
Alternative Pathway Is Involved in Hydrogen Peroxide-Enhanced Cadmium Tolerance in Hulless Barley Roots


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. H2O2 Staining
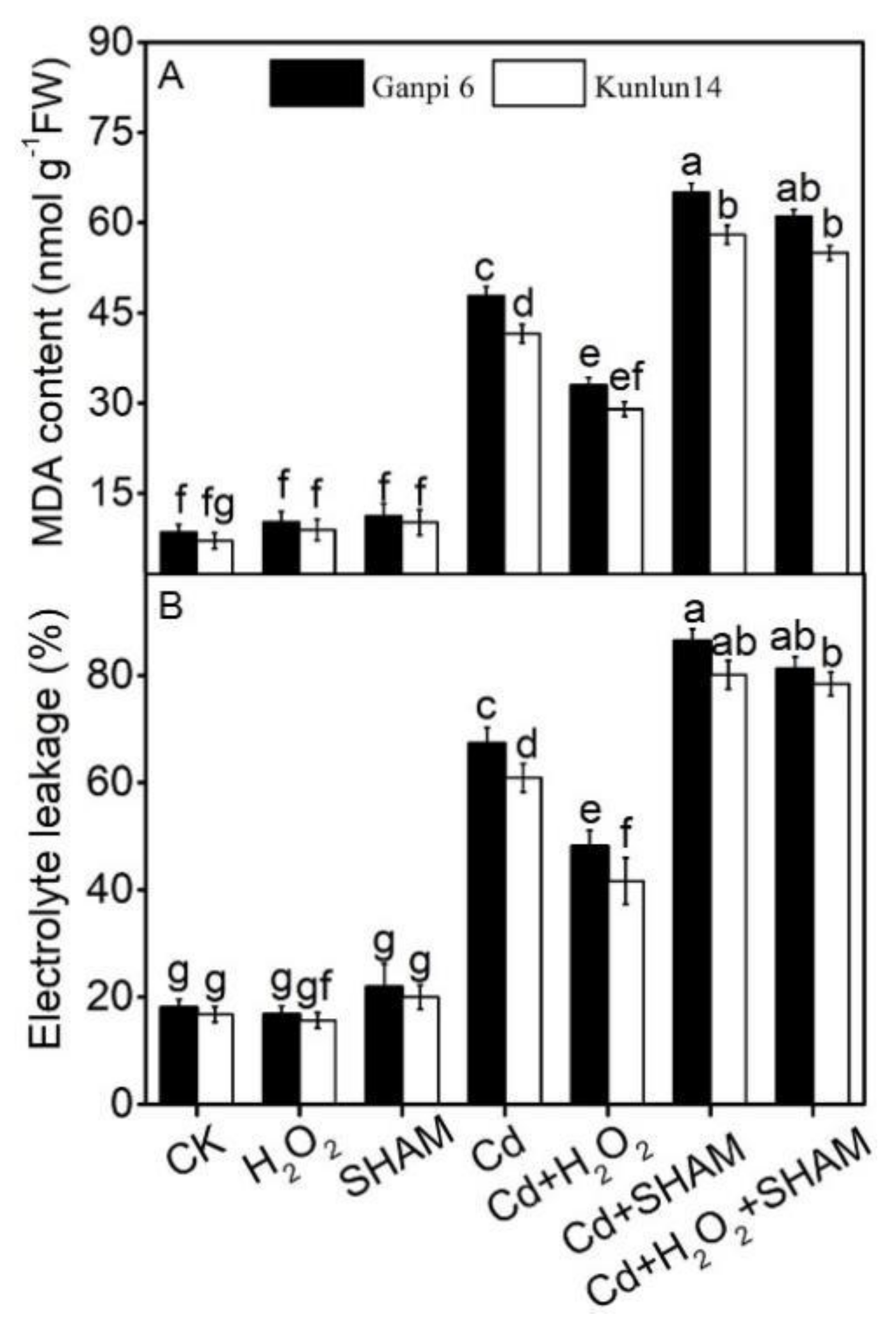
2.3. Determination of Electrolyte Leakage and Malondialdehyde Content
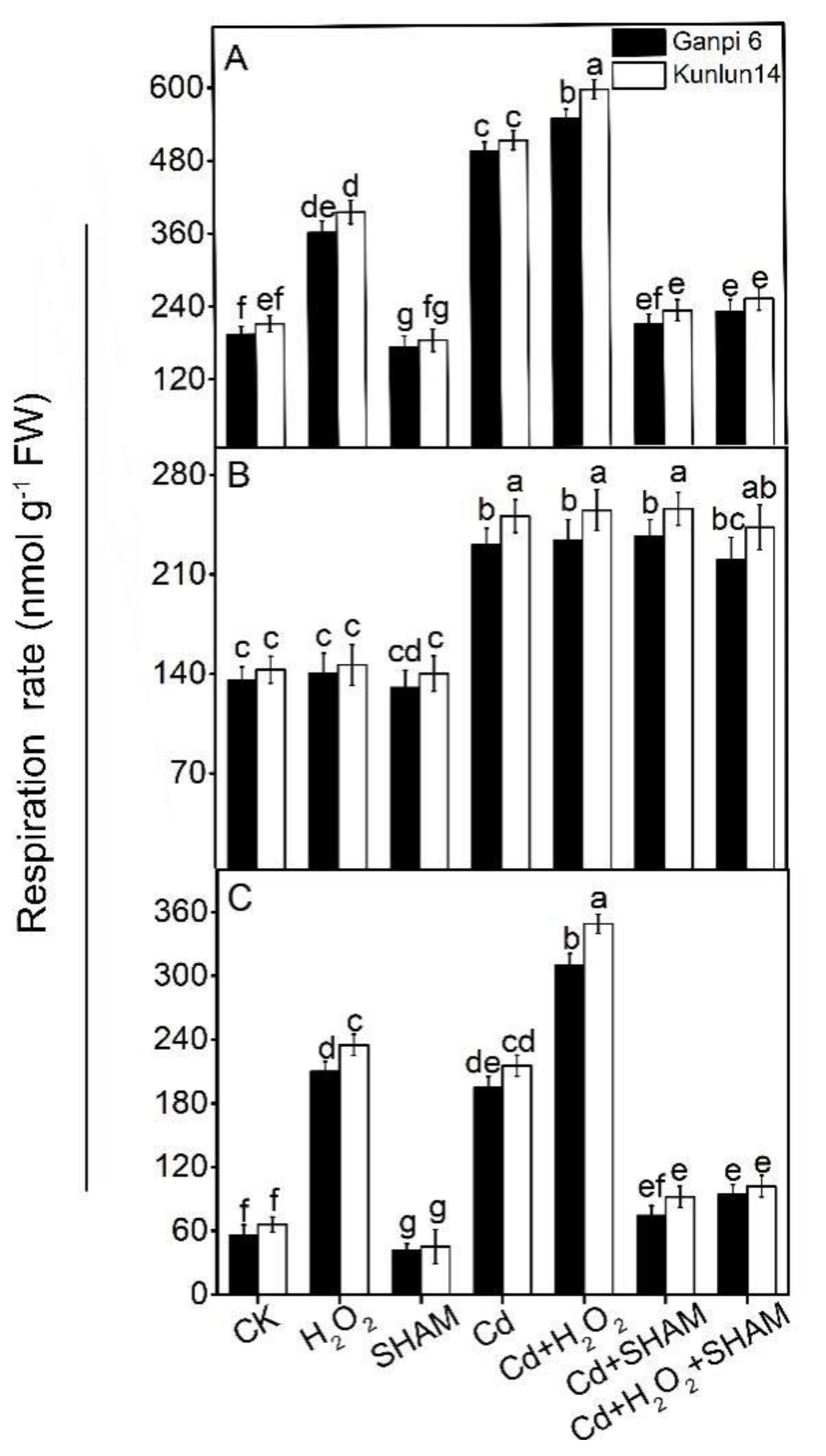
2.4. Measurements of Respiratory Rates
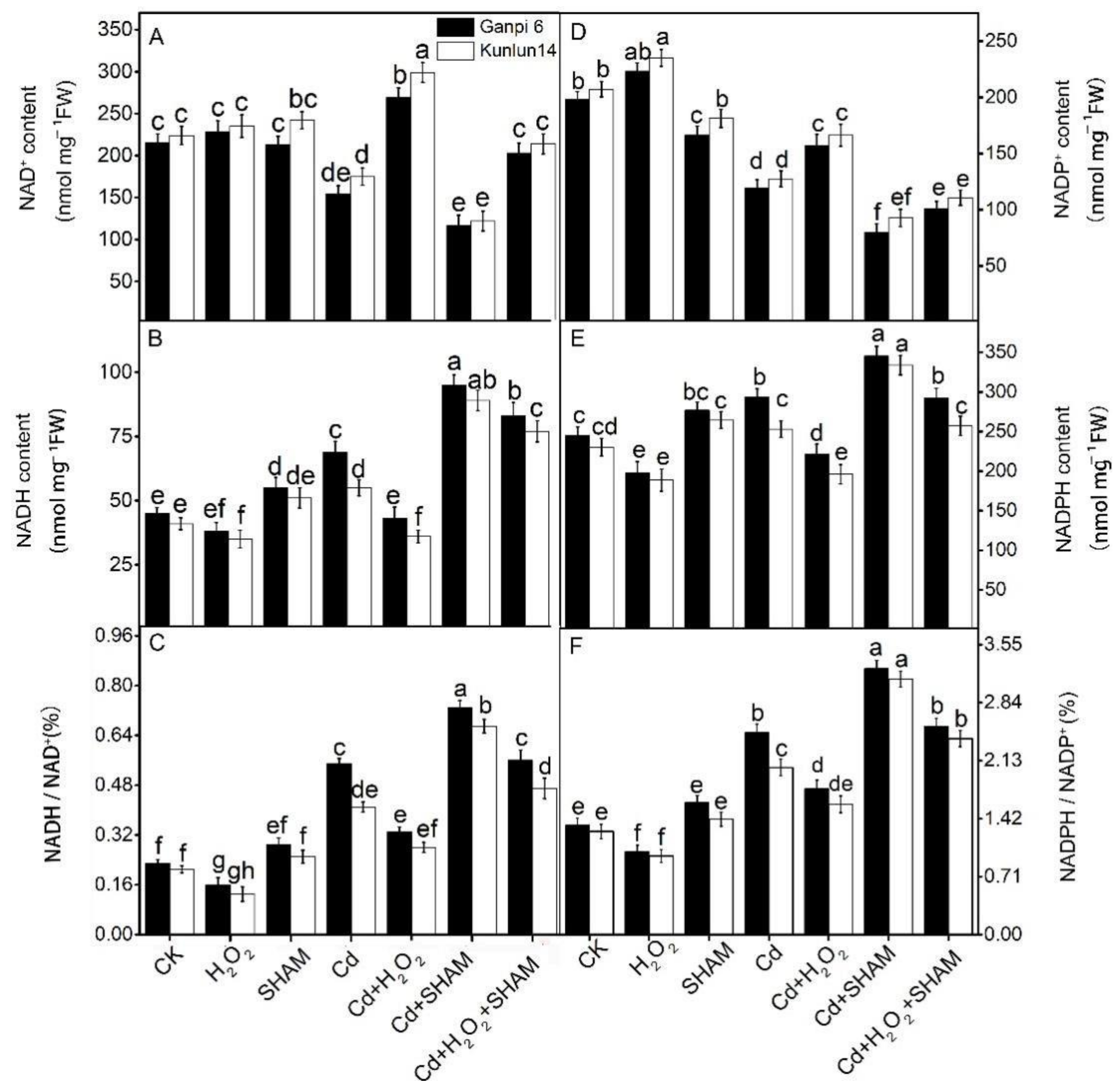
2.5. Determination of Antioxidant Contents
2.6. Antioxidant Enzyme Activity Assay
2.7. RNA Isolation and qRT-PCR
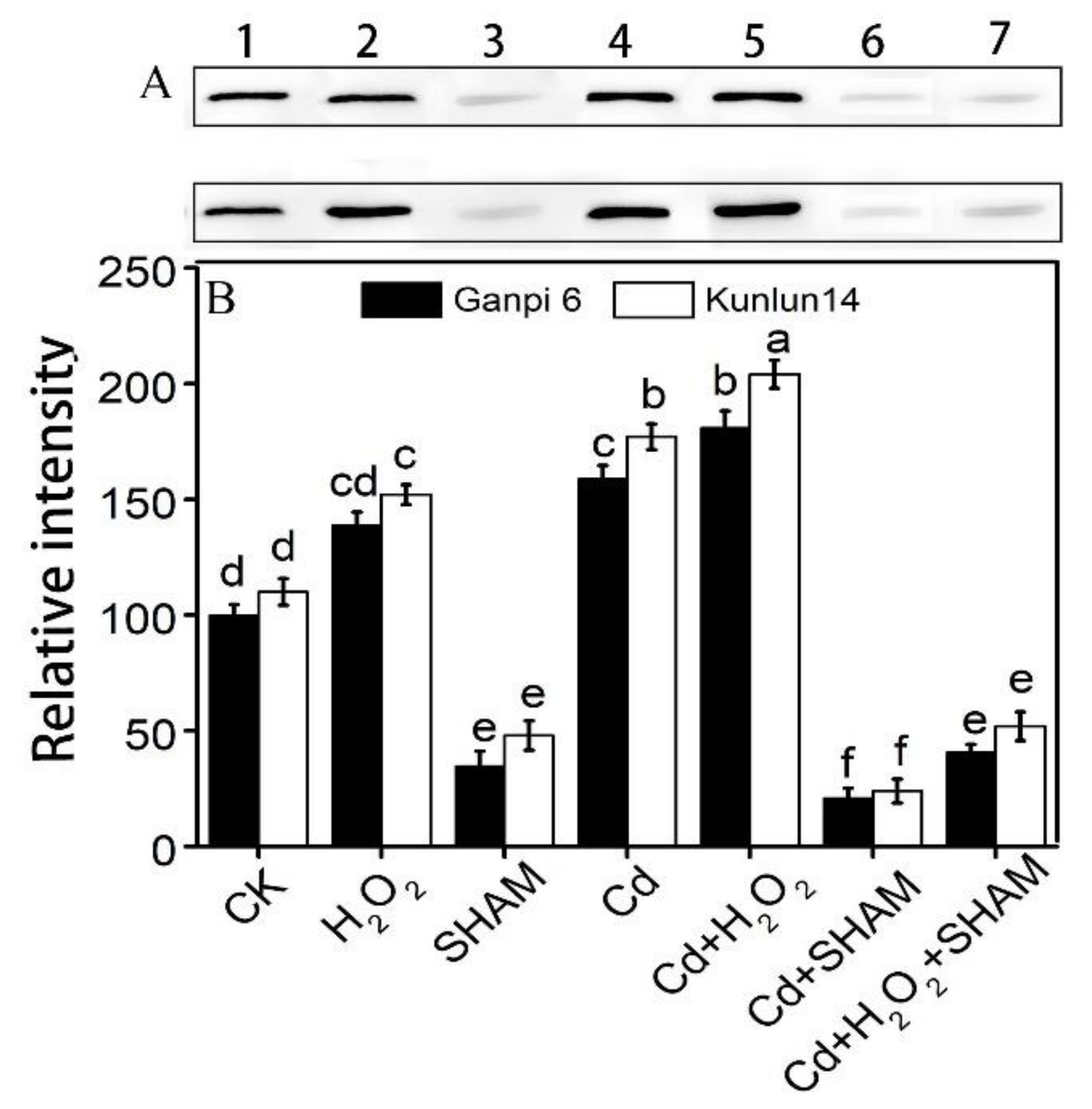
2.8. Western-Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of Cd Stress on H2O2 Content
3.2. Exogenous H2O2 Enhances HvAOXs Expression in Ganpi6 and Kunlun14 Roots under Cd Stress
3.3. Exogenous H2O2 Enhances AP Capacity (Valt) under Cd Stress
3.4. Exogenous H2O2 Enhances AOX Protein Accumulation under Cd Stress
3.5. AP Is Involved in the Regulation of H2O2 in Barley Tolerance to Cd Stress
3.6. Effects of H2O2 and SHAM on NADH/NAD+ and NADPH/NADP+ under Cd Stress
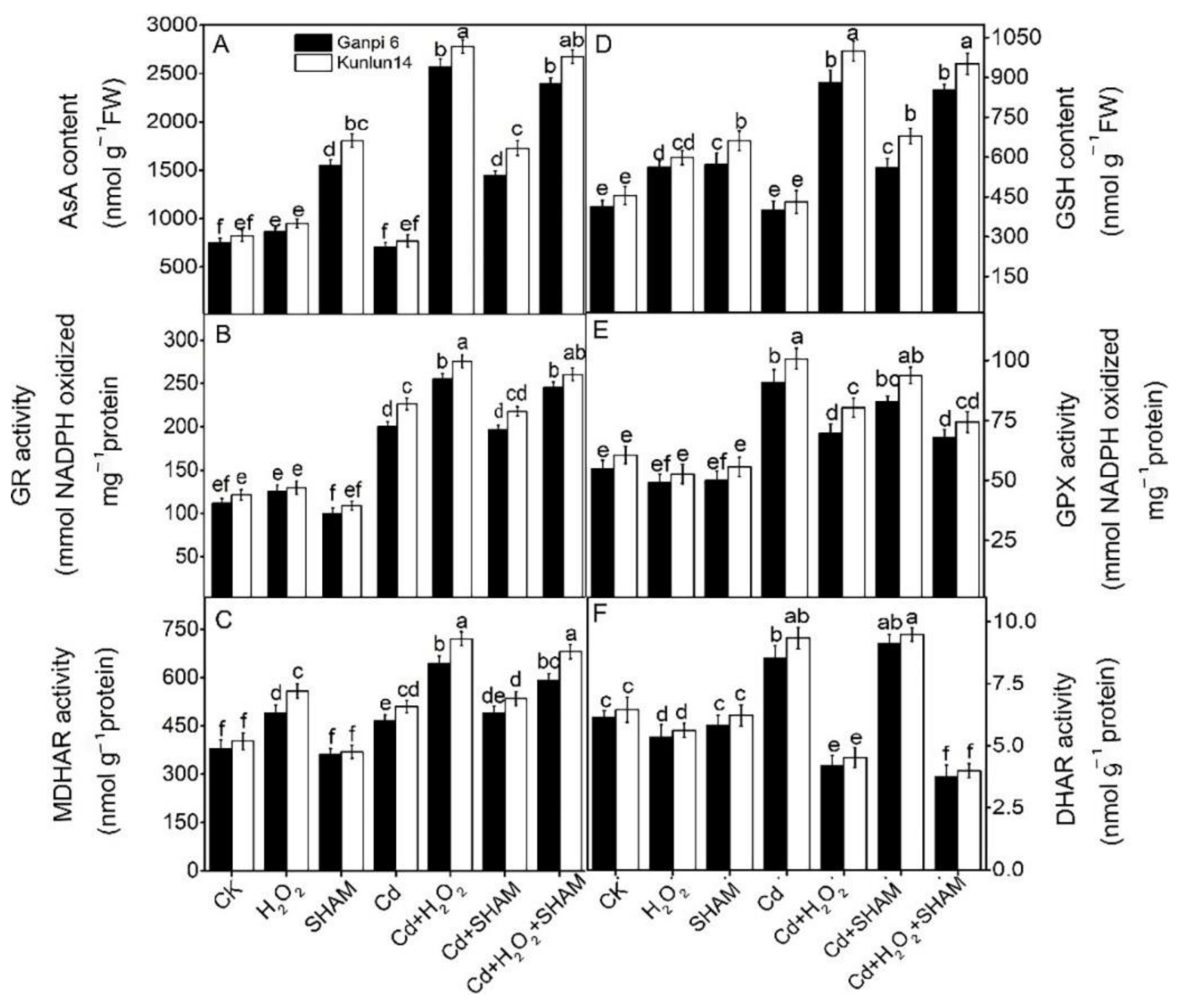
3.7. Effects of Exogenous H2O2 and SHAM on the AsA-GSH Cycle under Cd Stress
3.8. Effects of Exogenous H2O2 and SHAM on Antioxidant Enzyme Activities under Cd Stress
3.9. Effects of Exogenous H2O2 and SHAM on the Expression of Antioxidant Enzyme Genes under Cd Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizerss. