
AIP1, Encoding the Small Subunit of Acetolactate Synthase, Is Partially Responsible for Resistance to Hypoxic Stress in Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
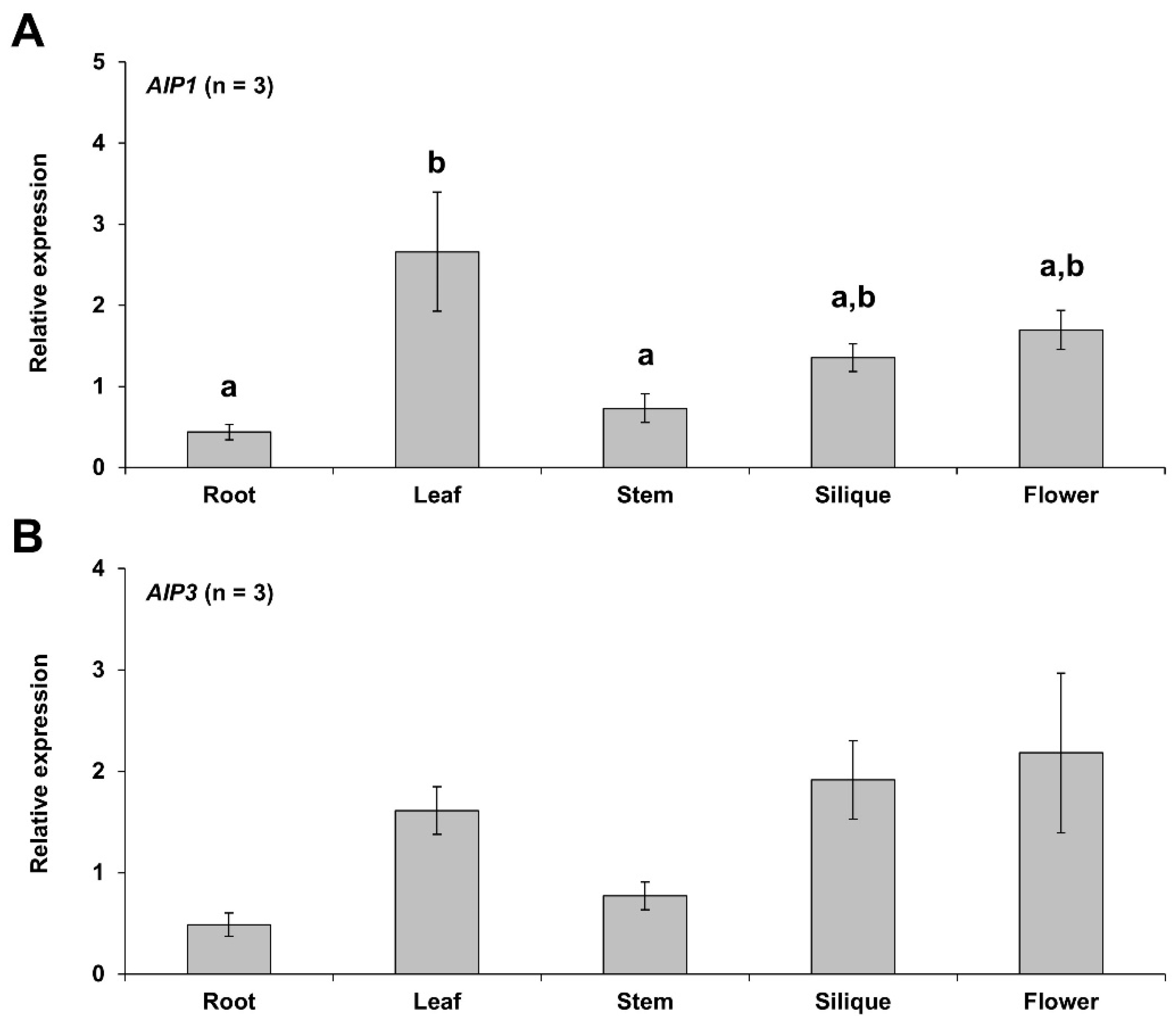
2.1. Organ-Specific Expression Patterns of AIP1 and AIP3 Genes
2.2. Expression Patterns of AIP1 and AIP3 under Submergence and Hypoxic Environments
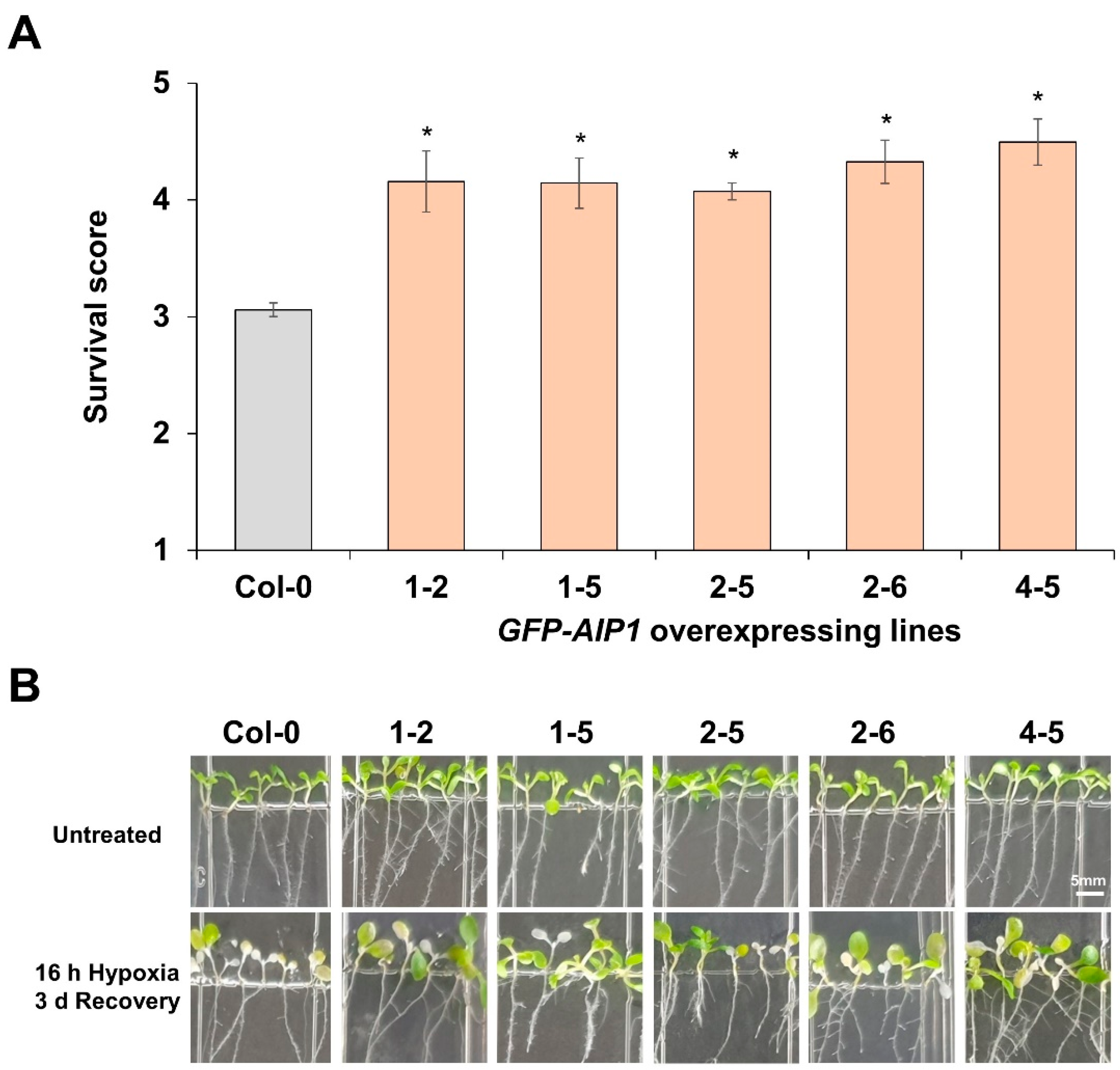
2.3. Assessment of the Tolerance to Hypoxia of AIP1-Overexpressing Lines by Recovery-Survival Rates
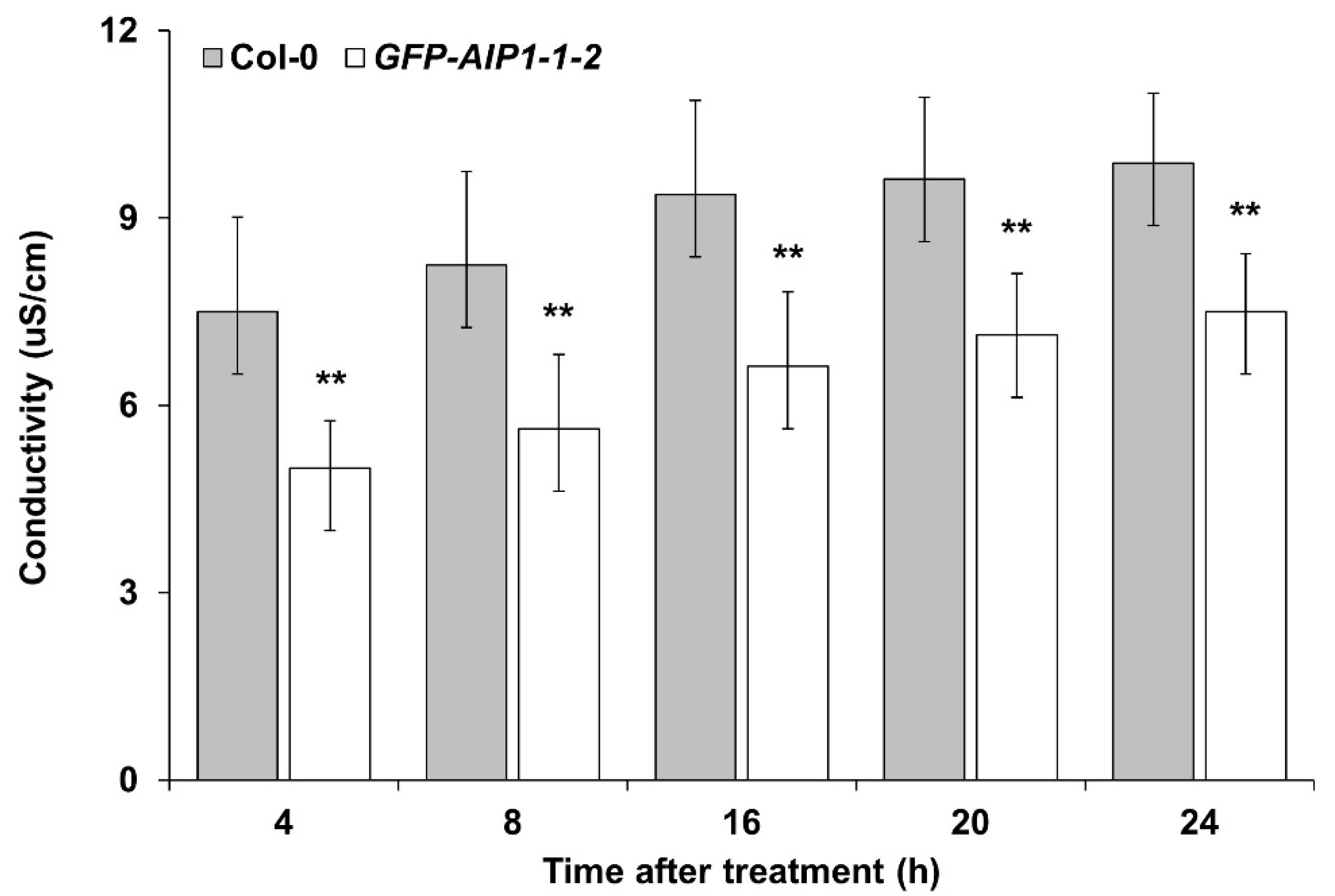
2.4. Assessment of the Tolerance to Hypoxia of an AIP1-Overexpressing Line by Electrolyte Leakage
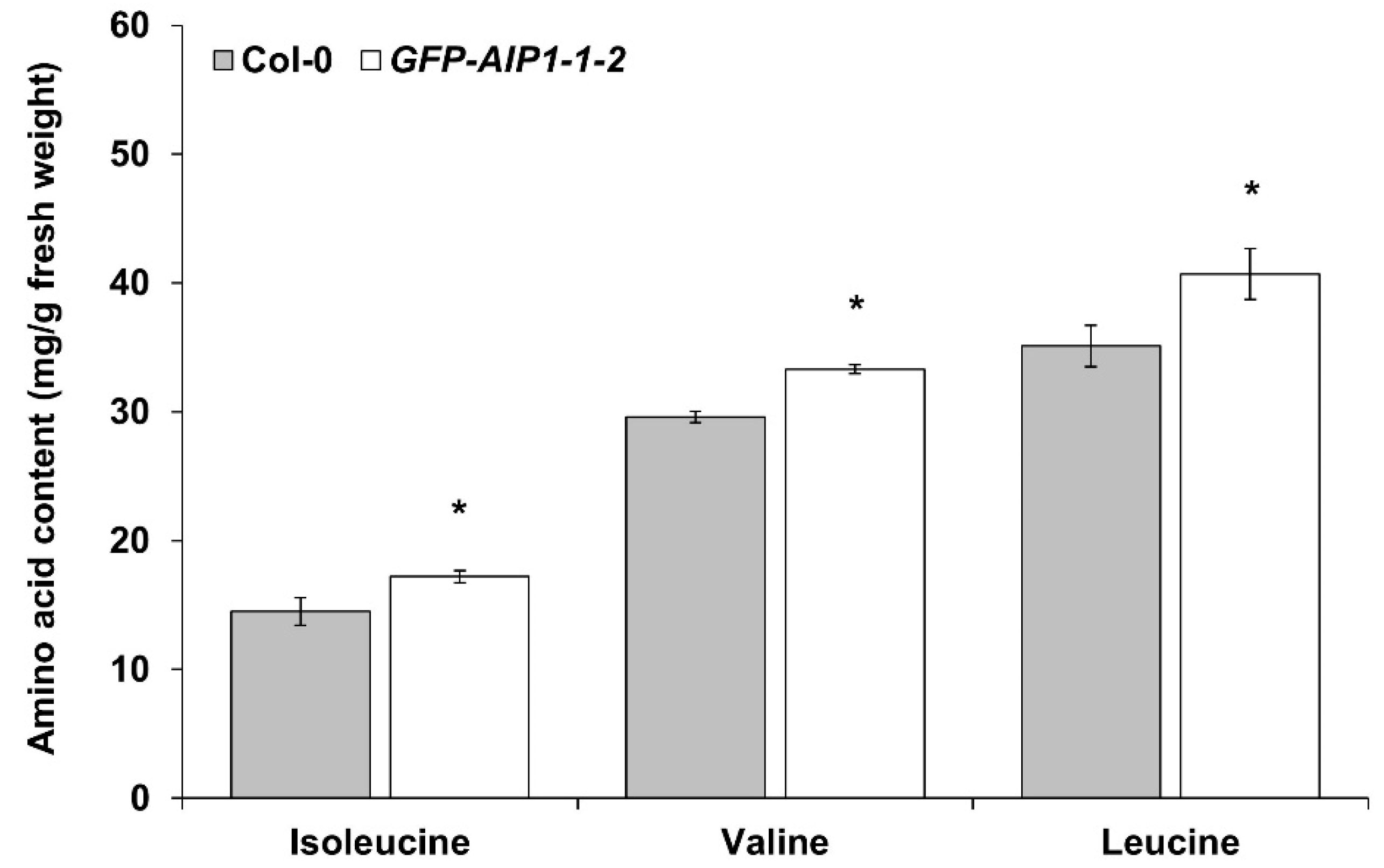
2.5. Measurement of BCAA Contents in a GFP-AIP1-Overexpressing Line
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. RNA Extraction and cDNA Synthesis
4.3. Tissue-Specific Expression of AIP1 and AIP3
4.4. Expression of AIP1 and AIP3 Genes upon Submergence and Hypoxia Conditions
4.5. Generation of AIP1-Overexpressing Arabidopsis Transformants
4.6. Assessment of Hypoxia Tolerance of AIP1 Transformants by Measuring Recovery Rates
4.7. Assessment of Tolerance to Hypoxia by AIP1 Transformants Based on Electrolyte Leakage Assay
4.8. Measurement of BCAA Contents in AIP1 Transformants
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically rethinking agriculture for the 21st century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drew, M.C. Plant injury and adaptation to oxygen deficiency in the root environment: A review. Plant Soil 1983, 75, 179–199. [Google Scholar] [CrossRef]
