrio molitor
4.3. Maize Growth Biostimulation Tests
4.3.1. Maize Seed Treatment
4.3.2. Growth Stimulation Bioassay
4.3.3. Fungal Nutrient Utilization Enhancement in Maize
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmad, I.; Jiménez-Gasco, M.D.M.; Luthe, D.S.; Shakeel, S.N.; Barbercheck, M.E. Endophytic Metarhizium robertsii promotes maize growth, suppresses insect growth, and alters plant defense gene expression. Biol. Control 2020, 114, 104167. [Google Scholar] [CrossRef]
- Barra-Bucarei, L.; González, M.G.; Iglesias, A.F.; Aguayo, G.S.; Peñalosa, M.G.; Vera, P.V. Beauveria bassiana multifunction as an endophyte: Growth promotion and biologic control of Trialeurodes vaporariorum, (Westwood) (Hemiptera: Aleyrodidae) in tomato. Insects 2020, 11, 591. [Google Scholar] [CrossRef] [PubMed]
- Reinbacher, L.; Bacher, S.; Knecht, F.; Schweizer, C.; Sostizzo, T.; Grabenweger, G. Preventive field application of Metarhizium brunneum in cover crops for wireworm control. Crop Prot. 2021, 150, 105811. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Jackson, M.A. Mass production of entomopathogenic Hypocreales. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 225–284. [Google Scholar]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.J.; Rehner, S.A.; Bruck, D.J. Diversity of rhizosphere associated entomopathogenic fungi of perennial herbs, shrubs and coniferous trees. J. Invertebr. Pathol. 2011, 106, 289–295. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Afolabi, O.G.; Hussain, M.; Qasim, M.; Wang, L. Prospects of endophytic fungal entomopathogens as biocontrol and plant growth promoting agents: An insight on how artificial inoculation methods affect endophytic colonization of host plants. Microbiol. Res. 2018, 217, 34–50. [Google Scholar] [CrossRef]
- Biere, A.; Bennett, A.E. Three-way interactions between plants, microbes and insects. Funct. Ecol. 2013, 27, 567–573. [Google Scholar] [CrossRef] [Green Version]
- Gruden, K.; Lidoy, J.; Petek, M.; Podpečan, V.; Flors, V.; Papadopoulou, K.K.; Pappas, M.L.; Martinez-Medina, A.; Bejarano, E.; Biere, A.; et al. Ménage à Trois: Unraveling the Mechanisms Regulating Plant-Microbe-Arthropod Interactions. Trends Plant Sci. 2020, 25, 1215–1226. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [Green Version]
- Rocha, I.; Ma, Y.; Souza-Alonso, P.; Vosátka, M.; Freitas, H.; Oliveira, R.S. Seed Coating: A Tool for Delivering Beneficial Microbes to Agricultural Crops. Front. Plant Sci. 2019, 10, 1357. [Google Scholar] [CrossRef] [Green Version]
- Waliwitiya, R.; Isman, M.B.; Vernon, R.S.; Riseman, A. Insecticidal Activity of Selected Monoterpenoids and Rosemary Oil to Agriotes obscurus (Coleoptera: Elateridae). J. Econ. Entomol. 2005, 98, 1560–1565. [Google Scholar] [CrossRef]
- Furlan, L. IPM thresholds for Agriotes wireworm species in maize in Southern Europe. J. Pest Sci. 2014, 87, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Canassa, F.; Tall, S.; Moral, R.A.; Lara, I.A.R.D.; Delalibera, I.; Meyling, N.V. Effects of bean seed treatment by the entomopathogenic fungi Metarhizium robertsii and Beauveria bassiana on plant growth, spider mite populations and behavior of predatory mites. Biol. Control 2019, 132, 199–208. [Google Scholar] [CrossRef]
- Qin, X.; Zhao, X.; Huang, S.; Deng, J.; Li, X.; Luo, Z.; Zhang, Y. Pest management via endophytic colonization of tobacco seedlings by the insect fungal pathogen Beauveria bassiana. Pest Manag. Sci. 2021, 77, 2007–2018. [Google Scholar] [CrossRef]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P.; Mahapatra, B.S. Endophytic colonization of white jute (Corchorus capsularis) plants by different Beauveria bassiana strains for managing stem weevil (Apion corchori). Phytoparasitica 2013, 41, 17–21. [Google Scholar] [CrossRef]
- Faeth, S.H.; Fagan, W.F. Fungal endophytes: Common host plant symbionts but uncommon mutualists. Integr. Comp. Biol. 2002, 42, 360–368. [Google Scholar] [CrossRef] [Green Version]
- Saikkonen, K.; Wäli, P.; Helander, M.; Faeth, S.H. Evolution of endophyte-plant symbioses. Trends Plant Sci. 2004, 9, 275–280. [Google Scholar] [CrossRef]
- Leger, R.J.S. Studies on adaptations of Metarhizium anisopliae to life in the soil. J. Invertebr. Pathol. 2008, 98, 271–276. [Google Scholar] [CrossRef]
- Bruck, D.J. Fungal entomopathogens in the rhizosphere. BioControl 2010, 55, 103–112. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Atanasova, L.; Crom, S.L.; Gruber, S.; Coulpier, F.; Seidl-Seiboth, V.; Kubicek, C.P.; Druzhinina, I.S. Comparative transcriptomics reveals different strategies of Trichoderma mycoparasitism. BMC Genom. 2013, 14, 121. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.M.; El-Katatny, M.H. Entomopathogenic fungi as biopesticides against the Egyptian cotton leaf worm, Spodoptera littoralis: Between biocontrol-promise and immune-limitation. J. Egypt. Soc. Toxicol. 2007, 37, 39–51. [Google Scholar]
- Shakeri, J.; Foster, H.A. Proteolytic activity and antibiotic production by Trichoderma harzianum in relation to pathogenicity to insects. Enzym. Microb. Technol. 2007, 40, 961–968. [Google Scholar] [CrossRef]
- Razinger, J.; Lutz, M.; Schroers, H.J.; Urek, G.; Grunder, J. Evaluation of insect associated and plant growth promoting fungi in the control of cabbage root flies. J. Econ. Entomol. 2014, 107, 1348–1354. [Google Scholar] [CrossRef] [Green Version]
- Berini, F.; Caccia, S.; Franzetti, E.; Congiu, T.; Marinelli, F.; Casartelli, M.; Tettamanti, G. Effects of Trichoderma viride chitinases on the peritrophic matrix of Lepidoptera. Pest Manag. Sci. 2016, 72, 980–989. [Google Scholar] [CrossRef]
- Dhinaut, J.; Balourdet, A.; Teixeira, M.; Chogne, M.; Moret, Y. A dietary carotenoid reduces immunopathology and enhances longevity through an immune depressive effect in an insect model. Sci. Rep. 2017, 7, 12429. [Google Scholar] [CrossRef] [Green Version]
- Sharma, L.; Oliveira, I.; Torres, L.; Marques, G. Entomopathogenic fungi in Portuguese vineyards soils: Suggesting a ‘Galleria-Tenebrio-bait method’ as bait-insects Galleria and Tenebrio significantly underestimate the respective recoveries of Metarhizium (robertsii) and Beauveria (bassiana). MycoKeys 2018, 38, 1–23. [Google Scholar] [CrossRef]
- Medo, J.; Cagáň, Ľ. Factors affecting the occurrence of entomopathogenic fungi in soils of Slovakia as revealed using two methods. Biol. Control 2011, 59, 200–208. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Thomsen, L.; Eilenberg, J.; Boomsma, J.J. Diversity of entomopathogenic fungi near leaf-cutting ant nests in a neotropical forest, with particular reference to Metarhizium anisopliae var. anisopliae. J. Invertebr. Pathol. 2004, 85, 46–53. [Google Scholar] [CrossRef]
- Esparza Mora, M.A.; Costa Rouws, J.R.; Fraga, M.E. Occurrence of entomopathogenic fungi in atlantic forest soils. Microbiol. Discov. 2016, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, R.A. Laboratory Evaluation of Entomopathogenic Fungi, Commercial Formulations, against the Rhinoceros Beetle, Oryctes agamemnon arabicus (Coleoptera: Scarabaeidae). Egypt. J. Biol. Pest Control 2017, 27, 49–55. [Google Scholar]
