
Development and Adoption of Genetically Engineered Plants for Virus Resistance: Advances, Opportunities and Challenges


Abstract
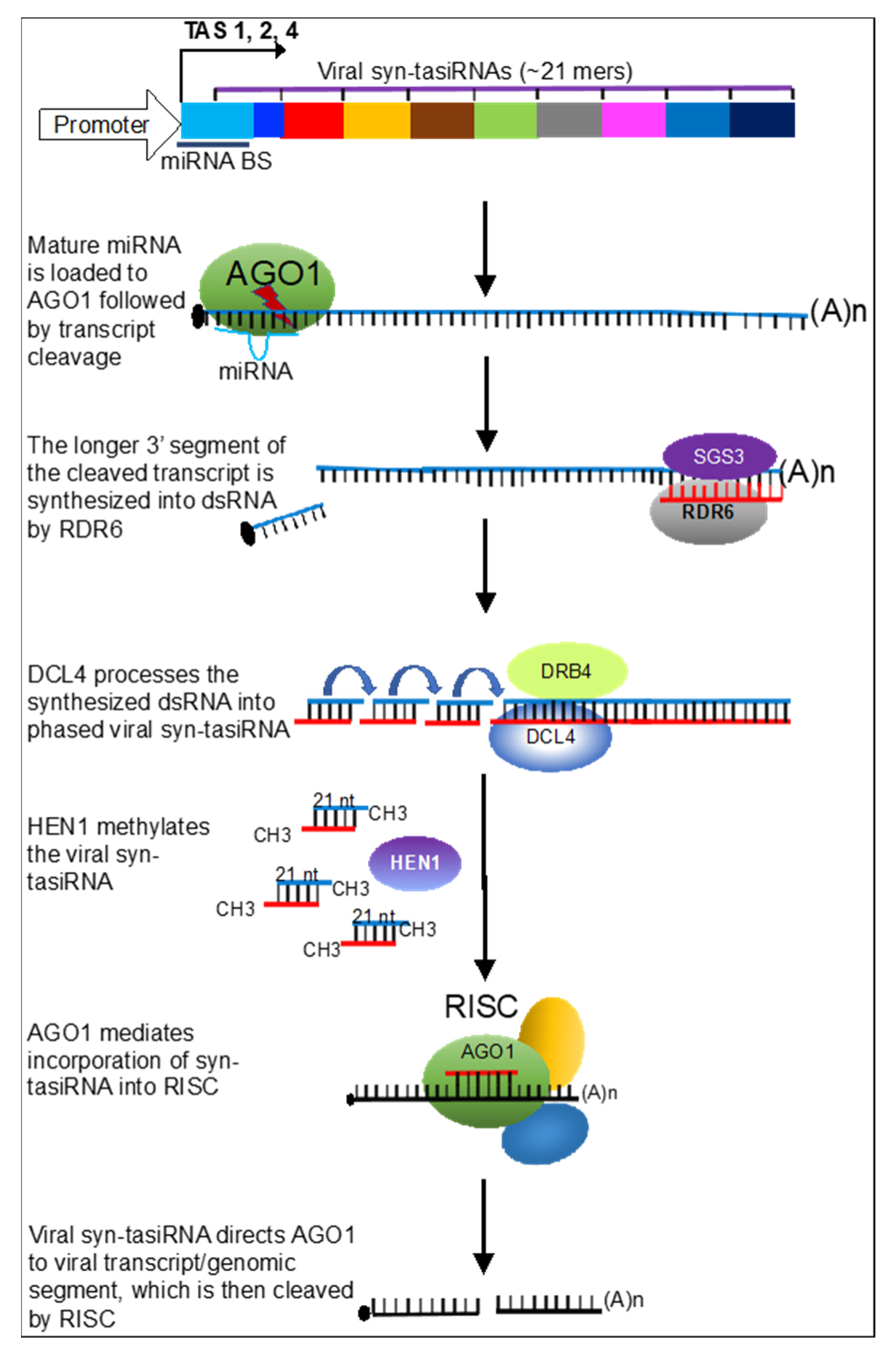
:1. Introduction
2. Functional Genomics and Genome Editing in Engineering Crops for Virus Resistance
3. Benefits of Genetic Engineering in Plant Virus Control
3.1. Short Lead Time
3.2. Comparatively Low Production Cost
3.3. Efficacy and Durability of GE-Mediated Virus Resistance
3.4. Transfer of Virus Resistance Genes Is Not Limited to Closely Related Species
3.5. No Major Effect on the Genetic Makeup of Cultivar
3.6. Clonally Propagated Crops
4. Potential Risks Associated with GM Crops
4.1. Horizontal Gene Transfer and Dissemination in the Environment
4.2. Genetic Erosion
4.3. Recombination of Transgene with Infecting Viruses
4.4. Toxicity of Transgene
5. Regulation of Genetically Modified Crops
6. Some Success Cases of Virus-Resistant GM Crops So Far Approved for Cultivation
6.1. Papaya (Resistance to PRSV Resistant)
6.2. Potato (Resistance to PVY and PLRV)
6.3. Bean (Resistance to BGMV)
6.4. Squash (Resistance to ZYMV, WMV and CMV)
6.5. Cassava (Resistance to CBSD)
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patil, B.L. Plant viral diseases: Economic implications. In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Oxford, UK, 2021; pp. 81–97. ISBN 978-0-12-814516-6. [Google Scholar]
- van Esse, H.P.; Reuber, T.L.; van der Does, D. Genetic Modification to Improve Disease Resistance in Crops. New Phytol. 2020, 225, 70–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Bevan, M.W.; Flavell, R.B.; Chilton, M.D. A Chimaeric Antibiotic Resistance Gene as a Selectable Marker for Plant Cell Transformation. 1983. Biotechnology 1992, 24, 367–370. [Google Scholar]
- Fraley, R.T.; Rogers, S.G.; Horsch, R.B.; Sanders, P.R.; Flick, J.S.; Adams, S.P.; Bittner, M.L.; Brand, L.A.; Fink, C.L.; Fry, J.S.; et al. Expression of Bacterial Genes in Plant Cells. Proc. Natl. Acad. Sci. USA 1983, 80, 4803–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Estrella, L.; Block, M.D.; Messens, E.; Hernalsteens, J.P.; Montagu, M.V.; Schell, J. Chimeric Genes as Dominant Selectable Markers in Plant Cells. EMBO J. 1983, 2, 987–995. [Google Scholar] [CrossRef]
- Lobato-Gómez, M.; Hewitt, S.; Capell, T.; Christou, P.; Dhingra, A.; Girón-Calva, P.S. Transgenic and Genome-Edited Fruits: Background, Constraints, Benefits, and Commercial Opportunities. Hortic. Res. 2021, 8, 1–16. [Google Scholar] [CrossRef]
- James, C. Global Status of Commercialized Biotech/GM Crops: 2005; International Service for the Acquisition of Agri-Biotech Applications: Ithaca, NY, USA, 2005. [Google Scholar]