- Jackson, M.B.; Colmer, T.D. Response and adaptation by plants to flooding stress. Ann. Bot. 2005, 96, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Choi, D. Molecular events underlying coordinated hormone action in submergence escape response of deepwater rice. J. Plant Biol. 2011, 54, 365. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Holdsworth, M.J. Every breath you take: New insights into plant and animal oxygen sensing. Cell 2020, 180, 22–24. [Google Scholar] [CrossRef]
- Loreti, E.; Perata, P. The many facets of hypoxia in plants. Plants 2020, 9, 745. [Google Scholar] [CrossRef]
- Hwang, J.H.; Lee, M.O.; Choy, Y.H.; Ha-Lee, Y.M.; Hong, C.B.; Lee, D.H. Expression profile analysis of hypoxia responses in Arabidopsis roots and shoots. J. Plant Biol. 2011, 54, 373. [Google Scholar] [CrossRef]
- Hwang, S.T.; Kim, Y.L.; Sohn, S.H.; Choi, D. Gene expression profiling provides insight into the escape behavior of deepwater rice during submergence. J. Plant Biol. 2018, 61, 373–383. [Google Scholar] [CrossRef]
- Minami, A.; Yano, K.; Gamuyao, R.; Nagai, K.; Kuroha, T.; Ayano, M.; Nakamori, M.; Koike, M.; Kondo, Y.; Niimi, Y.; et al. Time-course transcriptomics analysis reveals key responses of submerged deepwater rice to flooding. Plant Physiol. 2018, 176, 3081–3102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streeter, J.G.; Thompson, J.F. Anaerobic accumulation of y-aminobutyric acid and alanine in radish leaves (Raphanus Sativus, L.). Plant Physiol. 1972, 49, 572–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reggiani, R.; Cantu, C.A.; Brambilla, I.; Bertani, A. Accumulation and interconversion of amino acids in rice roots under anoxia. Plant Cell Physiol. 1988, 29, 981–987. [Google Scholar]