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Fasani, E.; Manara, A.; Martini, F.; Furini, A.; Dalcorso, G. The potential of genetic engineering of plants for the remediation of soils contaminated with heavy metals. Plant Cell Environ. 2018, 41, 1201–1232. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chen, L.; Li, X. Arabidopsis and rice showed a distinct pattern in ZIPs genes expression profile in response to Cd stress. Bot. Stud. 2018, 59, 22. [Google Scholar] [CrossRef]
- Zhang, W.; Yun, L.; Zhao, Y.L.; Xu, Z.G.; Huang, H.M.; Zhou, J.K.; Yang, G.Y. Morphological and physiological changes of Broussonetia papyrifera seedlings in cadmium contaminated soil. Plants 2020, 9, 1698. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.N.; Wang, H.T.; Song, J.; Cao, X.; Rogers, H.J.; Francis, D.; Jia, C.Y.; Sun, L.Z.; Hou, M.F.; Tai, P.D.; et al. Cell cycle arrest mediated by Cd-induced DNA damage in Arabidopsis root tips. Ecotoxicol. Environ. Saf. 2017, 145, 569–574. [Google Scholar] [CrossRef]
- Na, Q.C.; Lu, X.Y.; Guo, X.R.; Pan, Y.J.; Yu, B.F.; Tang, Z.H.; Guo, Q.X. Differential responses to Cd stress induced by exogenous application of Cu, Zn or Ca in the medicinal plant Catharanthus roseus. Ecotoxicol. Environ. Saf. 2018, 157, 266–275. [Google Scholar]
- Sharma, S.S.; Dietz, K.J.; Mimura, T. Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ. 2016, 39, 1112–1126. [Google Scholar] [CrossRef]
