
In Vitro Anti-Epstein Barr Virus Activity of Olea europaea L. Leaf Extracts


 and
and
Abstract
:1. Introduction
2. Results
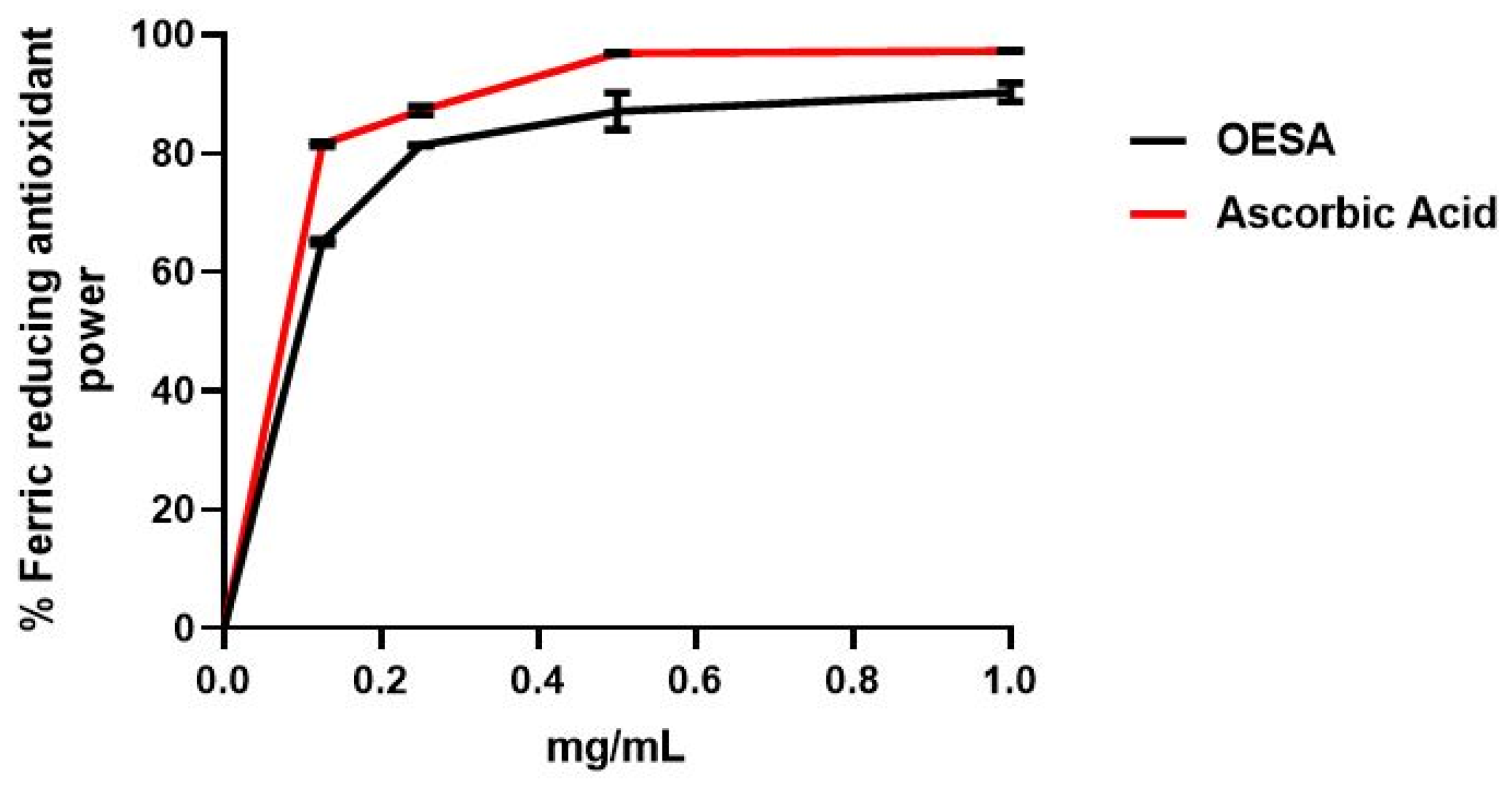
2.1. Antioxidant Capacity of OESA
2.2. MTT Cell Proliferation Assay
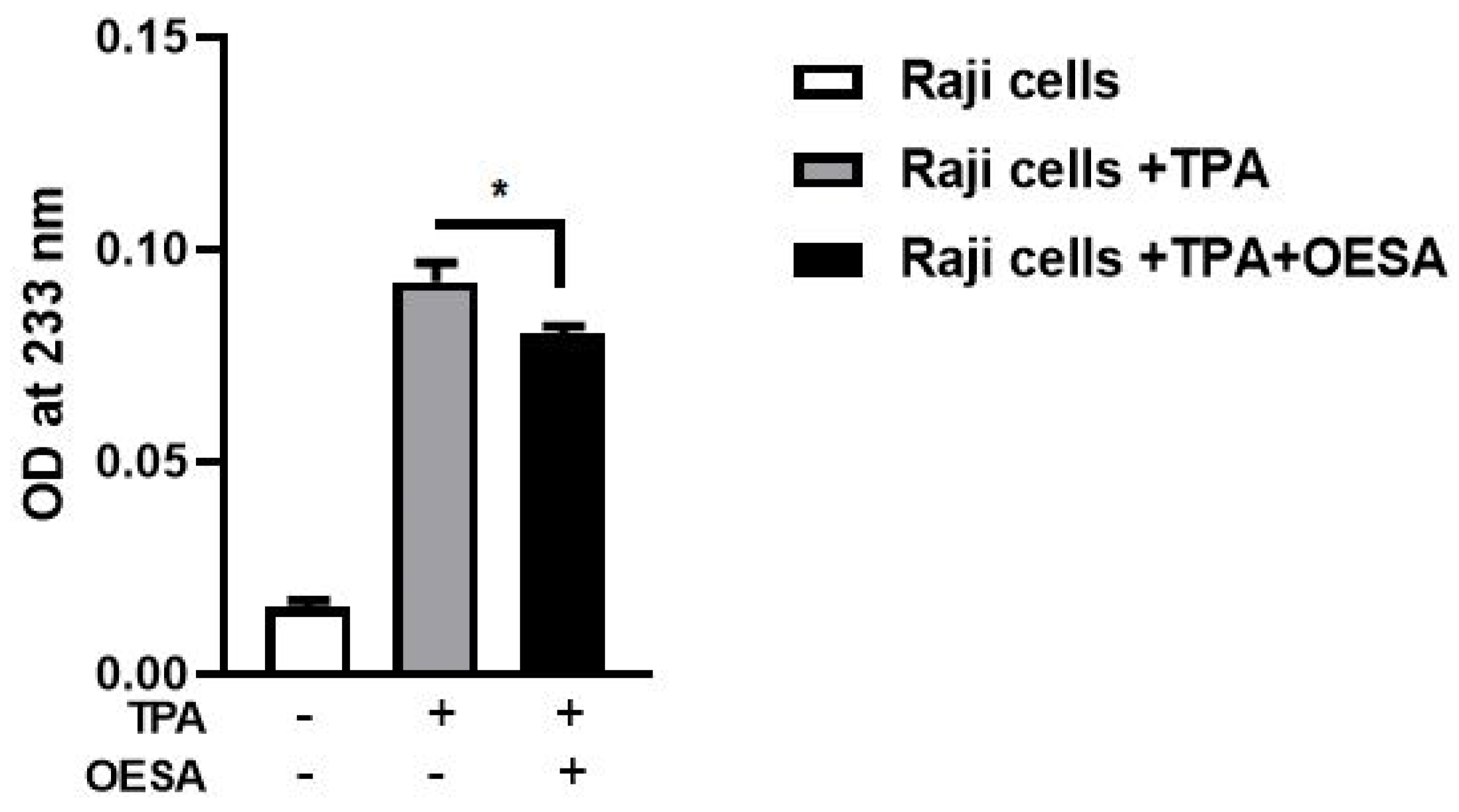
2.3. Evaluation of Lipid Peroxidation: MDA and DC Determination
2.4. Detection of Inhibitory Activity on the EBV Activation Mediated by OESA
3. Discussion
4. Materials and Methods
4.1. Olive Leaf Extracts Preparation
4.2. 2,2-Diphenyl- 1-Picrylhydrazyl (DPPH) Free Radical Scavenging Activity Assay
4.3. Ferric Reducing Antioxidant Power (FRAP) Assay
4.4. Raji Cell Culture
4.5. MTT Cell Proliferation Assay
4.6. EBV Antigens Induction in Raji Cells
4.7. Malondialdehyde (MDA) Determination
4.8. Conjugated Dienes (CD) Determination
4.9. Indirect Immunofluorescence Assay (IFA)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Ghasemian, M.; Owlia, S.; Owlia, M.B. Review of Anti-Inflammatory Herbal Medicines. Adv. Pharmacol. Sci. 2016, 2016, 9130979. [Google Scholar] [CrossRef] [Green Version]
- Yahia, Y.; Benabderrahim, M.A.; Tlili, N.; Bagues, M.; Nagaz, K. Bioactive compounds, antioxidant and antimicrobial activities of extracts from different plant parts of two Ziziphus Mill. species. PLoS ONE 2020, 15, e0232599. [Google Scholar] [CrossRef]
- Kapoor, R.; Sharma, B.; Kanwar, S.S. Antiviral Phytochemicals: An Overview. Biochem. Physiol. 2017, 6, 220. [Google Scholar] [CrossRef]
- Ghildiyal, R.; Prakash, V.; Chaudhary, V.K.; Gupta, V.; Gabrani, R. Phytochemicals as Antiviral Agents: Recent Updates. Plant-Deriv. Bioact. 2020, 279–295. [Google Scholar] [CrossRef]
- Ebob, O.T.; Babiaka, S.B.; Ntie-Kang, F. Natural products as potential lead compounds for drug discovery against SARS-CoV-2. Nat. Prod. Bioprospect. 2021, 1–18. [Google Scholar] [CrossRef]
- Omrani, M.; Keshavarz, M.; Ebrahimi, S.N.; Mehrabi, M.; McGaw, L.J.; Abdalla, M.A.; Mehrbod, P. Potential Natural Products Against Respiratory Viruses: A Perspective to Develop Anti-COVID-19 Medicines. Front. Pharmacol. 2021, 11, 586993. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Twilley, D.; Esmear, T.; Oosthuizen, C.B.; Reid, A.M.; Nel, M.; Lall, N. Anti-SARS-CoV natural products with the potential to inhibit SARS-CoV-2 (COVID-19). Front. Pharmacol. 2020, 11, 561334. [Google Scholar] [CrossRef] [PubMed]