- De Sousa, C.A.F.; Sodek, L. Alanine metabolism and alanine aminotransferase activity in soybean (Glycine Max) during hypoxia of the root System and subsequent return to normoxia. Environ. Exp. Bot. 2003, 50, 1–8. [Google Scholar] [CrossRef]
- Lee, D.J.; Chi, Y.T.; Kim, D.M.; Choi, S.H.; Lee, J.Y.; Choi, G.W. Ectopic Expression of CaRLK1 enhances hypoxia tolerance with increasing alanine production in Nicotiana Spp. Plant Mol. Biol. 2014, 86, 255–270. [Google Scholar] [CrossRef]
- Singh, B.K.; Shaner, D.L. Biosynthesis of branched chain amino acids: From test tube to field. Plant Cell 1995, 7, 935–944. [Google Scholar] [CrossRef]
- McCourt, J.A.; Pang, S.S.; King-Scott, J.; Guddat, L.W.; Duggleby, R.G. Herbicide-binding sites revealed in the structure of plant acetohydroxyacid synthase. Proc. Natl. Acad. Sci. USA 2006, 103, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y. Characterization of the Arabidopsis Glutamine Dumper1 Mutant Reveals Connections between Amino Acid Homeostasis and Plant Stress Responses. Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2015. [Google Scholar]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Binder, S. Branched-chain amino acid metabolism in Arabidopsis thaliana. Arab. Book 2010, 8, e0137. [Google Scholar] [CrossRef] [Green Version]