- Clifton, E.H.; Jaronski, S.T.; Hajek, A.E. Virulence of Commercialized Fungal Entomopathogens Against Asian Longhorned Beetle (Coleoptera: Cerambycidae). J. Insect Sci. 2020, 20, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Medo, J.; Medová, J.; Michalko, J.; Cagáň, Ľ. Variability in virulence of Beauveria spp. soil isolates against Ostrinia nubilalis. J. Appl. Entomol. 2021, 145, 92–103. [Google Scholar] [CrossRef]
- Lopes, R.B.; Mesquita, A.L.M.; Tigano, M.S.; Souza, D.A.; Martins, I.; Faria, M. Diversity of indigenous Beauveria and Metarhizium spp. in a commercial banana field and their virulence toward Cosmopolites sordidus (Coleoptera: Curculionidae). Fungal Ecol. 2013, 6, 356–364. [Google Scholar] [CrossRef]
- Hallouti, A.; Ait Hamza, M.; Zahidi, A.; Ait Hammou, R.; Bouharroud, R.; Ait Ben Aoumar, A.; Boubaker, H. Diversity of entomopathogenic fungi associated with Mediterranean fruit fly (Ceratitis capitata (Diptera: Tephritidae)) in Moroccan Argan forests and nearby area: Impact of soil factors on their distribution. BMC Ecol. 2020, 20, 64. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Ruiz-GarcíA, A.; Santiago-Álvarez, C. Laboratory evaluation of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against puparia and adults of Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol. 2006, 99, 1955–1966. [Google Scholar] [CrossRef]
- Meyling, N.V.; Lübeck, M.; Buckley, E.P.; Eilenberg, J.; Rehner, S.A. Community composition, host range and genetic structure of the fungal entomopathogen Beauveria in adjoining agricultural and seminatural habitats. Mol. Ecol. 2009, 18, 1282–1293. [Google Scholar] [CrossRef]
- Goble, T.A.; Dames, J.F.; Hill, M.P.; Moore, S.D. The effects of farming system, habitat type and bait type on the isolation of entomopathogenic fungi from citrus soils in the Eastern Cape Province, South Africa. BioControl 2010, 55, 399–412. [Google Scholar] [CrossRef]
- Vargas, W.A.; Mandawe, J.C.; Kenerley, C.M. Plant-derived sucrose is a key element in the symbiotic association between Trichoderma virens and maize plants. Plant Physiol. 2009, 151, 792–808. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.Q.; Wang, M.; Sun, J.; Gao, J.X.; Chen, J. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium Stalk rot. Sci. Rep. 2017, 7, 1771. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, J.; Shao, J.; Feng, H.; Zhang, R.; Shen, Q. Extracellular proteins of Trichoderma guizhouense elicit an immune response in maize (Zea mays) plants. Plant Soil 2020, 449, 133–149. [Google Scholar] [CrossRef]
- Ousley, M.A.; Lynch, J.M.; Whipps, J.M. Effect of Trichoderma on plant growth: A balance between inhibition and growth promotion. Microb. Ecol. 1993, 26, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Razinger, J.; Praprotnik, E.; Schroers, H.J. Bioaugmentation of Entomopathogenic Fungi for Sustainable Agriotes Larvae (Wireworms) Management in Maize. Front. Plant Sci. 2020, 11, 535005. [Google Scholar] [CrossRef] [PubMed]
- Kuzhuppillymyal-Prabhakarankutty, L.; Tamez-Guerra, P.; Gomez-Flores, R.; Rodriguez-Padilla, M.C.; Ek-Ramos, M.J. Endophytic Beauveria bassiana promotes drought tolerance and early flowering in corn. World J. Microbiol. Biotechnol. 2020, 36, 47. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; White, J.F. Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 2007, 16, 1701–1711. [Google Scholar] [CrossRef]
- Wang, B.; Kang, Q.; Lu, Y.; Bai, L.; Wang, C. Unveiling the biosynthetic puzzle of destruxins in Metarhizium species. Proc. Natl. Acad. Sci. USA 2012, 109, 1287–1292. [Google Scholar] [CrossRef] [Green Version]
- Machado, D.F.M.; Tavares, A.P.; Lopes, S.J.; da Silva, A.C.F. Trichoderma spp. na emergência e crescimento de mudas de cambará (Gochnatia polymorpha (Less.) Cabrera). Rev. Árvore 2015, 39, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Campos, B.F.; Araújo, A.J.C.; Felsemburgh, C.A.; Vieira, T.A.; Lustosa, D.C. Trichoderma contributes to the germination and seedling development of açaí palm. Agriculture 2020, 10, 456. [Google Scholar] [CrossRef]
- Eagles, H.A.; Brooking, I.R. Populations of maize with more rapid and reliable seedling emergence than cornbelt dents at low temperatures. Euphytica 1981, 30, 755–763. [Google Scholar] [CrossRef]
- Knezevic, S.Z.; Weise, S.F.; Swanton, C.J. Interference of Redroot Pigweed (Amaranthus retroflexus) in Corn (Zea mays). Weed Sci. 1994, 42, 568–573. [Google Scholar] [CrossRef]
- Chiduza, C.; Dube, E. Maize production challenges in high biomass input smallholder farmer conservation agriculture systems: A practical research experience from South Africa. Afr. Crop Sci. Soc. 2013, 11, 23–27. [Google Scholar]
- Rivas-Franco, F.; Hampton, J.G.; Morán-Diez, M.E.; Narciso, J.; Rostás, M.; Wessman, P.; Jackson, T.A.; Glare, T.R. Effect of coating maize seed with entomopathogenic fungi on plant growth and resistance against Fusarium graminearum and Costelytra giveni. Biocontrol Sci. Technol. 2019, 29, 877–900. [Google Scholar] [CrossRef]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Cabello, M.N.; Ferreri, N.; Pelizza, S. Endophytic Effects of Beauveria bassiana on Corn (Zea mays) and Its Herbivore, Rachiplusia nu (Lepidoptera: Noctuidae). Insects 2019, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Tall, S.; Meyling, N.V. Probiotics for Plants? Growth Promotion by the Entomopathogenic Fungus Beauveria bassiana Depends on Nutrient Availability. Microb. Ecol. 2018, 76, 1002–1008. [Google Scholar] [CrossRef]
- Kabaluk, J.T.; Ericsson, J.D. Metarhizium anisopliae seed treatment increases yield of field corn when applied for wireworm control. Agron. J. 2007, 99, 1377–1381. [Google Scholar] [CrossRef]
- Zimmermann, G. The ‘Galleria bait method’ for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Cooke, B.W. Fungi in Polluted Water and Sewage: II. Isolation Technique. Sewage Ind. Waste. 1954, 26, 661–674. [Google Scholar]
- Williams, J.; Clarkson, J.M.; Mills, P.R.; Cooper, R.M. A selective medium for quantitative reisolation of Trichoderma harzianum from Agaricus bisporus compost. Appl. Environ. Microbiol. 2003, 69, 4190–4191. [Google Scholar] [CrossRef] [Green Version]
- Razinger, J.; Schroers, H.J.; Urek, G. Virulence of Metarhizium brunneum to field collected Agriotes spp. Wireworms. J. Agric. Sci. Technol. 2018, 20, 309–320. [Google Scholar]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. Metarhizium frigidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. flavoviride complex. Mycologia 2006, 98, 737–745. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistlerr, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bastidas, Ó. Cell Counting with Neubauer Chamber—Basic Hemocytometer Usage. Available online: https://moam.info/cell-counting-with-neubauer-chamber-celeromics_597734c21723dde38bd38984.html (accessed on 20 September 2021).