- Musarra-Pizzo, M.; Pennisi, R.; Ben-Amor, I.; Mandalari, G.; Sciortino, M. Antiviral Activity Exerted by Natural Products against Human Viruses. Viruses 2021, 13, 828. [Google Scholar] [CrossRef] [PubMed]
- Musarra-Pizzo, M.; Pennisi, R.; Ben-Amor, I.; Smeriglio, A.; Mandalari, G.; Sciortino, M.T. In Vitro Anti-HSV-1 Activity of Polyphenol-Rich Extracts and Pure Polyphenol Compounds Derived from Pistachios Kernels (Pistacia vera L.). Plants 2020, 9, 267. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.; Taha, M.E.; Makeen, H.; Alhazmi, H.; Al Bratty, M.; Sultana, S.; Ahsan, W.; Najmi, A.; Khalid, A. Bioactive Natural Antivirals: An Updated Review of the Available Plants and Isolated Molecules. Molecules 2020, 25, 4878. [Google Scholar] [CrossRef]
- Thomas, E.; Stewart, L.E.; Darley, B.A.; Pham, A.M.; Esteban, I.; Panda, S.S. Plant-Based Natural Products and Extracts: Potential Source to Develop New Antiviral Drug Candidates. Molecules 2021, 26, 6197. [Google Scholar] [CrossRef]
- Astin, J.A.; Marie, A.; Pelletier, K.R.; Hansen, E.; Haskell, W.L. A review of the incorporation of complementary and alternative medicine by mainstream physicians. Arch. Int. Med. 1998, 158, 2303–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somova, L.; Shode, F.; Ramnanan, P.; Nadar, A. Antihypertensive, antiatherosclerotic and antioxidant activity of triterpenoids isolated from Olea europaea, subspecies africana leaves. J. Ethnopharmacol. 2003, 84, 299–305. [Google Scholar] [CrossRef]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, M.; Knez, Z. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Ben-Amor, I.; Musarra-Pizzo, M.; Smeriglio, A.; D’Arrigo, M.; Pennisi, R.; Attia, H.; Gargouri, B.; Trombetta, D.; Mandalari, G.; Sciortino, M. Phytochemical Characterization of Olea europea Leaf Extracts and Assessment of Their Anti-Microbial and Anti-HSV-1 Activity. Viruses 2021, 13, 1085. [Google Scholar] [CrossRef] [PubMed]
