Comparative Transcriptome Analysis Provides New Insights into the Molecular Regulatory Mechanism of Adventitious Root Formation in Ramie (Boehmeria nivea L.)

 , ,
, ,
Abstract
:1. Background
2. Methods
2.1. Ethics Statement
2.2. Plant Growth and Sampling
2.3. RNA Isolation, Library Preparation and Sequencing
2.4. Data Filtering and Mapping
2.5. Identification of Differentially Expressed Genes (DEGs)
2.6. Gene Ontology (GO) and Pathway Enrichment Analysis of DEGs
2.7. Quantitative PCR (qPCR) Validation of DEGs
3. Results
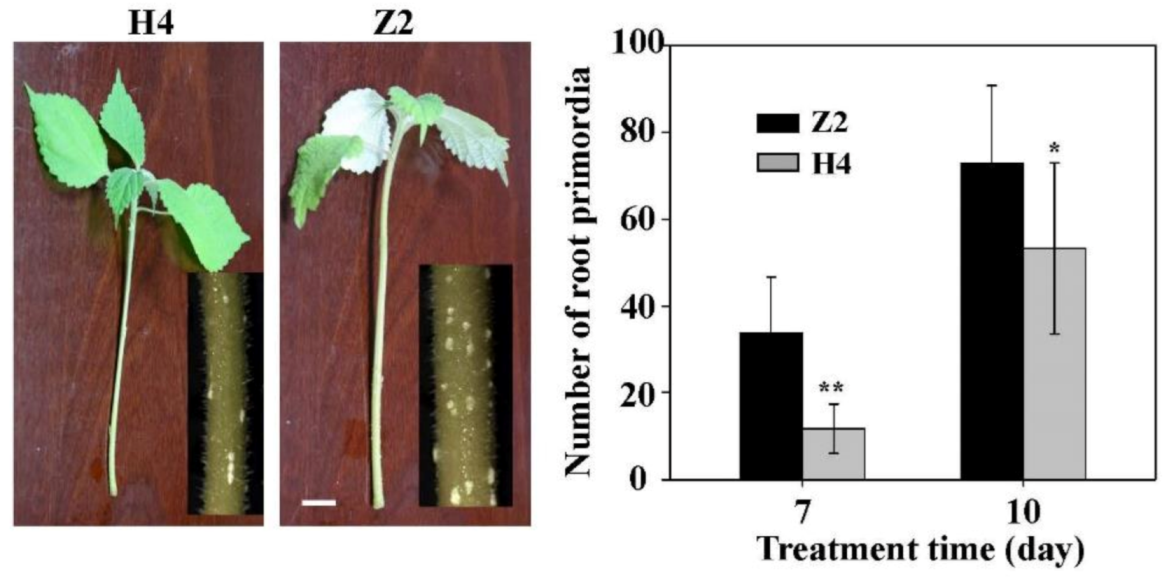
3.1. Adventitious Root Formation of Two Ramie Cultivars
3.2. Transcriptome Sequencing Analysis of Twelve cDNA Libraries