- Panevska, A.; Hodnik, V.; Skočaj, M.; Novak, M.; Modic, Š.; Pavlic, I.; Podržaj, S.; Zarić, M.; Resnik, N.; Maček, P.; et al. Pore-forming protein complexes from Pleurotus mushrooms kill western corn rootworm and Colorado potato beetle through targeting membrane ceramide phosphoethanolamine. Sci. Rep. 2019, 9, 5073. [Google Scholar] [CrossRef] [Green Version]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Falissard, B. psy: Various Procedures Used in Psychometry. R Package Version 1.1. 2012. Available online: https://CRAN.R-project.org/package=psy (accessed on 28 September 2021).
- Brown, C. dummies: Create Dummy/Indicator Variables Flexibly and Efficiently. R Package Version 1.5.6. 2012. Available online: https://CRAN.R-project.org/package=dummies (accessed on 26 October 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 8 October 2021).
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation. R Package Version 1.0.5. 2021. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 28 September 2021).
- Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R Package Version 0.7.0. 2021. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 28 September 2021).


{kind=link}
| Isolate | Taxon | Habitat or Origin | Isolation Type/ Host Organism | Host Developmental Stage | Host Origin | ACM 7.00 (%) | ACM 14.00 (%) | LT50 (d) |
|---|---|---|---|---|---|---|---|---|
| 1154 | MB | soil | Galleria mellonella | larvae | reared | 61.54 | 100.00 | 6.00 * |
| 1868 | MB | meadow | Agriotes sp. | adult | wild | 26.92 | 86.96 | 8.00 * |
| 2121 | BB | cauliflower field | Curculionidae | adult | wild | 65.38 | 100.00 | 6.00 * |
| 2631 | MR | maize field | Tenebrio molitor | larvae | reared | 10.34 | 52.00 | 12.5 * |
| 2632 | MR | maize field | Tenebrio molitor | larvae | reared | 10.34 | 32.00 | |
| 2245 | MR | maize field | Tenebrio molitor | larvae | reared | 6.90 | 28.00 | |
| 2246 | MR | maize field | Tenebrio molitor | larvae | reared | 0.00 | 20.00 | |
| 2215 | MR | maize field | Tenebrio molitor | larvae | reared | 11.11 | 37.50 | 14.00 |
| 2216 | MR | maize field | Tenebrio molitor | larvae | reared | 7.41 | 54.17 | 10.00 * |
| 2299 | BB | meadow | Galleria mellonella | larvae | reared | 18.52 | 54.17 | 10.00 * |
| 2300 | BB | meadow | Galleria mellonella | larvae | reared | 33.33 | 79.17 | 10.00 * |
| 2635 | MR | maize field | Tenebrio molitor | larvae | reared | 21.43 | 65.38 | 11.50 * |
| 2637 | MR | maize field | Tenebrio molitor | larvae | reared | 17.86 | 84.62 | 11.00 * |
| 2641 | MR | maize field | Tenebrio molitor | larvae | reared | 21.43 | 65.38 | 11.00 * |
| 2697 | ND | maize field | Tenebrio molitor | larvae | reared | −18.18 | −23.08 | |
| 2298 | BB | meadow | Galleria mellonella | larvae | reared | 42.11 | 88.24 | 6.00 * |
| 2243 | MR | maize field | Tenebrio molitor | larvae | reared | 0.00 | 37.50 | 14.00 |
| 2151 | MR | maize field | Tenebrio molitor | larvae | reared | −18.18 | 20.00 | 13.00 |
| 2703 | MB | soil | ND | ND | ND | 0.00 | 21.43 | * |
| 2009 | MR | soil | selective medium | - | - | 10.34 | 32.14 | 14.00 * |
| 2010 | MG | soil | selective medium | - | - | 3.45 | 32.14 | * |
| 2011 | MR | soil | Galleria mellonella | larvae | reared | 16.67 | 31.03 | |
| 2686 | MR | maize field | Tenebrio molitor | larvae | reared | 7.41 | 43.48 | 13.00 * |
| 2687 | MR | maize field | Tenebrio molitor | larvae | reared | 11.11 | 56.52 | 11.00 * |
| 2690 | MB | soil | ND | ND | ND | 7.41 | 47.83 | 12.00 * |
| 2692 | MR | maize field | Diabrotica v. virgifera | adult | wild | 7.41 | 39.13 | 14.00 * |
| 2152 | MR | maize field | Tenebrio molitor | larvae | reared | −3.45 | 26.92 | * |
| 2146 | MR | maize field | Tenebrio molitor | larvae | reared | 20.00 | 54.17 | 11.00 * |
| 2147 | MR | maize field | Tenebrio molitor | larvae | reared | 23.33 | 58.33 | 11.00 * |
| 2251 | MR | maize field | Tenebrio molitor | larvae | reared | 16.67 | 75.00 | 11.00 * |
| 2789 | MR | maize field | Diabrotica v. virgifera | larvae | wild | 16.67 | 41.67 | 13.00 * |
| 2793 | MR | maize field | selective medium | - | - | 20.00 | 70.83 | 11.00 * |
| 2794 | MR | maize field | selective medium | - | - | 10.00 | 20.83 | |
| 2795 | MR | maize field | selective medium | - | - | 36.67 | 66.67 | 11.00 * |
| 2645 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 26.92 | |
| 2691 | MR | blueberry field | Tenebrio molitor | larvae | reared | 6.67 | 53.85 | 11.00 * |