- Huesing, J.E.; Andres, D.; Braverman, M.P.; Burns, A.; Felsot, A.S.; Harrigan, G.G.; Hellmich, R.L.; Reynolds, A.; Shelton, A.M.; van Rijssen, W.J.; et al. Global Adoption of Genetically Modified (GM) Crops: Challenges for the Public Sector. J. Agric. Food Chem. 2016, 64, 394–402. [Google Scholar] [CrossRef] [Green Version]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2018: Biotech Crops Continue to Help Meet the Challenges of Increased Population and Climate Change; ISAAA: Ithaca, NY, USA, 2018. [Google Scholar]
- ISAAA Database, 2019 ISAAA’s GM Approval Database. Available online: http://www.isaaa.org/gmapprovaldatabase/ (accessed on 27 October 2021).
- English, J.J.; Mueller, E.; Baulcombe, D.C. Suppression of Virus Accumulation in Transgenic Plants Exhibiting Silencing of Nuclear Genes. Plant Cell 1996, 8, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, P.; Nelson, R.; De, B.; Hoffmann, N.; Rogers, S.; Fraley, R.; Beachy, R. Delay of Disease Development in Transgenic Plants That Express the Tobacco Mosaic Virus Coat Protein Gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef]
- Loesch-Fries, L.S.; Merlo, D.; Zinnen, T.; Burhop, L.; Hill, K.; Krahn, K.; Jarvis, N.; Nelson, S.; Halk, E. Expression of Alfalfa Mosaic Virus RNA 4 in Transgenic Plants Confers Virus Resistance. EMBO J. 1987, 6, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- van Dun, C.M.; Bol, J.F.; Van Vloten-Doting, L. Expression of Alfalfa Mosaic Virus and Tobacco Rattle Virus Coat Protein Genes in Transgenic Tobacco Plants. Virology 1987, 159, 299–305. [Google Scholar] [CrossRef]
- Tricoll, D.M.; Carney, K.J.; Russell, P.F.; McMaster, J.R.; Groff, D.W.; Hadden, K.C.; Himmel, P.T.; Hubbard, J.P.; Boeshore, M.L.; Quemada, H.D. Field Evaluation of Transgenic Squash Containing Single or Multiple Virus Coat Protein Gene Constructs for Resistance to Cucumber Mosaic Virus, Watermelon Mosaic Virus 2, and Zucchini Yellow Mosaic Virus. Nat. Biotechnol. 1995, 13, 1458–1465. [Google Scholar] [CrossRef]
- Fuchs, M.; Gonsalves, D. Resistance of Transgenic Hybrid Squash ZW-20 Expressing the Coat Protein Genes of Zucchini Yellow Mosaic Virus and Watermelon Mosaic Virus 2 to Mixed Infections by Both Potyviruses. Nat. Biotechnol. 1995, 13, 1466–1473. [Google Scholar] [CrossRef]
- Lius, S.; Manshardt, R.M.; Fitch, M.M.M.; Slightom, J.L.; Sanford, J.C.; Gonsalves, D. Pathogen-Derived Resistance Provides Papaya with Effective Protection against Papaya Ringspot Virus. Mol. Breed. 1997, 3, 161–168. [Google Scholar] [CrossRef]
- Collinge, D.B.; Sarrocco, S. Transgenic Approaches for Plant Disease Control: Status and Prospects 2021. Plant Pathol. 2021, ppa.13443. [Google Scholar] [CrossRef]
- Publication: USDA ARS. Available online: https://www.ars.usda.gov/research/publications/publication/?seqNo115=141655 (accessed on 2 March 2019).