- Xie, X.; Weiss, D.J.; Weng, B.; Liu, J.; Lu, H.; Yan, C. The short-term effect of cadmium on low molecular weight organic acid and amino acid exudation from mangrove (Kandelia obovata (S., L.) Yong) roots. Environ. Sci. Pollut. Res. 2013, 20, 997–1008. [Google Scholar] [CrossRef]
- Cuypers, A.; Smeets, K.; Ruytinx, J.; Opdenakker, K.; Keunen, E.; Remans, T.; Horemans, N.; Vanhoudt, N.; Van, S.S.; Van, B.F.; et al. The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings. J. Plant Physiol. 2011, 168, 309–316. [Google Scholar] [CrossRef]
- Huang, W.J.; Yang, X.D.; Yao, S.C.; Thet, L.; He, H.Y.; Wang, A.Q.; Li, C.Z.; He, L.F. Reactive oxygen species burst induced by aluminum stress triggers mitochondria-dependent programmed cell death in peanut root tip cells. Plant Physiol. Biochem. 2014, 82, 76–84. [Google Scholar] [CrossRef]
- Malá, J.; Cvrčková, H.; Máchová, P.; Dostál, J.; Šíma, P. Heavy metal accumulation by willow clones in short-time hydroponics. J. For. Sci. 2010, 56, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Helmut, S. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox. Biol. 2017, 11, 613–619. [Google Scholar]
- Zhou, J.; Xia, X.J.; Zhou, Y.H.; Shi, K.; Chen, Z.; Yu, J.Q. OH1- dependent H2O2 production and subsequent activation of MPK1/2 play an important role in acclimation-induced cross-tolerance in tomato. J. Exp. Bot. 2014, 65, 595–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildiz, M.; Terzi, H.; Bingül, N. Protective role of hydrogen peroxide pretreatmenton defense systems and BnMP1 gene expression in Cr (VI)- stressed canola seedlings. Ecotoxicology 2013, 22, 1303–1312. [Google Scholar] [CrossRef]