| 2693 | MR | blueberry field | Tenebrio molitor | larvae | reared | 6.67 | 73.08 | 11.00 * |
| 2790 | MR | maize field | Diabrotica v. virgifera | larvae | wild | 0 | 19.23 | 14.00 |
| 2634 | MR | maize field | Tenebrio molitor | larvae | reared | 10.34 | 8.33 | |
| 2214 | MR | maize field | Tenebrio molitor | larvae | reared | 10.34 | 33.33 | 14.00 |
| 2702 | MR | maize field | Tenebrio molitor | larvae | reared | 0 | 8.33 | |
| 2791 | MR | maize field | selective medium | - | - | 37.93 | 62.50 | 12.00 * |
| 2250 | MG | maize field | Tenebrio molitor | larvae | reared | 3.33 | 3.70 | |
| 2685 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 14.81 | |
| 2640 | MR | maize field | Tenebrio molitor | larvae | reared | 17.24 | 29.63 | |
| 2694 | MR | strawberry field | Tenebrio molitor | larvae | reared | −3.45 | 25.93 | |
| 2695 | MR | strawberry field | Tenebrio molitor | larvae | reared | 6.90 | 22.22 | |
| 2788 | MR | maize field | Diabrotica v. virgifera | larvae | wild | 10.34 | 29.63 | |
| 2792 | MR | maize field | selective medium | - | - | 3.45 | 29.63 | |
| 2796 | MR | maize field | selective medium | - | - | 3.45 | 22.22 | |
| 2688 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 5.27 | * |
| 2154 | MR | maize field | Tenebrio molitor | larvae | reared | 3.57 | −23.81 | |
| 2153 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 23.33 | * |
| 2217 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 10.00 | |
| 2698 | MR | basil leaf | unknown larva | larvae | wild | 3.33 | 13.33 | |
| 2699 | MR | blueberry field | Tenebrio molitor | larvae | reared | 3.33 | 10.00 | |
| 2700 | MR | maize field | Tenebrio molitor | larvae | reared | 10 | 20.00 | |
| 2701 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 13.33 | |
| 2239 | MR | maize field | Tenebrio molitor | larvae | reared | 0 | 20.00 | |
| 2704 | BB | meadow | unknown larva | larvae | wild | 0 | 53.33 | 14.00 * |
| 2247 | MG | maize field | Tenebrio molitor | larvae | reared | 3.33 | 13.33 | |
| 2752 | TA | decaying corn ear | natural substratum | - | - | 0 | 0.00 | |
| 2815 | TB | maize field | selective medium | - | - | 0 | 3.33 | |
| 2878 | TH | maize field | selective medium | - | - | 0 | 0.00 | |
| 2882 | TA | maize field | selective medium | - | - | 0 | 0.00 | |
| 2883 | TG | maize field | selective medium | - | - | 0 | 0.00 | |
| 2150 | MR | maize field | Galleria mellonella | larvae | reared | 16.67 | 46.67 | * |
| 2240 | MR | maize field | Tenebrio molitor | larvae | reared | 23.33 | 50.00 | 14.00 * |
| 2148 | MR | maize field | Galleria mellonella | larvae | reared | 23.33 | 53.33 | 14.00 * |
| 2636 | MR | maize field | Tenebrio molitor | larvae | reared | 16.67 | 63.33 | 12.50 * |
| 2642 | MR | maize field | Tenebrio molitor | larvae | reared | 3.33 | 23.33 | * |
| Actara 25 WG | - | - | - | - | - | 11.86 | 47.37 | 13.50 * |
| Mycotal | LM | - | - | - | - | −1.69 | −3.51 | |
| Force | - | - | - | - | - | 1.69 | 1.75 | |
| Met52 EC | MB | - | - | - | - | 2.54 | 11.18 |
| Isolate | Taxon | Maize Variety | Emergence Success [n] | Emergence Time [d] | Root Length [cm] | Shoot Length [cm] | Total Plant Length [cm] | Plant Dry Weight [g] |
|---|---|---|---|---|---|---|---|---|
| Control | Chapalu | 4.67 ± 0.33 | 4.33 ± 0.33 | 22.57 ± 1.15 | 32.87 ± 1.15 | 55.43 ± 2.45 | 0.30 ± 0.04 | |
| 1154 | MB | Chapalu | 4.67 ± 0.33 | 4.13 ± 0.13 | 21.67 ± 1.63 | 30.90 ± 0.78 | 52.57 ± 2.28 | 0.29 ± 0.02 |
| 1868 | MB | Chapalu | 4.67 ± 0.33 | 4.33 ± 0.33 | 21.50 ± 0.15 | 29.60 ± 1.19 | 51.09 ± 1.21 | 0.28 ± 0.02 |
| 2121 | BB | Chapalu | 5.00 ± 0.00 | 4.40 ± 0.23 | 21.50 ± 1.10 | 32.27 ± 0.74 | 53.75 ± 1.73 | 0.30 ± 0.01 |
| Control | Chapalu | 5.00 ± 0.00 | 4.27 ± 0.07 | 20.97 ± 0.45 | 29.97 ± 0.13 | 50.95 ± 0.37 | 0.27 ± 0.02 | |
| 2631 | MR | Chapalu | 4.67 ± 0.33 | 4.38 ± 0.32 | 21.23 ± 0.47 | 26.37 ± 0.43 | 47.58 ± 0.88 | 0.24 ± 0.01 |
| 2632 | MR | Chapalu | 4.33 ± 0.33 | 4.40 ± 0.31 | 21.40 ± 1.51 | 27.97 ± 0.65 | 49.37 ± 1.69 | 0.26 ± 0.01 |
| 2245 | MR | Chapalu | 5.00 ± 0.00 | 4.53 ± 0.07 | 18.80 ± 0.56 | 27.23 ± 1.43 | 46.04 ± 1.62 | 0.22 ± 0.02 |
| 2246 | MR | Chapalu | 4.67 ± 0.33 | 4.17 ± 0.17 | 18.50 ± 0.40 | 28.10 ± 0.81 | 46.56 ± 1.19 | 0.23 ± 0.01 |
| Control | Chapalu | 5.00 ± 0.00 | 4.93 ± 0.07 | 21.23 ± 0.64 | 31.47 ± 0.83 | 52.70 ± 1.02 | 0.27 ± 0.01 | |