- Haris, O.S. Oleuropein in olive and its pharmacological effects. Sci. Pharm. 2010, 78, 133–154. [Google Scholar] [CrossRef] [Green Version]
- Toussirot, E.; Roudier, J. Pathophysiological links between rheumatoid arthritis and the Epstein–Barr virus: An update. Jt. Bone Spine 2007, 74, 418–426. [Google Scholar] [CrossRef]
- Sarban, S.; Kocyigit, A.; Yazar, M.; Isikan, U.E. Plasma total antioxidant capacity, lipid peroxidation, and erythrocyte antioxidant enzyme activities in patients with rheumatoid arthritis and osteoarthritis. Clin. Biochem. 2005, 38, 981–986. [Google Scholar] [CrossRef]
- Gargouri, B.; Van Pelt, J.; Elfeki, A.; Attia, H.; Lassoued, S. Induction of Epstein Barr Virus (EBV) lytic cycle in vitro causes oxidative stress in lymphoblastoïde B cell lines. Mol. Cell Biochem. 2009, 324, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, B.; Nasr, R.; Ben Mansour, R.; Lassoued, S.; Mseddi, M.; Attia, H.; Feki, A.E.F.E.; Van Pelt, J. Reactive Oxygen Species Production and Antioxidant Enzyme Expression after Epstein–Barr Virus Lytic Cycle Induction in Raji Cell Line. Biol. Trace Elem. Res. 2011, 144, 1449–1457. [Google Scholar] [CrossRef]
- Lassoued, S.; Ben Ameur, R.; Ayadi, W.; Gargouri, B.; Ben Mansour, R.; Attia, H. Epstein-Barr virus induces an oxidative stress during the early stages of infection in B lymphocytes, epithelial, and lymphoblastoid cell lines. Mol. Cell. Biochem. 2008, 313, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Briante, R.; Patumi, M.; Terenziani, S.; Bismuto, E.; Febbraio, F.; Nucci, R. Olea europaea L. Leaf Extract and Derivatives: Antioxidant Properties. J. Agric. Food Chem. 2002, 50, 4934–4940. [Google Scholar] [CrossRef]
- Kurien, B.T.; Scofield, R. Free radical mediated peroxidative damage in systemic lupus erythematosus. Life Sci. 2003, 73, 1655–1666. [Google Scholar] [CrossRef]
- Romero, M.; Toral, M.; Gómez-Guzmán, M.; Jiménez, R.; Galindo, P.; Sánchez, M.; Olivares, M.; Gálvez, J.; Duarte, J. Antihypertensive effects of oleuropein-enriched olive leaf extract in spontaneously hypertensive rats. Food Funct. 2016, 7, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Fredrickson, W.R.; F&S Group, Inc. Method and Composition for Antiviral Therapy with Olive Leaves. U.S. Patent 6,117,844, 12 September 2000. [Google Scholar]