- Knill, T.; Schuster, J.; Reichelt, M.; Gershenzon, J.; Binder, S. Arabidopsis branched-chain aminotransferase 3 functions in both amino acid and glucosinolate biosynthesis. Plant Physiol. 2008, 146, 1028–1039. [Google Scholar] [CrossRef] [Green Version]
- Chipman, D.; Barak, Z.; Schloss, J.V. Biosynthesis of 2-aceto-2-hydroxy acids: Acetolactate synthases and acetohydroxyacid synthases. Biochim. Et Biophys. Acta—Protein Struct. Mol. Enzymol. 1998, 1385, 401–419. [Google Scholar] [CrossRef]
- Lee, Y.T.; Duggleby, R.G. Identification of the regulatory subunit of Arabidopsis thaliana acetohydroxyacid synthase and reconstitution with its catalytic subunit. Biochemistry 2001, 40, 6836–6844. [Google Scholar] [CrossRef]
- Lee, Y.T.; Duggleby, R.G. Regulatory interactions in Arabidopsis thaliana acetohydroxyacid synthase. FEBS Lett. 2002, 512, 180–184. [Google Scholar] [CrossRef] [Green Version]
- Gedi, V.; Yoon, M.Y. Bacterial acetohydroxyacid synthase and its inhibitors—A summary of their structure, biological activity and current status. FEBS J. 2012, 279, 946–963. [Google Scholar] [CrossRef] [PubMed]
- Dezfulian, M.H.; Foreman, C.; Jalili, E.; Pal, M.; Dhaliwal, R.K.; Roberto, D.K.; Imre, K.M.; Kohalmi, S.E.; Crosby, W.L. Acetolactate synthase regulatory subunits play divergent and overlapping roles in branched-chain amino acid synthesis and Arabidopsis development. BMC Plant Biol. 2017, 17, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggleby, R.G. Identification of an acetolactate synthase small subunit gene in two eukaryotes. Gene 1997, 190, 245–249. [Google Scholar] [CrossRef]