| 2215 | MR | Chapalu | 4.67 ± 0.33 | 4.43 ± 0.03 | 21.55 ± 0.75 | 29.30 ± 0.82 | 50.85 ± 0.08 | 0.25 ± 0.01 |
| 2216 | MR | Chapalu | 4.67 ± 0.33 | 4.57 ± 0.23 | 22.39 ± 0.55 | 29.82 ± 0.32 | 52.21 ± 0.73 | 0.25 ± 0.01 |
| 2299 | BB | Chapalu | 4.33 ± 0.33 | 4.40 ± 0.31 | 21.53 ± 0.48 | 26.43 ± 1.41 | 47.96 ± 1.81 | 0.21 ± 0.01 |
| 2300 | BB | Chapalu | 4.67 ± 0.33 | 4.40 ± 0.23 | 22.25 ± 1.02 | 27.40 ± 1.09 | 49.64 ± 2.10 | 0.22 ± 0.01 |
| Control | Chapalu | 5.00 ± 0.00 | 4.13 ± 0.07 | 18.60 ± 0.95 | 29.57 ± 0.78 | 48.16 ± 1.71 | 0.25 ± 0.01 | |
| 2635 | MR | Chapalu | 4.67 ± 0.33 | 4.37 ± 0.09 | 21.40 ± 1.14 | 28.53 ± 0.96 | 49.94 ± 1.57 | 0.23 ± 0.03 |
| 2637 | MR | Chapalu | 4.67 ± 0.33 | 4.62 ± 0.50 | 19.93 ± 0.46 | 28.67 ± 1.08 | 48.59 ± 1.51 | 0.23 ± 0.02 |
| 2641 | MR | Chapalu | 5.00 ± 0.00 | 4.40 ± 0.31 | 19.57 ± 0.71 | 28.27 ± 1.07 | 47.81 ± 0.70 | 0.22 ± 0.02 |
| Control | Chapalu | 4.67 ± 0.33 | 4.00 ± 0.00 | 16.30 ± 0.47 | 30.30 ± 1.30 | 46.61 ± 1.78 | 0.22 ± 0.01 | |
| 2697 | ND | Chapalu | 5.00 ± 0.00 | 4.07 ± 0.07 | 17.90 ± 0.23 | 27.27 ± 2.83 | 45.20 ± 2.70 | 0.23 ± 0.02 |
| 2698 | MR | Chapalu | 4.33 ± 0.67 | 4.44 ± 0.08 | 19.13 ± 0.99 | 27.60 ± 1.80 | 46.73 ± 2.17 | 0.21 ± 0.01 |
| 2699 | MR | Chapalu | 5.00 ± 0.00 | 4.20 ± 0.00 | 18.07 ± 0.61 | 31.27 ± 0.67 | 49.35 ± 1.20 | 0.22 ± 0.01 |
| 2700 | MR | Chapalu | 4.33 ± 0.33 | 4.53 ± 0.15 | 19.17 ± 0.24 | 33.30 ± 1.65 | 52.46 ± 1.83 | 0.26 ± 0.01 |
| Control | Chapalu | 5.00 ± 0.00 | 4.07 ± 0.07 | 20.07 ± 1.28 | 30.33 ± 1.21 | 50.43 ± 2.28 | 0.22 ± 0.03 | |
| 2298 | BB | Chapalu | 4.33 ± 0.33 | 4.85 ± 0.52 | 20.80 ± 1.12 | 30.30 ± 0.66 | 51.09 ± 1.80 | 0.26 ± 0.02 |
| 2243 | MR | Chapalu | 5.00 ± 0.00 | 4.93 ± 0.13 | 15.60 ± 1.21 | 27.00 ± 0.95 | 42.59 ± 1.96 | 0.21 ± 0.01 |
| 2636 | MR | Chapalu | 5.00 ± 0.00 | 4.80 ± 0.23 | 16.77 ± 0.70 | 28.47 ± 0.38 | 45.26 ± 0.91 | 0.20 ± 0.02 |
| 2642 | MR | Chapalu | 4.33 ± 0.33 | 4.50 ± 0.25 | 17.20 ± 0.85 | 34.53 ± 0.32 | 51.72 ± 0.73 | 0.25 ± 0.01 |
| Control | Chapalu | 4.67 ± 0.33 | 4.13 ± 0.07 | 20.70 ± 0.31 | 32.93 ± 1.88 | 53.61 ± 1.80 | 0.31 ± 0.05 | |
| 2148 | MR | Chapalu | 5.00 ± 0.00 | 4.27 ± 0.18 | 21.37 ± 0.45 | 35.77 ± 0.47 | 57.15 ± 0.50 | 0.32 ± 0.02 |
| 2151 | MR | Chapalu | 4.67 ± 0.33 | 4.57 ± 0.12 | 19.47 ± 0.58 | 36.77 ± 0.78 | 56.22 ± 0.50 | 0.32 ± 0.02 |
| 2152 | MR | Chapalu | 4.33 ± 0.33 | 4.28 ± 0.17 | 21.50 ± 0.64 | 35.67 ± 2.11 | 57.19 ± 2.76 | 0.31 ± 0.03 |
| 2701 | MR | Chapalu | 5.00 ± 0.00 | 4.47 ± 0.07 | 20.57 ± 0.62 | 33.60 ± 0.55 | 54.15 ± 0.38 | 0.33 ± 0.01 |
| Control | Chapalu | 4.67 ± 0.33 | 4.20 ± 0.12 | 20.27 ± 0.26 | 34.70 ± 0.59 | 54.98 ± 0.83 | 0.32 ± 0.01 | |
| 2703 | MB | Chapalu | 4.33 ± 0.33 | 4.25 ± 0.25 | 24.43 ± 1.56 | 35.00 ± 1.59 | 59.44 ± 0.11 | 0.36 ± 0.01 |
| 2009 | MR | Chapalu | 5.00 ± 0.00 | 4.47 ± 0.18 | 20.70 ± 0.46 | 33.97 ± 2.72 | 54.65 ± 2.84 | 0.31 ± 0.04 |
| 2010 | MG | Chapalu | 5.00 ± 0.00 | 4.13 ± 0.13 | 20.40 ± 1.68 | 31.53 ± 1.82 | 51.94 ± 3.47 | 0.27 ± 0.04 |
| 2011 | MR | Chapalu | 5.00 ± 0.00 | 4.53 ± 0.13 | 20.93 ± 0.94 | 30.83 ± 1.43 | 51.81 ± 2.37 | 0.23 ± 0.00 |
| Control | Chapalu | 4.67 ± 0.33 | 4.43 ± 0.07 | 13.03 ± 0.03 | 15.90 ± 1.46 | 28.91 ± 1.44 | 0.15 ± 0.01 | |
| 2239 | MR | Chapalu | 5.00 ± 0.00 | 4.59 ± 0.21 | 14.63 ± 1.76 | 18.90 ± 0.20 | 33.50 ± 1.56 | 0.19 ± 0.01 |
| 2704 | BB | Chapalu | 3.33 ± 0.33 | 5.00 ± 0.25 | 11.10 ± 0.50 | 18.00 ± 1.06 | 29.11 ± 1.30 | 0.17 ± 0.00 |
| 2247 | MG | Chapalu | 3.33 ± 0.33 | 4.81 ± 0.13 | 15.33 ± 1.06 | 18.80 ± 0.65 | 34.13 ± 1.61 | 0.19 ± 0.01 |
| 2752 | TA | Chapalu | 3.00 ± 0.58 | 5.29 ± 0.14 | 13.93 ± 0.73 | 18.40 ± 1.34 | 32.33 ± 2.03 | 0.19 ± 0.03 |
| 2815 | TB | Chapalu | 3.67 ± 0.33 | 4.71 ± 0.22 | 13.33 ± 1.48 | 18.20 ± 1.04 | 31.53 ± 2.35 | 0.20 ± 0.02 |
| 2878 | TH | Chapalu | 3.33 ± 0.33 | 4.44 ± 0.34 | 14.70 ± 1.51 | 18.73 ± 0.50 | 33.39 ± 1.62 | 0.17 ± 0.01 |
| 2882 | TA | Chapalu | 3.33 ± 0.33 | 4.84 ± 0.30 | 14.27 ± 1.08 | 20.00 ± 0.60 | 34.32 ± 1.11 | 0.21 ± 0.01 |
| 2883 | TG | Chapalu | 3.33 ± 0.33 | 4.41 ± 0.12 | 14.67 ± 1.21 | 19.47 ± 0.65 | 34.13 ± 1.83 | 0.17 ± 0.03 |
| Control | Belokranjka | 9.00 ± 0.58 | 5.19 ± 0.05 | 23.87 ± 1.58 | 32.73 ± 0.83 | 56.58 ± 2.41 | 0.31 ± 0.03 | |
| 2686 | MR | Belokranjka | 10.00 ± 0.00 | 5.07 ± 0.20 | 24.50 ± 3.10 | 30.97 ± 0.58 | 55.48 ± 2.55 | 0.30 ± 0.01 |
| 2687 | MR | Belokranjka | 10.00 ± 0.00 | 5.33 ± 0.07 | 24.30 ± 2.31 | 30.70 ± 0.23 | 54.96 ± 2.22 | 0.30 ± 0.01 |