- Pavel, V.; Fedurayev, K.S.; Mironov, D.A.; Gabrielyan, V.S.; Bedbenov, A.A.; Zorina, M.S.; Dmitry, A.L. Hydrogen peroxide participates in perception and transduction of cold stress signal in synechocystis. Plant Cell Physiol. 2018, 59, 1255–1264. [Google Scholar]
- Helmut, S. Role of metabolic H2O2 generation: Redox signaling and oxidative stress. J. Biol. Chem. 2014, 289, 8735–8741. [Google Scholar]
- Yang, C.Y. Hydrogen peroxide controls transcriptional responses of ERF73/HRE1 and ADH1 via modulation of ethylene signaling during hypoxic stress. Planta 2014, 39, 877–885. [Google Scholar] [CrossRef]
- Vanlerberghe, G.C.; Day, D.A.; Wiskich, J.T.; Vanlerberghe, A.E.; McIntosh, L. AOX activity in tobacco leaf mitochondria: Dependence on tricarboxylic acid cycle-mediated redox regulation and pyruvate activation. Plant Physiol. 1995, 109, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daurelio, L.D.; Checa, S.K.; Barrio, J.M.; Ottado, J.; Orellano, E.G. Characterization of Citrus sinensis type 1 mitochondrial alternative oxidase and expression analysis in biotic stress. Biosci. Rep. 2010, 30, 59–71. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.M.; Zhao, C.Z.; Wang, J.F.; Li, P.; Dou, Y.Q.; Bi, Y.R. Alternative pathway is involved in the tolerance of highland barley to the low-nitrogen stress by maintaining the cellular redox homeostasis. Plant Cell Rep. 2016, 35, 317–328. [Google Scholar] [CrossRef]