- Gülçin, I. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total Antioxidant Capacity of Plant Foods, Beverages and Oils Consumed in Italy Assessed by Three Different In Vitro Assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [Green Version]
- Neimkhum, W.; Anuchapreeda, S.; Lin, W.C.; Lue, S.C.; Lee, K.H.; Chaiyana, W. Effects of Carissa carandas Linn. Fruit, Pulp, Leaf, and Seed on Oxidation, Inflammation, Tyro-sinase, Matrix Metalloproteinase, Elastase, and Hyaluronidase Inhibition. Antioxidants 2021, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Abaj, F.; Rafiee, M.; Koohdani, F. Interaction between dietary total antioxidant capacity and BDNF Val66Met polymorphism on lipid profiles and atherogenic indices among diabetic patients. Sci. Rep. 2021, 11, 19108. [Google Scholar] [CrossRef]
- Bouaziz, M.; Chamkha, A.M.; Sayadi, S. Comparative Study on Phenolic Content and Antioxidant Activity during Maturation of the Olive Cultivar Chemlali from Tunisia. J. Agric. Food Chem. 2004, 52, 5476–5481. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Gerothanassis, I.P. Phenolic compounds and antioxidant activity of olive leaf extracts. Nat. Prod. Res. 2012, 26, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Čabarkapa, A.; Dekanski, D.; Živković, L.; Milanović-Čabarkapa, M.; Bajić, V.; Topalović, D.; Giampieri, F.; Gasparrini, M.; Battino, M.; Spremo-Potparević, B. Unexpected effect of dry olive leaf extract on the level of DNA damage in lymphocytes of lead intoxicated workers, before and after CaNa 2 EDTA chelation therapy. Food Chem. Toxicol. 2017, 106, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Topalović, D.; Dekanski, D.; Spremo-Potparević, B.; Pirković, A.; Borozan, S.; Bajić, V.; Stojanović, D.; Giampieri, F.; Gasparrini, M.; Živković, L. Dry olive leaf extract attenuates DNA damage induced by estradiol and diethylstilbestrol in human peripheral blood cells in vitro. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 845, 402993. [Google Scholar] [CrossRef]