| 2690 | MB | Belokranjka | 9.33 ± 0.33 | 5.14 ± 0.07 | 23.23 ± 0.43 | 29.10 ± 0.40 | 52.35 ± 0.21 | 0.30 ± 0.01 |
| 2692 | MR | Belokranjka | 10.00 ± 0.00 | 5.20 ± 0.06 | 24.13 ± 2.70 | 30.90 ± 1.06 | 55.01 ± 2.82 | 0.29 ± 0.03 |
| Control | Belokranjka | 9.67 ± 0.33 | 5.41 ± 0.11 | 25.17 ± 0.59 | 32.80 ± 1.36 | 57.95 ± 1.96 | 0.30 ± 0.02 | |
| 2152 | MR | Belokranjka | 10.00 ± 0.00 | 5.43 ± 0.07 | 28.00 ± 2.11 | 33.47 ± 0.23 | 61.46 ± 2.21 | 0.29 ± 0.01 |
| Control | Belokranjka | 9.33 ± 0.33 | 5.11 ± 0.00 | 28.53 ± 0.87 | 26.87 ± 0.93 | 55.39 ± 0.82 | 0.27 ± 0.01 | |
| 2146 | MR | Belokranjka | 9.33 ± 0.33 | 5.22 ± 0.16 | 26.33 ± 1.87 | 30.00 ± 0.35 | 56.36 ± 1.75 | 0.35 ± 0.02 |
| 2147 | MR | Belokranjka | 10.00 ± 0.00 | 4.90 ± 0.21 | 24.83 ± 1.42 | 26.03 ± 0.43 | 50.84 ± 1.51 | 0.25 ± 0.02 |
| 2251 | MR | Belokranjka | 9.33 ± 0.33 | 5.31 ± 0.22 | 28.33 ± 3.80 | 28.00 ± 0.78 | 56.27 ± 4.42 | 0.27 ± 0.03 |
| 2789 | MR | Belokranjka | 9.67 ± 0.33 | 5.07 ± 0.14 | 27.23 ± 1.95 | 26.10 ± 1.01 | 53.37 ± 2.55 | 0.28 ± 0.02 |
| 2793 | MR | Belokranjka | 9.33 ± 0.67 | 4.93 ± 0.09 | 26.27 ± 2.03 | 27.47 ± 1.83 | 53.71 ± 1.97 | 0.30 ± 0.00 |
| 2794 | MR | Belokranjka | 8.67 ± 0.88 | 4.85 ± 0.03 | 29.03 ± 2.22 | 29.43 ± 0.50 | 58.47 ± 2.68 | 0.35 ± 0.03 |
| 2795 | MR | Belokranjka | 9.67 ± 0.33 | 5.27 ± 0.18 | 24.60 ± 2.01 | 29.10 ± 1.46 | 53.69 ± 0.85 | 0.27 ± 0.01 |
| Control | Belokranjka | 9.67 ± 0.33 | 5.11 ± 0.21 | 19.60 ± 1.91 | 25.23 ± 1.12 | 44.81 ± 2.82 | 0.25 ± 0.01 | |
| 2645 | MR | Belokranjka | 9.67 ± 0.33 | 5.00 ± 0.06 | 23.57 ± 0.52 | 26.23 ± 1.36 | 49.76 ± 1.90 | 0.34 ± 0.06 |
| 2691 | MR | Belokranjka | 9.00 ± 0.00 | 5.26 ± 0.04 | 24.40 ± 3.21 | 25.50 ± 1.19 | 49.91 ± 4.36 | 0.30 ± 0.01 |
| 2693 | MR | Belokranjka | 10.00 ± 0.00 | 5.07 ± 0.09 | 23.93 ± 0.91 | 26.27 ± 0.78 | 50.19 ± 1.61 | 0.32 ± 0.04 |
| 2790 | MR | Belokranjka | 9.67 ± 0.33 | 5.34 ± 0.18 | 21.50 ± 0.49 | 27.37 ± 1.92 | 48.86 ± 2.42 | 0.31 ± 0.02 |
| Control | Belokranjka | 8.33 ± 0.33 | 5.47 ± 0.16 | 20.63 ± 1.77 | 23.40 ± 1.08 | 44.05 ± 2.27 | 0.31 ± 0.03 | |
| 2634 | MR | Belokranjka | 9.33 ± 0.33 | 5.56 ± 0.20 | 21.20 ± 1.10 | 25.40 ± 0.71 | 46.57 ± 1.06 | 0.34 ± 0.06 |
| 2214 | MR | Belokranjka | 9.33 ± 0.33 | 5.25 ± 0.04 | 23.57 ± 0.73 | 27.07 ± 0.87 | 50.63 ± 0.22 | 0.33 ± 0.03 |
| 2243 | MR | Belokranjka | 9.00 ± 0.58 | 5.23 ± 0.07 | 23.90 ± 1.99 | 28.37 ± 1.06 | 52.25 ± 2.88 | 0.36 ± 0.00 |
| 2702 | MR | Belokranjka | 9.00 ± 0.00 | 5.07 ± 0.04 | 22.10 ± 2.80 | 27.07 ± 1.39 | 49.18 ± 3.64 | 0.32 ± 0.00 |
| 2791 | MR | Belokranjka | 9.33 ± 0.33 | 5.10 ± 0.10 | 25.33 ± 4.53 | 27.10 ± 1.61 | 52.42 ± 6.14 | 0.32 ± 0.03 |
| Control | Belokranjka | 9.00 ± 0.58 | 4.91 ± 0.21 | 24.20 ± 0.38 | 28.30 ± 0.51 | 52.52 ± 0.21 | 0.31 ± 0.01 | |
| 2250 | MG | Belokranjka | 8.33 ± 0.88 | 5.13 ± 0.07 | 24.20 ± 2.08 | 26.77 ± 0.42 | 50.98 ± 1.81 | 0.26 ± 0.02 |
| 2685 | MR | Belokranjka | 9.00 ± 0.58 | 4.99 ± 0.11 | 22.50 ± 0.25 | 29.97 ± 1.39 | 52.45 ± 1.59 | 0.30 ± 0.02 |
| Control | Belokranjka | 8.67 ± 0.33 | 5.23 ± 0.17 | 24.10 ± 0.75 | 26.63 ± 0.12 | 50.75 ± 0.67 | 0.29 ± 0.02 | |
| 2640 | MR | Belokranjka | 9.67 ± 0.33 | 5.17 ± 0.09 | 23.37 ± 2.00 | 26.10 ± 0.29 | 49.47 ± 1.89 | 0.28 ± 0.02 |
| 2694 | MR | Belokranjka | 9.67 ± 0.33 | 5.28 ± 0.32 | 24.87 ± 1.07 | 27.90 ± 0.66 | 52.77 ± 1.05 | 0.26 ± 0.00 |
| 2695 | MR | Belokranjka | 9.33 ± 0.33 | 5.11 ± 0.06 | 24.40 ± 2.94 | 26.57 ± 0.90 | 50.96 ± 3.77 | 0.28 ± 0.02 |
| 2788 | MR | Belokranjka | 8.67 ± 0.67 | 5.15 ± 0.27 | 23.87 ± 2.12 | 28.20 ± 1.33 | 52.07 ± 2.48 | 0.32 ± 0.02 |
| 2792 | MR | Belokranjka | 9.33 ± 0.33 | 5.57 ± 0.36 | 25.20 ± 3.54 | 28.10 ± 0.96 | 53.27 ± 4.46 | 0.29 ± 0.01 |
| 2796 | MR | Belokranjka | 8.67 ± 1.33 | 4.98 ± 0.11 | 21.90 ± 1.20 | 27.70 ± 2.06 | 49.62 ± 0.86 | 0.32 ± 0.02 |
| Control | Belokranjka | 8.67 ± 0.88 | 4.67 ± 0.13 | 22.93 ± 1.71 | 28.00 ± 1.13 | 50.96 ± 2.47 | 0.29 ± 0.02 | |
| 2688 | MR | Belokranjka | 9.00 ± 0.58 | 5.10 ± 0.23 | 26.80 ± 0.68 | 28.97 ± 0.37 | 55.76 ± 0.48 | 0.36 ± 0.03 |
| Control | Belokranjka | 8.33 ± 0.67 | 4.52 ± 0.24 | 15.53 ± 1.49 | 21.43 ± 0.88 | 36.97 ± 2.00 | 0.15 ± 0.01 | |
| 2154 | MR | Belokranjka | 7.67 ± 0.33 | 5.09 ± 0.25 | 14.90 ± 0.92 | 22.43 ± 0.78 | 37.35 ± 1.07 | 0.14 ± 0.01 |
| Control | Belokranjka | 9.67 ± 0.33 | 4.14 ± 0.10 | 22.97 ± 0.84 | 26.53 ± 0.55 | 49.54 ± 1.18 | 0.23 ± 0.01 | |