- Zhao, C.Z.; Wang, X.M.; Wu, K.; Li, P.; Chang, N.; Wang, J.F.; Wang, F.; Li, J.; Bi, Y.R. Glucose-6-phosphate dehydrogenase and alternative oxidase are involved in the cross tolerance of highland barley to salt stress and UV-B radiation. J. Plant Physiol. 2015, 181, 83–95. [Google Scholar] [CrossRef]
- Castro-Guerrero, N.A.; Rodríguez-Zavala, J.S.; Marín-Hernández, A.; Rodríguez-Enríquez, S.; Moreno-Sánchez, R. Enhanced alternative oxidase and antioxidant enzymes under Cd2+ stress in Euglena. J. Bioenerg. Biomembr. 2008, 40, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, X.; Dou, Y.; Liu, D.; Si, W.; Fang, H.; Zhao, C.; Chen, S.; Xi, J.; Li, J. Hydrogen sulfide-cysteine cycle system enhances cadmium tolerance through alleviating cadmium-induced oxidative stress and ion toxicity in Arabidopsis roots. Sci. Rep. 2016, 6, 39702. [Google Scholar] [CrossRef] [PubMed]
- Yukioka, H.; Inagaki, S.; Tanaka, R.; Katoh, K.; Miki, N.; Mizutani, A.; Masuko, M. Transcriptional activation of the alternative oxidase gene of the fungus Magnaporthe grisea by a respiratory-inhibiting fungicide and hydrogen peroxide. Biochim. Biophys. Acta 1998, 1442, 161–169. [Google Scholar] [CrossRef]
- Wang, H.H.; Huang, J.J.; Bi, Y.R. Induction of alternative respiratory pathway involves nitric oxide, hydrogen peroxide and ethylene under salt stress. Plant Signal. Behavior. 2010, 5, 1636–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Huang, J.J.; Liang, X.L.; Bi, Y.R. Involvement of hydrogen peroxide, calcium, and ethylene in the induction of the alternative pathway in chilling-stressed Arabidopsis callus. Planta 2012, 235, 53–67. [Google Scholar] [CrossRef]