- Yang, M.D.; Ouyang, A.M. Antioxidant activity from Olea leaf extract depended on seasonal variations and chromatography treatment. Int. J. Org. Chem. 2012, 2, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Svetla, Y.; Petros, M.; Lidiya, G. Polyphenol profile and antioxidant activity of extracts from olive leaves. J. Cent. Eur. Agric. 2016, 17, 154–163. [Google Scholar]
- Lafka, I.T.; Lazou, E.A.; Sinanoglou, J.V.; Lazos, E. Phenolic extracts from wild olive leaves and their potential as edible oils antioxidants. Foods 2013, 2, 18–31. [Google Scholar] [CrossRef]
- Lo, A.K.F.; Lo, K.W.; Tsao, S.W.; Wong, H.L.; Hui, J.W.Y.; To, K.F.; Hayward, S.D.; Chui, Y.L.; Lau, Y.L.; Takada, K.; et al. Epstein-Barr Virus Infection Alters Cellular Signal Cascades in Human Nasopharyngeal Epithelial Cells. Neoplasia 2006, 8, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Dekanski, D.; Ristić, S.; Mitrović, D.M. Antioxidant effect of dry olive (Olea europaea L.) leaf extract on ethanol-induced gastric lesions in rats. Mediterr. J. Nutr. Metab. 2009, 2, 205–211. [Google Scholar] [CrossRef]
- Zahra, K. In vitro studies on the antiviral effect of olive leaf against infectious laryngotracheitis. Glob. Vet. 2007, 1, 24–30. [Google Scholar]
- Micol, V.; Caturla, N.; Perez, F.L.; Mas, V.; Perez, L.; Estepa, A. The olive leaf extract exhibits antiviral activity against viral haemorrhagic septicaemia rhabdovirus (VHSV). Antivir. Res. 2005, 66, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lee-Hung, S.; Zhang, L.; Huang, P.L.; Chang, Y.; Huang, P.L. Anti-HIV activity of olive leaf extract (OLE) and modulation of host cell gene expression by HIV-1 infection and OLE treatment. Biochem. Biophys. Res. 2003, 307, 1029. [Google Scholar] [CrossRef]
- Motamedifar, M.; Nekooeian, A.A.; Moatari, A. The effect of hydroalcoholic extract of olive leaves against herpes simplex virus type 1. Iran. J. Med. Sci. 2007, 32, 222–227. [Google Scholar]
- Niedobitek, G.; Agathanggelou, A.; Herbst, H. Epstein- Barr virus (EBV) infection in infectious mononucleosis: Virus latency, replication and phenotype of EBV-infected cells. J. Pathol. 1997, 182, 151–159. [Google Scholar] [CrossRef]