| 2150 | MR | Belokranjka | 10.00 ± 0.00 | 4.47 ± 0.37 | 23.13 ± 1.39 | 27.07 ± 1.20 | 50.23 ± 2.56 | 0.26 ± 0.01 |
| 2240 | MR | Belokranjka | 9.67 ± 0.33 | 4.28 ± 0.09 | 24.07 ± 0.77 | 27.53 ± 1.78 | 51.60 ± 2.41 | 0.25 ± 0.02 |
| Emergence Success [n] | Emergence Time [d] | Root Length [cm] | Shoot Length [cm] | Total Plant Length [cm] | Plant Dry Weight [g] | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate | Taxon | No Fertilizer | Fertilizer | No Fertilizer | Fertilizer | No Fertilizer | Fertilizer | No Fertilizer | Fertilizer | No Fertilizer | Fertilizer | No Fertilizer | Fertilizer |
| Control | 5.00 ± 0.00 | 4.33 ± 0.67 | 4.25 ± 0.00 | 4.42 ± 0.21 | 18.90 ± 0.00 | 16.27 ± 0.58 | 14.10 ± 0.00 | 16.77 ± 1.30 | 32.93 ± 0.00 | 33.05 ± 1.72 | 0.21 ± 0.00 | 0.27 ± 0.02 | |
| 1154 | MB | 5.00 ± 0.00 | 4.67 ± 0.33 | 4.60 ± 0.12 | 4.83 ± 0.34 | 16.23 ± 1.42 | 14.13 ± 0.74 | 12.93 ± 0.72 | 16.40 ± 0.46 | 29.11 ± 1.18 | 30.53 ± 1.00 | 0.25 ± 0.02 | 0.42 ± 0.04 |
| 1868 | MB | 4.67 ± 0.33 | 5.00 ± 0.00 | 4.33 ± 0.29 | 4.53 ± 0.07 | 17.90 ± 0.70 | 16.73 ± 0.72 | 13.50 ± 0.06 | 16.77 ± 0.81 | 31.38 ± 0.77 | 33.49 ± 0.40 | 0.24 ± 0.01 | 0.38 ± 0.05 |
| 2121 | BB | 5.00 ± 0.00 | 4.67 ± 0.33 | 4.72 ± 0.17 | 5.02 ± 0.13 | 15.53 ± 0.87 | 15.27 ± 0.23 | 15.07 ± 0.74 | 18.40 ± 0.59 | 30.65 ± 1.24 | 33.68 ± 0.76 | 0.30 ± 0.00 | 0.41 ± 0.09 |
| Control | 5.00 ± 0.00 | 4.67 ± 0.33 | 4.67 ± 0.13 | 5.17 ± 0.20 | 14.63 ± 0.90 | 15.43 ± 0.92 | 13.40 ± 2.15 | 18.70 ± 1.36 | 28.04 ± 2.92 | 34.15 ± 2.29 | 0.28 ± 0.03 | 0.30 ± 0.02 | |
| 2631 | MR | 4.67 ± 0.33 | 5.00 ± 0.00 | 4.63 ± 0.09 | 4.40 ± 0.12 | 20.03 ± 0.96 | 18.77 ± 0.19 | 13.80 ± 0.65 | 17.47 ± 0.50 | 33.84 ± 1.20 | 36.23 ± 0.57 | 0.30 ± 0.01 | 0.31 ± 0.04 |
| 2632 | MR | 4.33 ± 0.67 | 5.00 ± 0.00 | 4.42 ± 0.14 | 4.53 ± 0.07 | 17.47 ± 0.99 | 19.67 ± 1.62 | 12.87 ± 0.43 | 17.50 ± 0.86 | 30.38 ± 1.32 | 37.17 ± 1.98 | 0.30 ± 0.03 | 0.27 ± 0.03 |
| 2245 | MR | 4.67 ± 0.33 | 5.00 ± 0.00 | 5.37 ± 0.63 | 4.53 ± 0.24 | 18.63 ± 0.58 | 19.33 ± 1.43 | 14.07 ± 0.73 | 17.83 ± 0.47 | 32.72 ± 0.99 | 37.16 ± 1.35 | 0.23 ± 0.02 | 0.27 ± 0.02 |
| 2246 | MR | 5.00 ± 0.00 | 4.67 ± 0.33 | 4.67 ± 0.24 | 5.15 ± 0.18 | 18.13 ± 0.64 | 19.33 ± 1.83 | 12.37 ± 0.37 | 18.67 ± 0.76 | 30.49 ± 1.00 | 37.99 ± 2.43 | 0.26 ± 0.03 | 0.30 ± 0.02 |
| Control | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.53 ± 0.37 | 5.47 ± 0.29 | 18.93 ± 0.92 | 17.95 ± 1.23 | 13.34 ± 1.26 | 18.80 ± 0.96 | 32.27 ± 1.92 | 36.75 ± 1.70 | 0.20 ± 0.02 | 0.23 ± 0.02 | |
| 2215 | MR | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.87 ± 0.35 | 5.80 ± 0.12 | 17.15 ± 1.35 | 19.25 ± 1.35 | 13.19 ± 1.10 | 17.18 ± 0.33 | 30.33 ± 2.06 | 36.43 ± 1.40 | 0.19 ± 0.00 | 0.21 ± 0.01 |
| 2216 | MR | 4.67 ± 0.33 | 4.00 ± 0.58 | 5.68 ± 0.28 | 5.27 ± 0.27 | 18.92 ± 1.85 | 17.62 ± 1.43 | 13.50 ± 0.14 | 16.97 ± 0.97 | 32.42 ± 1.85 | 34.59 ± 1.36 | 0.25 ± 0.01 | 0.21 ± 0.01 |
| 2299 | BB | 4.67 ± 0.33 | 5.00 ± 0.00 | 5.93 ± 0.58 | 5.87 ± 0.29 | 12.94 ± 1.10 | 15.99 ± 1.95 | 14.98 ± 0.26 | 18.75 ± 0.36 | 27.92 ± 1.27 | 34.73 ± 1.60 | 0.18 ± 0.01 | 0.19 ± 0.01 |
| 2300 | BB | 4.67 ± 0.33 | 4.67 ± 0.33 | 5.18 ± 0.32 | 5.98 ± 0.21 | 19.94 ± 1.20 | 14.48 ± 0.74 | 15.33 ± 0.42 | 19.26 ± 0.22 | 35.26 ± 1.58 | 33.74 ± 0.58 | 0.23 ± 0.00 | 0.25 ± 0.02 |
| Control | 5.00 ± 0.00 | 4.33 ± 0.67 | 5.27 ± 0.29 | 5.22 ± 0.51 | 19.67 ± 1.56 | 17.50 ± 1.53 | 13.97 ± 0.58 | 18.40 ± 0.57 | 33.63 ± 0.98 | 35.90 ± 1.97 | 0.19 ± 0.01 | 0.20 ± 0.02 | |
| 2635 | MR | 4.67 ± 0.33 | 4.67 ± 0.33 | 5.97 ± 0.27 | 6.13 ± 0.77 | 18.20 ± 0.90 | 16.30 ± 1.40 | 13.47 ± 1.22 | 19.30 ± 1.05 | 31.68 ± 1.18 | 35.62 ± 2.23 | 0.18 ± 0.01 | 0.21 ± 0.00 |
| 2637 | MR | 4.33 ± 0.33 | 4.67 ± 0.33 | 5.68 ± 0.09 | 5.65 ± 0.13 | 17.00 ± 0.75 | 19.60 ± 1.04 | 14.23 ± 0.38 | 19.63 ± 0.26 | 31.25 ± 1.14 | 39.23 ± 1.25 | 0.17 ± 0.00 | 0.22 ± 0.03 |
| 2641 | MR | 5.00 ± 0.00 | 5.00 ± 0.00 | 4.93 ± 0.07 | 5.07 ± 0.13 | 17.40 ± 0.61 | 18.73 ± 2.03 | 14.03 ± 0.42 | 20.07 ± 0.61 | 31.42 ± 0.61 | 38.77 ± 2.58 | 0.20 ± 0.01 | 0.21 ± 0.01 |
| Control | 5.00 ± 0.00 | 4.67 ± 0.33 | 4.93 ± 0.07 | 5.07 ± 0.07 | 19.73 ± 1.36 | 18.27 ± 1.92 | 15.90 ± 0.23 | 19.33 ± 0.44 | 35.61 ± 1.28 | 37.59 ± 2.35 | 0.42 ± 0.04 | 0.41 ± 0.02 | |
| 2697 | ND | 5.00 ± 0.00 | 5.00 ± 0.00 | 4.40 ± 0.12 | 4.87 ± 0.24 | 19.03 ± 1.68 | 16.77 ± 1.93 | 14.30 ± 1.21 | 20.00 ± 0.96 | 33.34 ± 2.80 | 36.75 ± 2.64 | 0.40 ± 0.03 | 0.46 ± 0.04 |