- Ding, S.W.; Voinnet, O. Antiviral Immunity Directed by Small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Duan, C.-G.; Wang, C.-H.; Guo, H.-S. Application of RNA Silencing to Plant Disease Resistance. Silence 2012, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassengger, M.; Tepfer, M. Strategies for Antiviral Resistance in Transgenic Plants. Mol. Plant Pathol. 2008, 9, 73–83. [Google Scholar] [CrossRef]
- Reddy, D.; Sudarshana, M.; Fuchs, M.; Rao, N.; Thottappilly, G. Genetically Engineered Virus-Resistant Plants in Developing Countries: Current Status and Future Prospects. Adv. Virus Res. 2009, 75, 185–220. [Google Scholar] [CrossRef]
- Cisneros, A.E.; Carbonell, A. Artificial Small RNA-Based Silencing Tools for Antiviral Resistance in Plants. Plants 2020, 9, 669. [Google Scholar] [CrossRef]
- Fondong, V.N. The Search for Resistance to Cassava Mosaic Geminiviruses: How Much We Have Accomplished, and What Lies Ahead. Front. Plant Sci. 2017, 8, 408. [Google Scholar] [CrossRef] [Green Version]
- Fondong, V.N.; Nagalakshmi, U.; Dinesh-Kumar, S.P. Novel Functional Genomics Approaches: A Promising Future in the Combat Against Plant Viruses. Phytopathology 2016, 106, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-R.; Zhou, J.-J.; Hu, C.-G.; Wei, C.-L.; Zhang, J.-Z. MicroRNA-Mediated Gene Silencing in Plant Defense and Viral Counter-Defense. Front. Microbiol. 2017, 8, 1801. [Google Scholar] [CrossRef]
- Taliansky, M.; Samarskaya, V.; Zavriev, S.K.; Fesenko, I.; Kalinina, N.O.; Love, A.J. RNA-Based Technologies for Engineering Plant Virus Resistance. Plants 2021, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Maule, A.; Caranta, C.; Boulton, M. Sources of Natural Resistance to Plant Viruses: Status and Prospects. Mol. Plant Pathol. 2007, 8, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Ye, J.; Fang, R. Artificial MicroRNA-Mediated Virus Resistance in Plants. J. Virol. 2007, 81, 6690–6699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.-B.; Cai, W.-J.; Wang, J.-W.; Hong, G.-J.; Tao, X.-Y.; Wang, L.-J.; Huang, Y.-P.; Chen, X.-Y. Silencing a Cotton Bollworm P450 Monooxygenase Gene by Plant-Mediated RNAi Impairs Larval Tolerance of Gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Hada, A.; Patil, B.L.; Bajpai, A.; Kesiraju, K.; Dinesh-Kumar, S.; Paraselli, B.; Sreevathsa, R.; Rao, U. Micro RNA-Induced Gene Silencing Strategy for the Delivery of SiRNAs Targeting Meloidogyne Incognita in a Model Plant Nicotiana Benthamiana. Pest Manag. Sci. 2021, 77, 3396–3405. [Google Scholar] [CrossRef]
- Stella, S.; Montoya, G. The Genome Editing Revolution: A CRISPR-Cas TALE off-Target Story. Inside Cell 2016, 1, 7–16. [Google Scholar] [CrossRef]
- Piatek, A.; Mahfouz, M.M. Targeted Genome Regulation via Synthetic Programmable Transcriptional Regulators. Crit. Rev. Biotechnol. 2016, 37, 429–440. [Google Scholar] [CrossRef]
- Sera, T. Inhibition of Virus DNA Replication by Artificial Zinc Finger Proteins. J. Virol. 2005, 79, 2614–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; Takenaka, K.; Domoto, F.; Aoyama, Y.; Sera, T. Inhibition of Binding of Tomato Yellow Leaf Curl Virus Rep to Its Replication Origin by Artificial Zinc-Finger Protein. Mol. Biotechnol. 2013, 54, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, F.; Cai, J.; Chen, W.; Zhao, N.; Sun, Y.; Guo, Y.; Yang, X.; Wu, X. Artificial TALE as a Convenient Protein Platform for Engineering Broad-Spectrum Resistance to Begomoviruses. Viruses 2015, 7, 4772–4782. [Google Scholar] [CrossRef]
- Ceasar, S.A.; Rajan, V.; Prykhozhij, S.V.; Berman, J.N.; Ignacimuthu, S. Insert, Remove or Replace: A Highly Advanced Genome Editing System Using CRISPR/Cas9. Biochim. Biophys. Acta 2016, 1863, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.-A.; Tashkandi, M.; Mansoor, S.; Mahfouz, M.M. Engineering Plant Immunity: Using CRISPR/Cas9 to Generate Virus Resistance. Front. Plant Sci. 2016, 7, 1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating Highly Efficient Resistance against Wheat Dwarf Virus in Barley by Employing CRISPR/Cas9 System. Plant Biotechnol. J. 2019, 17, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR–Cas-like Immune System Conferring DNA Virus Resistance in Plants. Nat. Plants 2015, 1, 1–4. [Google Scholar] [CrossRef]