- Dalpke, A.H.; Thomssen, R.; Ritter, K. Oxidative injury to endothelial cells due to Epstein-Barr virus-induced autoantibodies against manganese superoxide dismutase. J. Med. Virol. 2003, 71, 408–416. [Google Scholar] [CrossRef]
- Boughariou, E.; Allouche, N.; Jmal, I.; Mokadem, N.; Ayed, B.; Hajji, S.; Khanfir, H.; Bouri, S. Modeling aquifer behaviour under climate change and high consumption: Case study of the Sfax region, southeast Tunisia. J. Afr. Earth Sci. 2018, 141, 118–129. [Google Scholar] [CrossRef]
- Osawa, T.; Namiki, M. A novel type of antioxidant isolated from leaf wax of Eucalyptus leaves. Agric. Biol. Chem. 1981, 45, 735–739. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction. Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.-M.; Oh, J.-M.; Choi, S.-C.; Kim, S.-W.; Han, W.-C.; Kim, T.-H.; Park, D.-S.; Jun, C.-D. An efficient method for the rapid establishment of Epstein-Barr virus immortalization of human B lymphocytes. Cell Prolif. 2003, 36, 191–197. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ASSAY | IC50 OESA | IC50 BHT | IC50 Ascorbic Acid |
|---|---|---|---|
| DPPH | 0.1218 mg/mL | 0.09 mg/mL | - |
| FRAP | 0.05737 mg/mL | - | 0.03068 mg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Amor, I.; Gargouri, B.; Attia, H.; Tlili, K.; Kallel, I.; Musarra-Pizzo, M.; Sciortino, M.T.; Pennisi, R. In Vitro Anti-Epstein Barr Virus Activity of Olea europaea L. Leaf Extracts. Plants 2021, 10, 2445. https://doi.org/10.3390/plants10112445
Ben-Amor I, Gargouri B, Attia H, Tlili K, Kallel I, Musarra-Pizzo M, Sciortino MT, Pennisi R. In Vitro Anti-Epstein Barr Virus Activity of Olea europaea L. Leaf Extracts. Plants. 2021; 10(11):2445. https://doi.org/10.3390/plants10112445
Chicago/Turabian StyleBen-Amor, Ichrak, Bochra Gargouri, Hamadi Attia, Khaoula Tlili, Imen Kallel, Maria Musarra-Pizzo, Maria Teresa Sciortino, and Rosamaria Pennisi. 2021. "In Vitro Anti-Epstein Barr Virus Activity of Olea europaea L. Leaf Extracts" Plants 10, no. 11: 2445. https://doi.org/10.3390/plants10112445