| 2698 | MR | 4.67 ± 0.33 | 5.00 ± 0.00 | 4.80 ± 0.12 | 5.20 ± 0.40 | 17.10 ± 1.12 | 16.50 ± 1.65 | 15.10 ± 0.45 | 18.17 ± 0.90 | 32.20 ± 1.39 | 34.69 ± 2.31 | 0.37 ± 0.03 | 0.45 ± 0.04 |
| 2699 | MR | 4.67 ± 0.33 | 5.00 ± 0.00 | 5.42 ± 0.42 | 5.20 ± 0.23 | 19.87 ± 2.63 | 21.47 ± 2.86 | 14.67 ± 1.28 | 21.13 ± 0.78 | 34.51 ± 3.89 | 42.59 ± 2.23 | 0.37 ± 0.04 | 0.36 ± 0.02 |
| 2700 | MR | 4.67 ± 0.33 | 5.00 ± 0.00 | 5.08 ± 0.08 | 4.67 ± 0.24 | 19.43 ± 1.93 | 20.33 ± 0.83 | 15.33 ± 0.41 | 19.27 ± 0.48 | 34.73 ± 2.28 | 39.63 ± 0.99 | 0.34 ± 0.03 | 0.39 ± 0.01 |
| Control | 5.00 ± 0.00 | 4.67 ± 0.33 | 5.33 ± 0.35 | 5.50 ± 0.06 | 21.07 ± 1.69 | 19.00 ± 1.30 | 15.67 ± 1.17 | 21.63 ± 0.85 | 36.75 ± 2.84 | 40.65 ± 0.84 | 0.39 ± 0.02 | 0.28 ± 0.07 | |
| 2298 | BB | 4.67 ± 0.33 | 4.67 ± 0.33 | 5.57 ± 0.12 | 4.92 ± 0.14 | 21.03 ± 1.07 | 22.33 ± 1.32 | 16.60 ± 0.61 | 21.63 ± 0.81 | 37.63 ± 1.58 | 43.97 ± 1.46 | 0.45 ± 0.03 | 0.44 ± 0.03 |
| 2243 | MR | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.40 ± 0.23 | 5.20 ± 0.23 | 20.80 ± 1.22 | 21.27 ± 0.89 | 16.10 ± 0.30 | 21.27 ± 0.72 | 36.89 ± 1.52 | 42.51 ± 1.38 | 0.39 ± 0.04 | 0.39 ± 0.08 |
| 2636 | MR | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.60 ± 0.35 | 5.13 ± 0.24 | 19.90 ± 1.35 | 20.87 ± 1.01 | 14.83 ± 1.03 | 20.20 ± 1.54 | 34.77 ± 2.35 | 41.04 ± 2.56 | 0.33 ± 0.04 | 0.37 ± 0.03 |
| 2642 | MR | 5.00 ± 0.00 | 4.67 ± 0.33 | 5.60 ± 0.12 | 5.20 ± 0.12 | 19.57 ± 1.12 | 21.07 ± 1.16 | 13.27 ± 1.03 | 18.93 ± 1.13 | 32.83 ± 2.10 | 39.99 ± 2.30 | 0.36 ± 0.01 | 0.32 ± 0.08 |
| Control | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.33 ± 0.47 | 4.87 ± 0.07 | 21.87 ± 0.84 | 18.77 ± 1.53 | 16.97 ± 0.38 | 21.03 ± 1.65 | 38.89 ± 1.14 | 39.79 ± 1.96 | 0.19 ± 0.01 | 0.21 ± 0.01 | |
| 2148 | MR | 5.00 ± 0.00 | 5.00 ± 0.00 | 4.40 ± 0.23 | 4.87 ± 0.07 | 17.67 ± 1.02 | 18.20 ± 1.51 | 14.63 ± 1.04 | 19.53 ± 1.02 | 32.28 ± 1.21 | 37.74 ± 1.92 | 0.20 ± 0.01 | 0.24 ± 0.01 |
| 2151 | MR | 4.67 ± 0.33 | 4.67 ± 0.33 | 4.83 ± 0.28 | 4.83 ± 0.34 | 20.80 ± 2.21 | 20.07 ± 0.70 | 15.87 ± 0.34 | 22.20 ± 0.42 | 36.65 ± 2.56 | 42.27 ± 0.38 | 0.21 ± 0.02 | 0.21 ± 0.00 |
| 2152 | MR | 4.33 ± 0.33 | 5.00 ± 0.00 | 5.15 ± 0.69 | 4.87 ± 0.13 | 20.50 ± 0.85 | 18.90 ± 0.87 | 16.07 ± 1.55 | 21.40 ± 0.85 | 36.59 ± 1.96 | 40.31 ± 0.08 | 0.19 ± 0.00 | 0.21 ± 0.02 |
| 2701 | MR | 5.00 ± 0.00 | 4.33 ± 0.33 | 4.80 ± 0.12 | 5.43 ± 0.70 | 20.90 ± 0.78 | 18.40 ± 0.87 | 15.50 ± 0.95 | 21.33 ± 0.38 | 36.37 ± 1.19 | 39.72 ± 1.23 | 0.20 ± 0.01 | 0.23 ± 0.00 |
| Control | 4.33 ± 0.33 | 5.00 ± 0.00 | 4.90 ± 0.21 | 4.73 ± 0.24 | 19.47 ± 2.25 | 19.13 ± 0.58 | 14.83 ± 1.39 | 20.30 ± 1.30 | 34.27 ± 3.62 | 39.43 ± 1.83 | 0.21 ± 0.01 | 0.22 ± 0.00 | |
| 2703 | MB | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.53 ± 0.27 | 5.87 ± 0.64 | 19.10 ± 1.66 | 21.13 ± 0.52 | 15.90 ± 0.32 | 18.40 ± 0.35 | 34.95 ± 1.35 | 39.51 ± 0.65 | 0.20 ± 0.00 | 0.23 ± 0.01 |
| 2009 | MR | 5.00 ± 0.00 | 3.67 ± 0.67 | 6.00 ± 0.53 | 5.87 ± 0.59 | 17.57 ± 0.62 | 17.97 ± 1.08 | 15.13 ± 1.82 | 22.87 ± 1.67 | 32.71 ± 2.32 | 40.83 ± 2.67 | 0.21 ± 0.02 | 0.23 ± 0.01 |
| 2010 | MG | 5.00 ± 0.00 | 5.00 ± 0.00 | 7.40 ± 0.90 | 6.20 ± 0.35 | 19.80 ± 1.59 | 16.37 ± 0.47 | 15.40 ± 1.56 | 21.93 ± 1.03 | 35.21 ± 0.57 | 38.30 ± 1.49 | 0.21 ± 0.00 | 0.21 ± 0.02 |
| 2011 | MR | 4.33 ± 0.33 | 4.67 ± 0.33 | 5.22 ± 0.46 | 5.60 ± 0.35 | 19.53 ± 0.78 | 19.40 ± 0.06 | 19.13 ± 0.42 | 22.97 ± 0.60 | 38.66 ± 1.16 | 42.40 ± 0.54 | 0.22 ± 0.02 | 0.23 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Praprotnik, E.; Lončar, J.; Razinger, J. Testing Virulence of Different Species of Insect Associated Fungi against Yellow Mealworm (Coleoptera: Tenebrionidae) and Their Potential Growth Stimulation to Maize. Plants 2021, 10, 2498. https://doi.org/10.3390/plants10112498
Praprotnik E, Lončar J, Razinger J. Testing Virulence of Different Species of Insect Associated Fungi against Yellow Mealworm (Coleoptera: Tenebrionidae) and Their Potential Growth Stimulation to Maize. Plants. 2021; 10(11):2498. https://doi.org/10.3390/plants10112498
Chicago/Turabian StylePraprotnik, Eva, Jernej Lončar, and Jaka Razinger. 2021. "Testing Virulence of Different Species of Insect Associated Fungi against Yellow Mealworm (Coleoptera: Tenebrionidae) and Their Potential Growth Stimulation to Maize" Plants 10, no. 11: 2498. https://doi.org/10.3390/plants10112498