- Sanfaçon, H. Plant Translation Factors and Virus Resistance. Viruses 2015, 7, 3392–3419. [Google Scholar] [CrossRef] [Green Version]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-Mediated Editing of Cassava EIF4E Isoforms NCBP-1 and NCBP-2 Reduces Cassava Brown Streak Disease Symptom Severity and Incidence. Plant Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of Broad Virus Resistance in Non-Transgenic Cucumber Using CRISPR/Cas9 Technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [Green Version]
- Conner, A.J.; Jacobs, J.M. Genetic Engineering of Crops as Potential Source of Genetic Hazard in the Human Diet. Mutat. Res. 1999, 443, 223–234. [Google Scholar] [CrossRef]
- Thompson, J.R.; Tepfer, M. Assessment of the Benefits and Risks for Engineered Virus Resistance. Adv. Virus Res. 2010, 76, 33–56. [Google Scholar] [CrossRef]
- Masters, A.; Kang, M.; McCaw, M.; Zobrist, J.D.; Gordon-Kamm, W.; Jones, T.; Wang, K. Agrobacterium-Mediated Immature Embryo Transformation of Recalcitrant Maize Inbred Lines Using Morphogenic Genes. J. Vis. Exp. 2020, 156, e60782. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Liang, Y.; Lee, M.Y.; Pidatala, V.R.; Mortimer, J.C.; Scheller, H.V. Agrobacterium-Mediated Transient Transformation of Sorghum Leaves for Accelerating Functional Genomics and Genome Editing Studies. BMC Res. Notes 2020, 13, 116. [Google Scholar] [CrossRef] [Green Version]
- Elegba, W.; McCallum, E.; Gruissem, W.; Vanderschuren, H. Efficient Genetic Transformation and Regeneration of a Farmer-Preferred Cassava Cultivar From Ghana. Front. Plant Sci. 2021, 12, 668042. [Google Scholar] [CrossRef]
- Wagaba, H.; Beyene, G.; Aleu, J.; Odipio, J.; Okao-Okuja, G.; Chauhan, R.D.; Munga, T.; Obiero, H.; Halsey, M.E.; Ilyas, M.; et al. Field Level RNAi-Mediated Resistance to Cassava Brown Streak Disease across Multiple Cropping Cycles and Diverse East African Agro-Ecological Locations. Front. Plant Sci. 2017, 7, 2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, S.H.; Doudna, J.A. Expanding the Biologist’s Toolkit with CRISPR-Cas9. Mol. Cell 2015, 58, 568–574. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 Multiplex Editing Capability with the Endogenous TRNA-Processing System. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.; Kalam, A.; Amin, L.; Sidik, N.M. Gene Technology for Papaya Ringspot Virus Disease Management. Sci. World J. 2014, 2014, 768038. [Google Scholar] [CrossRef] [PubMed]
- Fei, Q.; Xia, R.; Meyers, B.C. Phased, Secondary, Small Interfering RNAs in Posttranscriptional Regulatory Networks. Plant Cell 2013, 25, 2400–2415. [Google Scholar] [CrossRef] [Green Version]
- Herben, T.; Suda, J.; Klimešová, J. Polyploid Species Rely on Vegetative Reproduction More than Diploids: A Re-Examination of the Old Hypothesis. Ann. Bot. 2017, 120, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadakuduti, S.S.; Buell, C.R.; Voytas, D.F.; Starker, C.G.; Douches, D.S. Genome Editing for Crop Improvement – Applications in Clonally Propagated Polyploids With a Focus on Potato (Solanum tuberosum L.). Front. Plant Sci. 2018, 9, 1607. [Google Scholar] [CrossRef] [Green Version]
- Sengeløv, G.; Kristensen, K.J.; Sørensen, A.H.; Kroer, N.; Sørensen, S.J. Effect of Genomic Location on Horizontal Transfer of a Recombinant Gene Cassette between Pseudomonas Strains in the Rhizosphere and Spermosphere of Barley Seedlings. Curr. Microbiol. 2001, 42, 160–167. [Google Scholar] [CrossRef]
- Panaud, O. Horizontal Transfers of Transposable Elements in Eukaryotes: The Flying Genes. Comptes Rendus Biol. 2016, 339, 296–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal Transfer of Transposable Elements and Why It Matters for Eukaryotic Evolution. Trends Ecol. Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Wallau, G.L.; Ortiz, M.F.; Loreto, E.L.S. Horizontal Transposon Transfer in Eukarya: Detection, Bias, and Perspectives. Genome Biol. Evol. 2012, 4, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.L.; Blackshaw, R.E.; Roy, J.; He, T. Investigation on Gene Transfer from Genetically Modified Corn (Zea mays L.) Plants to Soil Bacteria. J. Environ. Sci. Health B 2011, 46, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Valanzuolo, S.; Monti, M.M.; Colombo, M.; Cassani, G. Field release of transgenic virus tolerant tomatoes. In Fertilization in Higher Plants: Molecular and Cytological Aspects; Cresti, M., Cai, G., Moscatelli, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 405–412. ISBN 978-3-642-59969-9. [Google Scholar]
- Monti, M.; Valanzuolo, S.; Cassani, G.; Colombo, M. Transgenic tomatoes expressing a cucumber mosaic virus satellite RNA: Field testing and analysis of satellite RNA spread. J. Plant Pathol. 1999, 81, 113–122. [Google Scholar] [CrossRef]