3.3. Comparison of Gene Expression Pattern between Z2 and H4
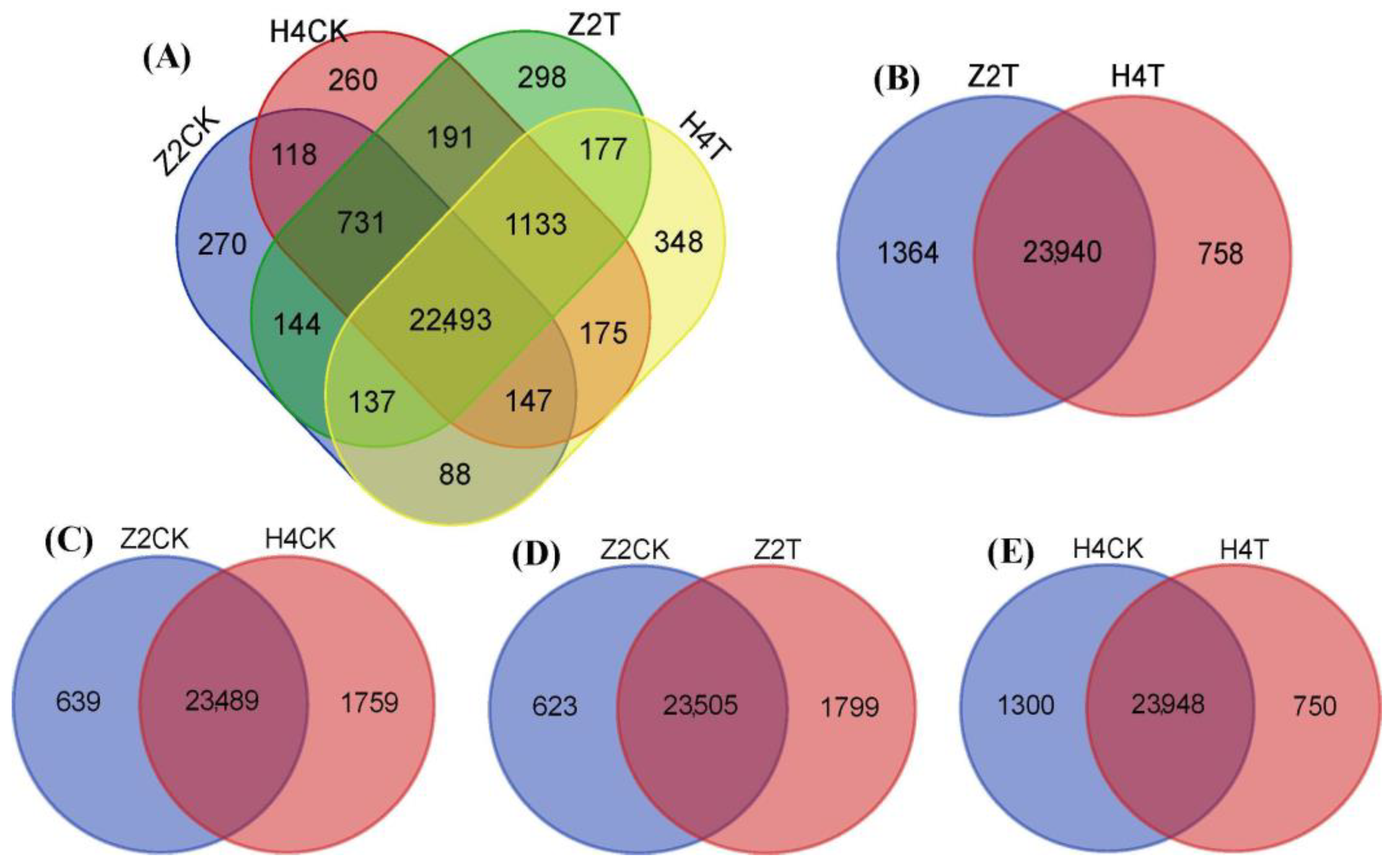
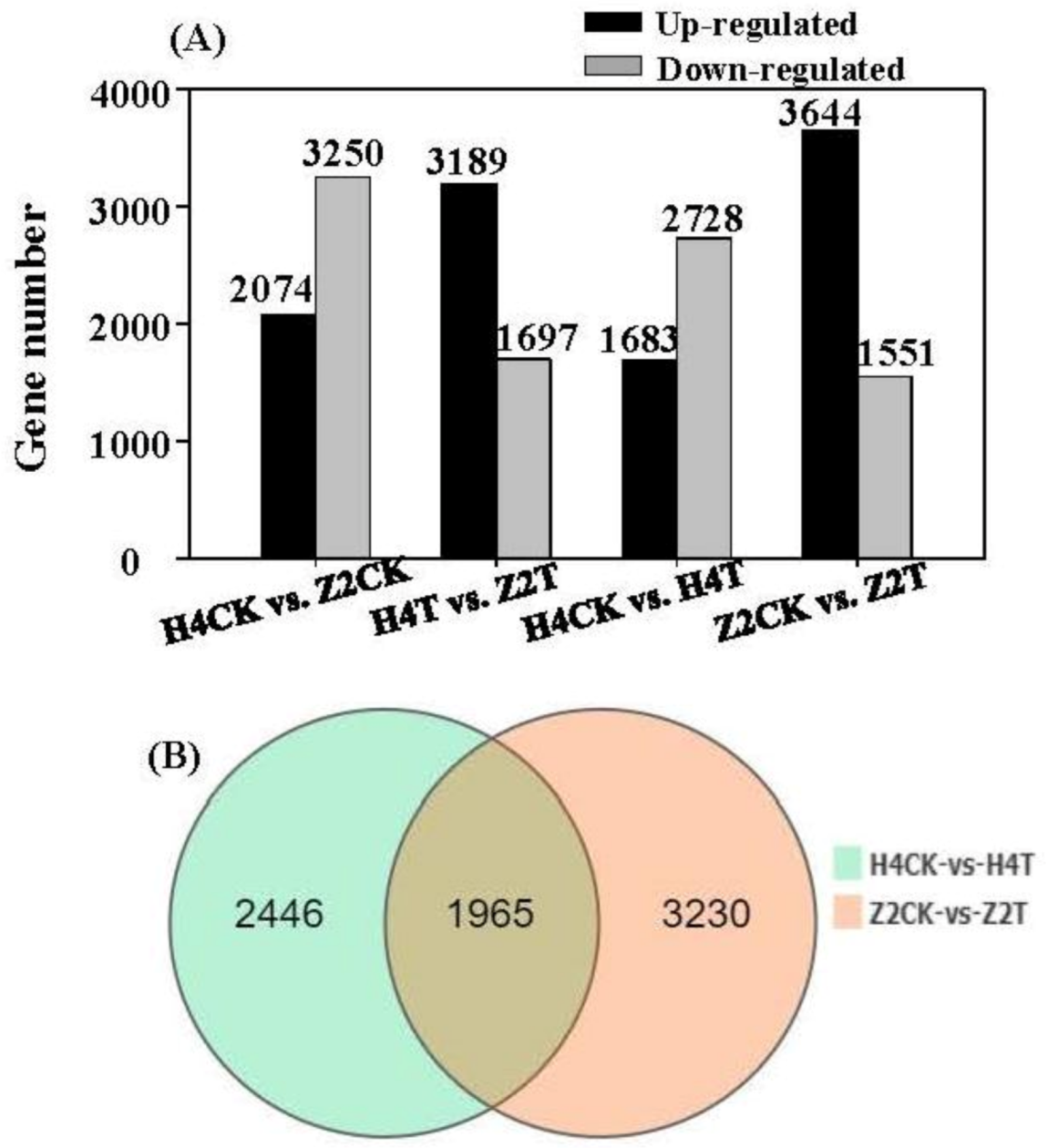
3.4. Identification and Expression of DEGs
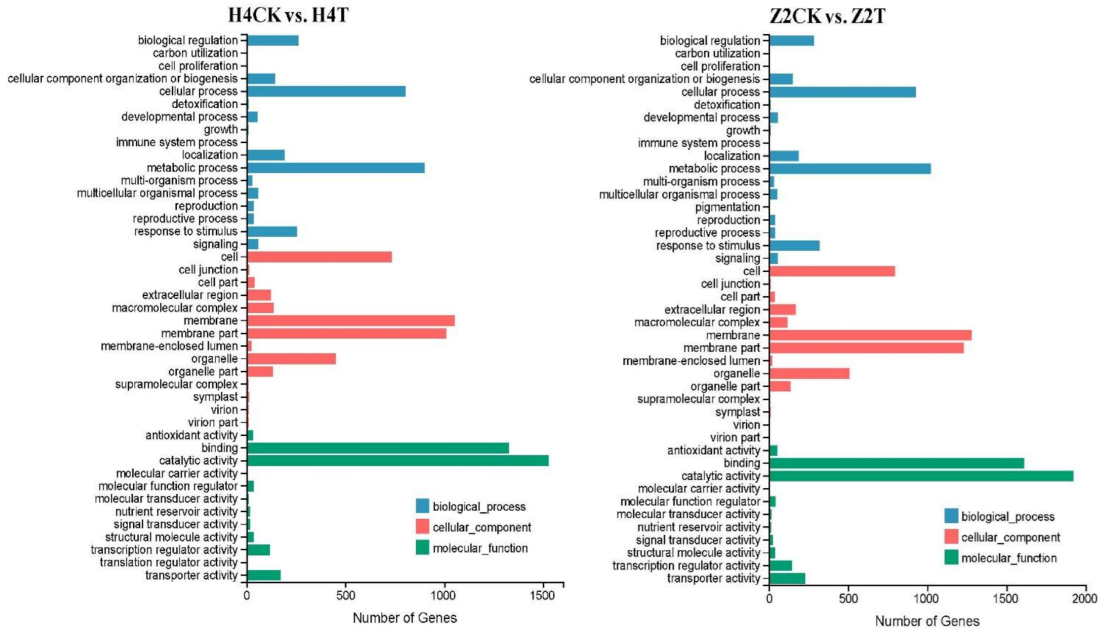
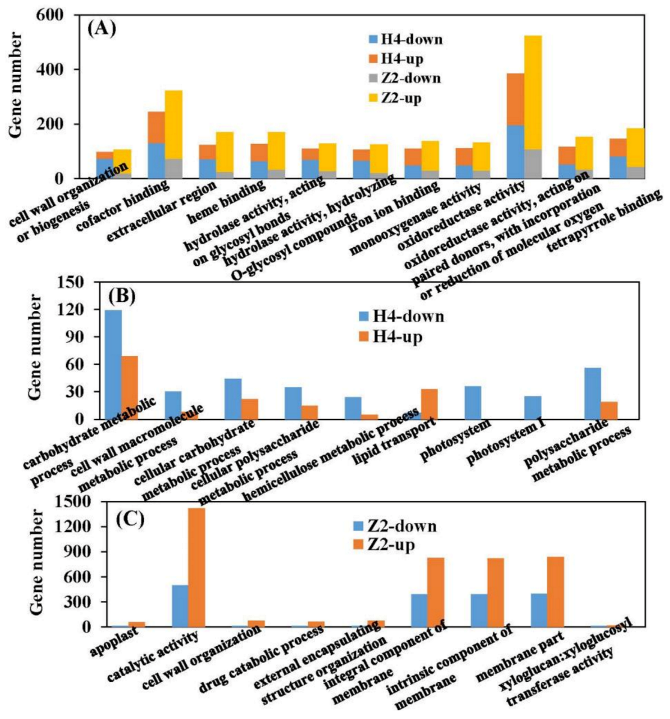
3.5. GO (Gene Ontology) Clustering of DEGs
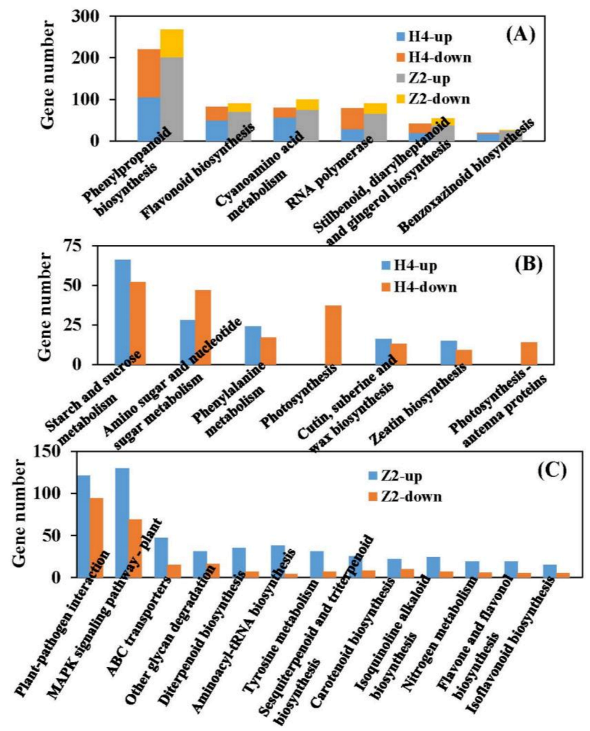