- Paradiso, A.; Berardino, R.; Pinto, M.C.; Sanità, T.L.; Storelli, M.M.; Tommasi, F.; De, G.L. Increase in ascorbate-glutathione metabolism as local and precocious systemic responses induced by cadmium in durum wheat plants. Plant Cell Physiol. 2008, 49, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Skórzyńska, P.E.; Dra, M.; Krupa, Z. The activity of the antioxidative system in cadmium-treated Arabidopsis thaliana. Biol. Plant. 2003, 47, 71–78. [Google Scholar] [CrossRef]
- Jian, W.; Zhang, D.W.; Zhu, F.; Wang, S.X.; Pu, X.J.; Deng, X.G.; Luo, S.S.; Lin, H.H. Alternative oxidase pathway is involved in the exogenous SNP-elevated tolerance of Medicago truncatula to salt stress. J. Plant Physiol. 2016, 193, 79–87. [Google Scholar] [CrossRef]
- He, L.; Wang, X.M.; Feng, R.J.; He, Q.; Wang, S.W.; Liang, C.F.; Yan, L.L.; Bi, Y.R. Alternative pathway is involved in nitric oxide enhanced tolerance to cadmium stress in barley roots. Plants 2019, 8, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandin, A.; Lapointe, L.; Dizengremel, P. The alternative respiratory pathway allows sink to cope with changes in carbon availability in the sink-limited plant Erythronium americanum. J. Exp. Bot. 2009, 60, 