- Ouedraogo, R.; Roossinck, M. CHAPTER 18: Molecular Evolution. In Cucumber Mosaic Virus; The American Phytopathological Society: St. Paul, MN, USA, 2019; pp. 207–215. ISBN 978-0-89054-610-9. [Google Scholar]
- Fondong, V.N.; Pita, J.S.; Rey, M.E.C.; De Kochko, A.; Beachy, R.N.; Fauquet, C.M. Evidence of Synergism between African Cassava Mosaic Virus and a New Double-Recombinant Geminivirus Infecting Cassava in Cameroon. J. Gen. Virol. 2000, 81, 287–297. [Google Scholar] [CrossRef]
- Pita, J.S.; Fondong, V.N.; Sangaré, A.; Otim-Nape, G.W.; Ogwal, S.; Fauquet, C.M. Recombination, Pseudorecombination and Synergism of Geminiviruses Are Determinant Keys to the Epidemic of Severe Cassava Mosaic Disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef]
- Patil, B.L.; Dangwal, M.; Mishra, R. Variability of Emaravirus Species Associated with Sterility Mosaic Disease of Pigeonpea in India Provides Evidence of Segment Reassortment. Viruses 2017, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Tentchev, D.; Verdin, E.; Marchal, C.; Jacquet, M.; Aguilar, J.M.; Moury, B. Evolution and Structure of Tomato Spotted Wilt Virus Populations: Evidence of Extensive Reassortment and Insights into Emergence Processes. J. Gen. Virol. 2011, 92, 961–973. [Google Scholar] [CrossRef]
- Catoni, M.; Noris, E.; Vaira, A.M.; Jonesman, T.; Matić, S.; Soleimani, R.; Behjatnia, S.A.A.; Vinals, N.; Paszkowski, J.; Accotto, G.P. Virus-Mediated Export of Chromosomal DNA in Plants. Nat. Commun. 2018, 9, 5308. [Google Scholar] [CrossRef]
- Chiba, S.; Kondo, H.; Tani, A.; Saisho, D.; Sakamoto, W.; Kanematsu, S.; Suzuki, N. Widespread Endogenization of Genome Sequences of Non-Retroviral RNA Viruses into Plant Genomes. PLoS Pathog. 2011, 7, e1002146. [Google Scholar] [CrossRef] [Green Version]
- Harper, G.; Hull, R.; Lockhart, B.; Olszewski, N. Viral Sequences Integrated into Plant Genomes. Annu. Rev. Phytopathol. 2002, 40, 119–136. [Google Scholar] [CrossRef]
- Liu, H.; Fu, Y.; Jiang, D.; Li, G.; Xie, J.; Cheng, J.; Peng, Y.; Ghabrial, S.A.; Yi, X. Widespread Horizontal Gene Transfer from Double-Stranded RNA Viruses to Eukaryotic Nuclear Genomes. J. Virol. 2010, 84, 11876–11887. [Google Scholar] [CrossRef] [Green Version]
- Belyi, V.A.; Levine, A.J.; Skalka, A.M. Unexpected Inheritance: Multiple Integrations of Ancient Bornavirus and Ebolavirus/Marburgvirus Sequences in Vertebrate Genomes. PLoS Pathog. 2010, 6, e1001030. [Google Scholar] [CrossRef] [Green Version]
- Crochu, S.; Cook, S.; Attoui, H.; Charrel, R.N.; De Chesse, R.; Belhouchet, M.; Lemasson, J.-J.; de Micco, P.; de Lamballerie, X. Sequences of Flavivirus-Related RNA Viruses Persist in DNA Form Integrated in the Genome of Aedes Spp. Mosquitoes. J. Gen. Virol. 2004, 85, 1971–1980. [Google Scholar] [CrossRef]
- Horie, M.; Honda, T.; Suzuki, Y.; Kobayashi, Y.; Daito, T.; Oshida, T.; Ikuta, K.; Jern, P.; Gojobori, T.; Coffin, J.M.; et al. Endogenous Non-Retroviral RNA Virus Elements in Mammalian Genomes. Nature 2010, 463, 84–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanne, E.; Sela, I. Occurrence of a DNA Sequence of a Non-Retro RNA Virus in a Host Plant Genome and Its Expression: Evidence for Recombination between Viral and Host RNAs. Virology 2005, 332, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.J.; Leach, R.W.; Bruenn, J. Filoviruses Are Ancient and Integrated into Mammalian Genomes. BMC Evol. Biol. 2010, 10, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.R.; Liu, H.; Shabman, R.S.; Ginell, G.M.; Luthra, P.; Ramanan, P.; Keefe, L.J.; Köllner, B.; Amarasinghe, G.K.; Taylor, D.J.; et al. Conservation of Structure and Immune Antagonist Functions of Filoviral VP35 Homologs Present in Microbat Genomes. Cell Rep. 2018, 24, 861–872.e6. [Google Scholar] [CrossRef] [Green Version]
- da Fonseca, G.C.; de Oliveira, L.F.V.; de Morais, G.L.; Abdelnor, R.V.; Nepomuceno, A.L.; Waterhouse, P.M.; Farinelli, L.; Margis, R. Unusual RNA Plant Virus Integration in the Soybean Genome Leads to the Production of Small RNAs. Plant Sci. 2016, 246, 62–69. [Google Scholar] [CrossRef]
- Stein, T.; Ran, G.; Bohmer, M.; Sharbati, S.; Einspanier, R. Expression Profiling of Key Pathways in Rat Liver after a One-Year Feeding Trial with Transgenic Maize MON810. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vendômois, J.S.; Roullier, F.; Cellier, D.; Séralini, G.-E. A Comparison of the Effects of Three GM Corn Varieties on Mammalian Health. Int. J. Biol. Sci. 2009, 5, 706–726. [Google Scholar] [CrossRef] [Green Version]