- Shimizu, M.; Fujii, T.; Masuo, S.; Takaya, N. Mechanism of de novo branched-chain amino acid synthesis as an alternative electron Sink in hypoxic Aspergillus nidulans cells. Appl. Environ. Microbiol. 2010, 76, 1507–1515. [Google Scholar] [CrossRef] [Green Version]
- Binder, S.; Knill, T.; Schuster, J. Branched-chain amino acid metabolism in higher plants. Physiol. Plant 2007, 129, 68–78. [Google Scholar] [CrossRef]
- Hershey, H.P.; Schwartz, L.J.; Gale, J.P.; Abell, L.M. Cloning and functional expression of the small subunit of acetolactate synthase from Nicotiana Plumbaginifolia. Plant Mol. Biol. 1999, 40, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.H.; Dennis, E.S.; Peacock, W.J. Arabidopsis roots and shoots have different mechanisms for hypoxic stress tolerance. Plant Physiol. 1999, 119, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Dolferus, R.; Ellis, M.; De Bruxelles, G.; Trevaskis, B.; Hoeren, F.; Dennis, E.S.; Peacock, W.J. Strategies of gene action in Arabidopsis during hypoxia. Ann. Bot. 1997, 79, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Ismond, K.P.; Dolferus, R.; De Pauw, M.; Dennis, E.S.; Good, A.G. Enhanced low oxygen survival in Arabidopsis through increased metabolic flux in the fermentative pathway. Plant Physiol. 2003, 132, 1292–1302. [Google Scholar] [CrossRef] [Green Version]
- Geigenberger, P. Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol. 2003, 6, 247–256. [Google Scholar] [CrossRef]