4235–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, L.H.; Giraud, E.; Uggalla, V.; Lister, R.; Clifton, R.; Glen, A.; Thirkettle-Watts, D.; Van Aken, O.; Whelan, J. Identification of regulatory pathways controlling gene expression of stress-responsive mitochondrial proteins in Arabidopsis. Plant Physiol. 2008, 147, 1858–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative oxidase impacts the plant response to biotic stress by influencing the mitochondrial generation of reactive oxygen species. Plant Cell Environ. 2013, 36, 721–732. [Google Scholar] [CrossRef]
- Wagner, A.M. A role for active oxygen species as second messengers in the induction of alternative oxidase gene expression in Petunia hybrida cells. Febs Lett. 1995, 368, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Duan, J.; Li, H.; Liang, H.; Li, X.; Han, N. Alternative respiratory pathway under drought is partially mediated by hydrogen peroxide and contributes to antioxidant protection in wheat leaves. Plant Prod. Sci. 2008, 11, 59–66. [Google Scholar] [CrossRef]
- Sotirios, S.M.; Leondios, L.; George, P.; Evy, M.Z.; Georgios, A. Expression, purification, and physicochemical characterization of the N-terminal active site of human angiotensin-I converting enzyme. J. Pept. Sci. 2007, 13, 31–36. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5’ to 3’) |
|---|---|
| qHvAOX1a-F | GCAACGAACCTACAAGCGTG |
| qHvAOX1a-R | AAGAGCCCAGCACCAACAA |
| qHvAOX1d1-F | CCTCCCATTAGCTTTTCGACCAG |
| qHvAOX1d1-R | CGGTAGCACGTAACAGCGTGGACT |