- Martelli, G.P. Transgenic resistance to plant pathogens: Benefits and risks. J. Plant. Pathol. 2001, 83, 37–46. [Google Scholar]
- Nielsen, K.M.; Bones, A.M.; Smalla, K.; van Elsas, J.D. Horizontal Gene Transfer from Transgenic Plants to Terrestrial Bacteria--a Rare Event? FEMS Microbiol. Rev. 1998, 22, 79–103. [Google Scholar] [CrossRef]
- Patterson, L.A.; Josling, T. Regulating biotechnology: Comparing EU and US approaches. In Environmental Policy in the EU; Routledge: London, UK, 2005. [Google Scholar]
- Jaffe, G. Regulatory Slowdown on GM Crop Decisions. Nat. Biotechnol. 2006, 24, 748–749. [Google Scholar] [CrossRef]
- Smyth, S.J. Canadian Regulatory Perspectives on Genome Engineered Crops. GM Crop. Food 2016, 8, 35–43. [Google Scholar] [CrossRef]
- Papadopoulou, N.; Devos, Y.; Álvarez-Alfageme, F.; Lanzoni, A.; Waigmann, E. Risk Assessment Considerations for Genetically Modified RNAi Plants: EFSA’s Activities and Perspective. Front. Plant Sci. 2020, 11, 445. [Google Scholar] [CrossRef] [Green Version]
- Okeno, J.A.; Wolt, J.D.; Misra, M.K.; Rodriguez, L. Africa’s Inevitable Walk to Genetically Modified (GM) Crops: Opportunities and Challenges for Commercialization. New Biotechnol. 2013, 30, 124–130. [Google Scholar] [CrossRef]
- Kreuze, J.F.; Valkonen, J.P. Utilization of Engineered Resistance to Viruses in Crops of the Developing World, with Emphasis on Sub-Saharan Africa. Curr. Opin. Virol. 2017, 26, 90. [Google Scholar] [CrossRef]
- Gonsalves, D. Control of Papaya Ringspot Virus in Papaya: A Case Study. Annu. Rev. Phytopathol. 1998, 36, 415–437. [Google Scholar] [CrossRef] [Green Version]
- Gonsalves, D. Transgenic Papaya: Development, Release, Impact and Challenges. Adv. Virus Res. 2006, 67, 317–354. [Google Scholar] [CrossRef]
- Thomas, P.E.; Kaniewski, W.K.; Lawson, E.C. Reduced Field Spread of Potato Leafroll Virus in Potatoes Transformed with the Potato Leafroll Virus Coat Protein Gene. Plant Dis. 1997, 81, 1447–1453. [Google Scholar] [CrossRef]
- Kaniewski, W.; Thomas, P. The Potato Story. AgBioForum 2004, 7, 41–46. [Google Scholar]
- Newell, C.A.; Rozman, R.; Hinchee, M.A.; Lawson, E.C.; Haley, L.; Sanders, P.; Kaniewski, W.; Tumer, N.E.; Horsch, R.B.; Fraley, R.T. Agrobacterium-Mediated Transformation of Solanum Tuberosum L. Cv. ‘Russet Burbank’. Plant Cell Rep. 1991, 10, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Scorza, R.; Ravelonandro, M.; Callahan, A.M.; Cordts, J.M.; Fuchs, M.; Dunez, J.; Gonsalves, D. Transgenic Plums (Prunus Domestica L.) Express the Plum Pox Virus Coat Protein Gene. Plant Cell Rep. 1994, 14, 18–22. [Google Scholar] [CrossRef]
- Ilardi, V.; Tavazza, M. Biotechnological Strategies and Tools for Plum Pox Virus Resistance: Trans-, Intra-, Cis-Genesis, and Beyond. Front. Plant Sci. 2015, 6, 379. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.J.; Ying, Z. Development of Papaya Breeding Lines with Transgenic Resistance to Papaya Ringspot Virus. Plant Dis. 2004, 88, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.-X.; Ou-Yang, W.-J.; Zhang, Y.-F.; Chen, Z.-L. Transgenic Sweet Pepper Plants from Agrobacterium Mediated Transformation. Plant Cell Rep. 1996, 16, 71–75. [Google Scholar] [CrossRef]
- Yang, R.; Xu, H.; Long, M.; Yu, W.; Lu, C.; Pan, N.; Chen, Z. Transgenic Tomato Plants Expressing Cucumber Mosaic Virus Coat Protein and Their Resistance to CMV. Jiangsu J. Agric. Sci. 1995, 11, 40–44. [Google Scholar]
- Bonfim, K.; Faria, J.C.; Nogueira, E.O.P.L.; Mendes, É.A.; Aragão, F.J.L. RNAi-Mediated Resistance to Bean Golden Mosaic Virus in Genetically Engineered Common Bean (Phaseolus Vulgaris). Mol. Plant Microbe Interact. 2007, 20, 717–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollefson, J. Brazil Cooks up Transgenic Bean. Nature 2011, 478, 168. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.A.; Pitz, K.Y.; Manshardt, R.; Zee, F.; Fitch, M.; Gonsalves, D. Virus Coat Protein Transgenic Papaya Provides Practical Control of Papaya Ringspot Virus in Hawaii. Plant Dis. 2002, 86, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, S.; Suzuki, J.; Gonsalves, D. Development of Genetically Engineered Resistant Papaya for Papaya Ringspot Virus in a Timely Manner: A Comprehensive and Successful Approach. Methods Mol. Biol. 2007, 354, 197–240. [Google Scholar] [CrossRef]