- Van Dongen, J.T.; Schurr, U.; Pfister, M.; Geigenberger, P. Phloem metabolism and function have to cope with low internal oxygen. Plant Physiol. 2003, 131, 1529–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. Characterization of AtHIGD1 Gene in Arabidopsis. Master’s Thesis, Kunsan National University, Gunsan-si, Jeollabuk-do (Miryong-dong), Korea, 2016. [Google Scholar]
- Hwang, S.T. Molecular Physiological Characterization of HIGD Genes in Rice and Arabidopsis under Submergence or Hypoxia. Ph.D. Thesis, Kunsan National University, Gunsan-si, Korea, 2016. [Google Scholar]
- Hwang, S.T.; Li, H.; Alavilli, H.; Lee, B.H.; Choi, D. Molecular and physiological characterization of AtHIGD1 in Arabidopsis. Biochem. Biophys. Res. Commun. 2017, 487, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Lee, S.C.; Isa, N.M.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatsugai, N.; Katagiri, F. Quantification of plant cell death by electrolyte leakage assay. Bio Protoc. 2018, 8, e2758. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Im, G.; Choi, D. AIP1, Encoding the Small Subunit of Acetolactate Synthase, Is Partially Responsible for Resistance to Hypoxic Stress in Arabidopsis thaliana. Plants 2021, 10, 2251. https://doi.org/10.3390/plants10112251
Im G, Choi D. AIP1, Encoding the Small Subunit of Acetolactate Synthase, Is Partially Responsible for Resistance to Hypoxic Stress in Arabidopsis thaliana. Plants. 2021; 10(11):2251. https://doi.org/10.3390/plants10112251
Chicago/Turabian StyleIm, Geunmuk, and Dongsu Choi. 2021. "AIP1, Encoding the Small Subunit of Acetolactate Synthase, Is Partially Responsible for Resistance to Hypoxic Stress in Arabidopsis thaliana" Plants 10, no. 11: 2251. https://doi.org/10.3390/plants10112251