| qHvAOX1d2-F | TACGACCACGAGTTTCGCGAGCA |
| qHvAOX1d2-R | GCTAAAGAGCCCTCATTTCCTC |
| HvMnSOD-F | CAGGTCGTACAACWCGATTA |
| HvMnSOD-R | CGTCAAGAAATCCAAACAGTC |
| HvFeSOD-F | GCAACGTTGGTACAACGGA |
| HvFeSOD-R | CGTAAAGAGCGTCATTTGG |
| HvPOD-F | GGTCCCATTACCTTTTCGTGGTC |
| HvPOD-R | GCCTAGCACGTAACACGCTGACT |
| HvCAT1-F | TAGCAGGACGAGTAACGCCTGGT |
| HvCAT1-R | CGTAAAGAGCCCTCTAATCG |
| HvCAT2-F | GCAACGAACCTACAACCGTC |
| HvCAT2-R | AAGAGCCCAGCACCAACAAT |
| HvAPX-F | GCTCCCATTAGCTTTTCGACAC |
| HvAPX-R | GCCTAGCACGTAACAGCGTTCA |
| HvACTIN-F | GTGGTCGTACAACWGGTATTGTG |
| HvACTIN-R | GCTCATCAAATCCAAACACTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Wang, X.; Na, X.; Feng, R.; He, Q.; Wang, S.; Liang, C.; Yan, L.; Zhou, L.; Bi, Y. Alternative Pathway Is Involved in Hydrogen Peroxide-Enhanced Cadmium Tolerance in Hulless Barley Roots. Plants 2021, 10, 2329. https://doi.org/10.3390/plants10112329
He L, Wang X, Na X, Feng R, He Q, Wang S, Liang C, Yan L, Zhou L, Bi Y. Alternative Pathway Is Involved in Hydrogen Peroxide-Enhanced Cadmium Tolerance in Hulless Barley Roots. Plants. 2021; 10(11):2329. https://doi.org/10.3390/plants10112329
Chicago/Turabian StyleHe, Li, Xiaomin Wang, Xiaofan Na, Ruijun Feng, Qiang He, Shengwang Wang, Cuifang Liang, Lili Yan, Libin Zhou, and Yurong Bi. 2021. "Alternative Pathway Is Involved in Hydrogen Peroxide-Enhanced Cadmium Tolerance in Hulless Barley Roots" Plants 10, no. 11: 2329. https://doi.org/10.3390/plants10112329