- Guo, J.; Yang, L.; Liu, X.; Guan, X.; Jiang, L.; Zhang, D. Characterization of the Exogenous Insert and Development of Event-Specific PCR Detection Methods for Genetically Modified Huanong No. 1 Papaya. J. Agric. Food Chem. 2009, 57, 7205–7212. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.; Dale, M.F.B. Resistance to viruses in potato. In Natural Resistance Mechanisms of Plants to Viruses; Loebenstein, G., Carr, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 341–366. [Google Scholar]
- Palukaitis, P. Resistance to Viruses of Potato and Their Vectors. Plant Pathol. J. 2012, 28, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering Resistance to Mixed Virus Infection in a Commercial Potato Cultivar: Resistance to Potato Virus X and Potato Virus Y in Transgenic Russet Burbank. Nat. Biotechnol. 1990, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kaniewski, W.; Lawson, C.; Sammons, B.; Haley, L.; Hart, J.; Delannay, X.; Tumer, N.E. Field Resistance of Transgenic Russeet Burbank Potato to Effects of Infection by Potato Virus X and Potato Virus Y. Nat. Biotechnol. 1990, 8, 750–754. [Google Scholar] [CrossRef]
- Tomar, V.; Saini, S.S.; Juneja, K.; Agrawal, P.K.; Sircar, D. Transgenic technologies and their potential applications in horticultural crop improvement. In Advances in Plant Transgenics: Methods and Applications; Sathishkumar, R., Kumar, S.R., Hema, J., Baskar, V., Eds.; Springer: Singapore, 2019; pp. 189–212. ISBN 9789811396243. [Google Scholar]
- Thomas, P.E.; Lawson, E.C.; Zalewski, J.C.; Reed, G.L.; Kaniewski, W.K. Extreme Resistance to Potato Leafroll Virus in Potato Cv. Russet Burbank Mediated by the Viral Replicase Gene. Virus Res. 2000, 71, 49–62. [Google Scholar] [CrossRef]
- Lawson, E.C.; Weiss, J.D.; Thomas, P.E.; Kaniewski, W.K. NewLeaf Plus® Russet Burbank Potatoes: Replicase-Mediated Resistance to Potato Leafroll Virus. Mol. Breed. 2001, 7, 1–12. [Google Scholar] [CrossRef]
- Kenyon, L.; Tsai, W.S.; Shih, S.L.; Lee, L.M. Emergence and Diversity of Begomoviruses Infecting Solanaceous Crops in East and Southeast Asia. Virus Res. 2014, 186, 104–113. [Google Scholar] [CrossRef]
- Leke, W.N.; Mignouna, D.B.; Brown, J.K.; Kvarnheden, A. Begomovirus Disease Complex: Emerging Threat to Vegetable Production Systems of West and Central Africa. Agric. Food Secur. 2015, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Morales, F.J.; Jones, P.G. The Ecology and Epidemiology of Whitefly-Transmitted Viruses in Latin America. Virus Res. 2004, 100, 57–65. [Google Scholar] [CrossRef]
- Morales, F.J. History and Current Distribution of Begomoviruses in Latin America. Adv. Virus Res. 2006, 67, 127–162. [Google Scholar] [CrossRef]
- Faria, J.; Aragão, F.J.L.; Souza, T.L.P.O.; Quintela, E.; Kitajima, E.W.; Ribeiro, S. Golden Mosaic of Common Beans in Brazil: Management with a Transgenic Approach. APS Features 2016, 159, 727–737. [Google Scholar]
- Pinheiro-Lima, B.; Pereira-Carvalho, R.C.; Alves-Freitas, D.M.T.; Kitajima, E.W.; Vidal, A.H.; Lacorte, C.; Godinho, M.T.; Fontenele, R.S.; Faria, J.C.; Abreu, E.F.M.; et al. Transmission of the Bean-Associated Cytorhabdovirus by the Whitefly Bemisia Tabaci MEAM1. Viruses 2020, 12, 1028. [Google Scholar] [CrossRef]
- Kyle, M. Resistance to Viral Diseases of Vegetables: Genetic and Breeding; Universidad Autónoma Chapingo: Portland, OR, USA, 1993; p. 278. [Google Scholar]
- Johnson, S.; Strome, S. Quantification of the Impacts on US Agriculture of Biotechnology-Derived Crops Planted in 2006; National Center for Food and Agricultural Policy: Washington, DC, USA, 2006. [Google Scholar]
- Mbanzibwa, D.R.; Tian, Y.P.; Tugume, A.K.; Patil, B.L.; Yadav, J.S.; Bagewadi, B.; Abarshi, M.M.; Alicai, T.; Changadeya, W.; Mkumbira, J.; et al. Evolution of Cassava Brown Streak Disease-Associated Viruses. J. Gen. Virol. 2011, 92, 974–987. [Google Scholar] [CrossRef]
- Patil, B.L.; Legg, J.P.; Kanju, E.; Fauquet, C.M. Cassava Brown Streak Disease: A Threat to Food Security in Africa. J. Gen. Virol. 2015, 96, 956–968. [Google Scholar] [CrossRef] [Green Version]
- Patil, B.L.; Fauquet, C.M. Cassava Mosaic Geminiviruses: Actual Knowledge and Perspectives. Mol. Plant Pathol. 2009, 10, 685–701. [Google Scholar] [CrossRef]
- National Biosafety Authority. NBA Board Approves Environmental Release Application for GM Cassava. Available online: https://www.biosafetykenya.go.ke/index.php?option=com_content&view=article&id=258:nba-board-approves-environmental-release-application-for-gm-cassava&catid=16&Itemid=156 (accessed on 24 September 2021).
- Odipio, J.; Ogwok, E.; Taylor, N.J.; Halsey, M.; Bua, A.; Fauquet, C.M.; Alicai, T. RNAi-Derived Field Resistance to Cassava Brown Streak Disease Persists across the Vegetative Cropping Cycle. GM Crop. Food 2014, 5, 16–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogwok, E.; Odipio, J.; Halsey, M.; Gaitán-Solís, E.; Bua, A.; Taylor, N.J.; Fauquet, C.M.; Alicai, T. Transgenic RNA Interference (RNAi)-Derived Field Resistance to Cassava Brown Streak Disease. Mol. Plant Pathol. 2012, 13, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- WHO. 2014 Frequently Asked Questions on Genetically Modified Food. Available online: https://www.who.int/news-room/q-a-detail/food-genetically-modified (accessed on 27 October 2021).
- NASEM. Genetically Engineered Crops: Experiences and Prospects; The National Academies Press: Washington, DC, USA, 2016; p. 388. [Google Scholar]
- Lee, R. Review of The Regulation of Genetically Modified Organisms: Comparative Approaches. J. Environ. Law 2010, 22, 511–514. [Google Scholar] [CrossRef]
- Ichim, M.C. The More Favorable Attitude of the Citizens toward GMOs Supports a New Regulatory Framework in the European Union. GM Crops Food 2021, 12, 18–24. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Country | Year of Approval | Crop /Cultivated Area | Technology Company/Organization | Trade Name | Target Viruses | Gene Targeted | References |
|---|---|---|---|---|---|---|---|
| USA | 1996 | Squash * | Seminis & Monsanto | CZW3, ZW20 | Cucumber mosaic virus (CMV), Watermelon mosaic virus (WMV), Zucchini yellow mosaic virus (ZYMV) | Coat protein | [16] |
| 1997 | Papaya * | Cornell University and the University of Hawaii | 55-1 (Rainbow & SunUp), 63-1 | Papaya ringspot virus (PRSV) | Coat protein | [91,92] | |
| 1998 | Potato * | Monsanto | NewLeaf™Plus Russet Burbank | Potato leaf roll virus (PLRV) | Replicase and helicase | [93,94] | |
| Shepody NewLeaf™Y potato | Potato virus Y (PVY) | Coat protein | [94,95] | ||||
| 2007 | Plum | USDA-ARS | C-5 | Plum pox virus (PPV) | Coat protein | [96,97] | |
| 2009 | Papaya * | University of Florida | X17-2 | Papaya ringspot virus (PRSV) | Coat protein | [98] | |
| China | 1998 | Sweet pepper | Beijing University | PK-SP01 | Cucumber mosaic virus (CMV) | Coat protein | [99] |
| 1999 | Tomato * | Beijing University | PK-TM8805R(8805R) | Cucumber mosaic virus (CMV) | Coat protein | [100] | |
| 2006 | Papaya * | Cornell University and the University of Hawaii | 55-1(Rainbow & SunUp), 63-1 | Papaya ringspot virus (PRSV) | Coat protein | [91,92] | |
| Canada | 1999 | Potato * | Monsanto | Shepody NewLeaf™Y potato | Potato virus Y (PVY) | Coat protein | [94,95] |
| Brazil | 2011 | Bean * | EMBRAPA | BRSFc401 RMD | Bean golden mosaic virus (BGMV) | RNA of viral replication protein | [101,102] |
| Japan | 2011 | Papaya * | Cornell University and the University of Hawaii | 55-1 (Rainbow & SunUp), 63-1 | Papaya ringspot virus (PRSV) | Coat protein | [91,92] |
| Kenya | 2021 | Cassava | Danforth Plant Science Center, St. Louis, USA | Cassava Event-4046 | Cassava brown streak virus (CBSV) | RNAi against CP | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niraula, P.M.; Fondong, V.N. Development and Adoption of Genetically Engineered Plants for Virus Resistance: Advances, Opportunities and Challenges. Plants 2021, 10, 2339. https://doi.org/10.3390/plants10112339
Niraula PM, Fondong VN. Development and Adoption of Genetically Engineered Plants for Virus Resistance: Advances, Opportunities and Challenges. Plants. 2021; 10(11):2339. https://doi.org/10.3390/plants10112339
Chicago/Turabian StyleNiraula, Prakash M., and Vincent N. Fondong. 2021. "Development and Adoption of Genetically Engineered Plants for Virus Resistance: Advances, Opportunities and Challenges" Plants 10, no. 11: 2339. https://doi.org/10.3390/plants10112339