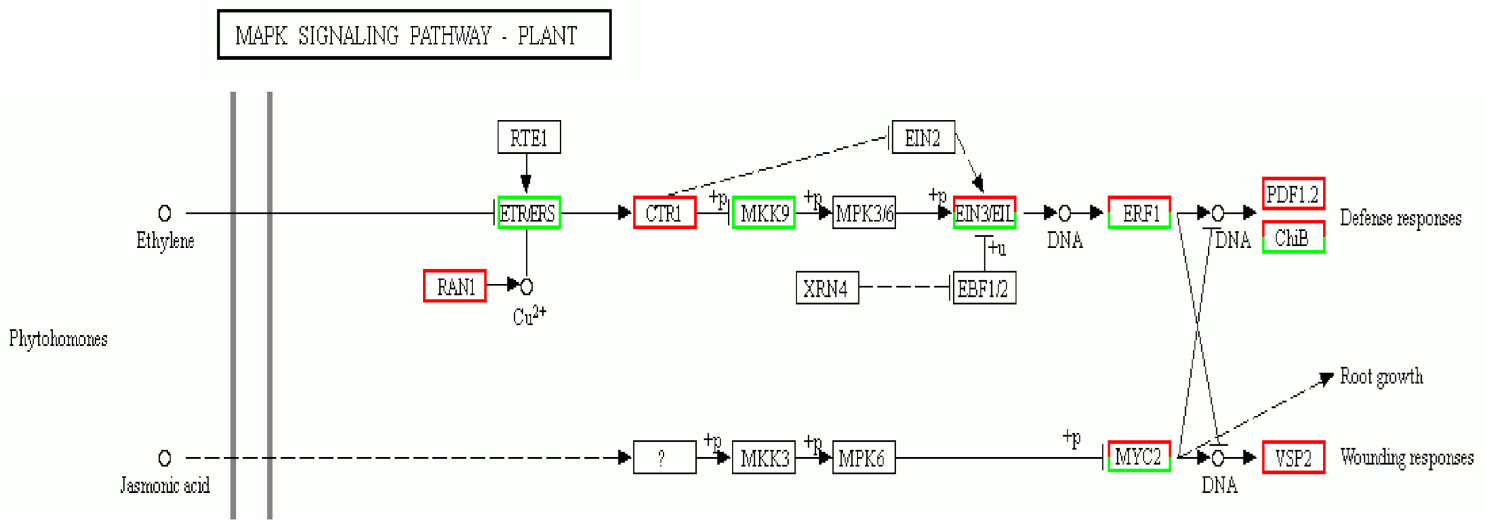
3.6. KEGG Pathway Analysis of DEGs
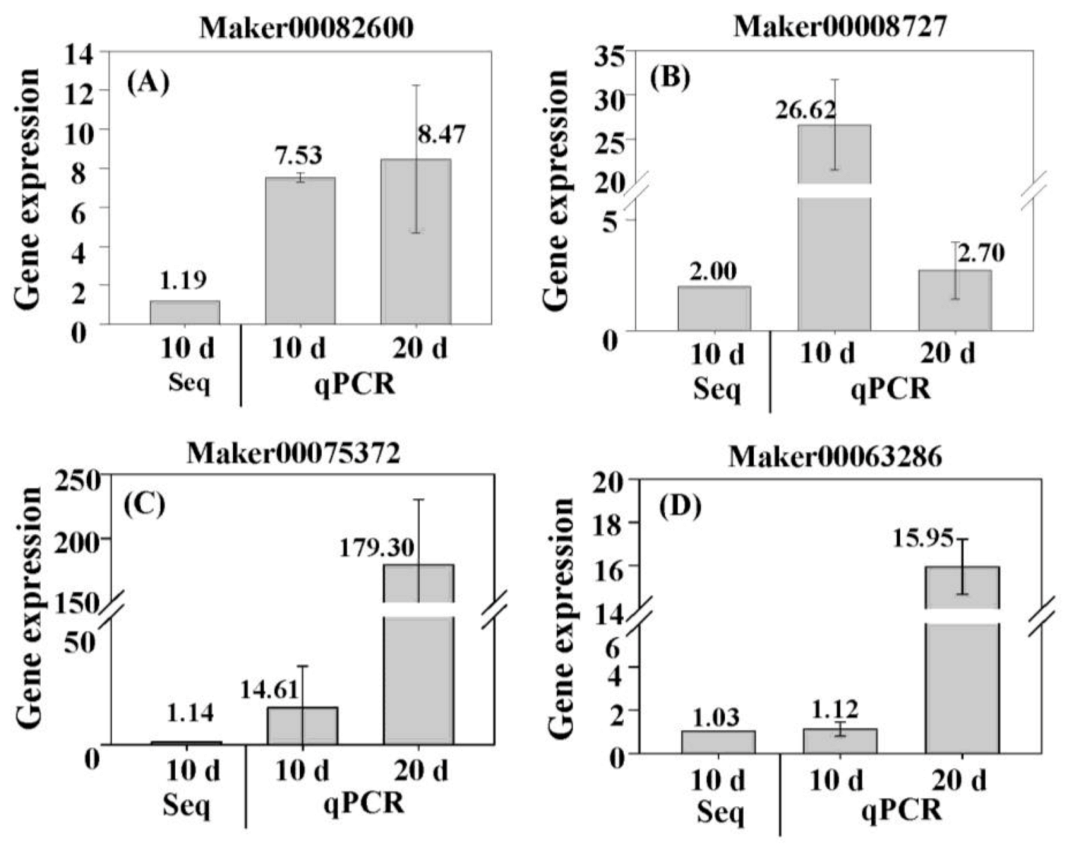
3.7. Verification of Expression of DEGs by qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AR | adventitious root |
| DEGs | differentially expressed genes |
| GO | gene ontology |
| JA | jasmonic acid |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| qPCR | quantitative PCR. |
References
- Vidoz, M.L.; Loreti, E.; Mensuali, A.; Alpi, A.; Perata, P. Hormonal interplay during adventitious root formation in flooded tomato plants. Plant J. 2010, 63, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Polanco, M.; Señorans, J.; Zwiazek, J.J. Role of adventitious roots in water relations of tamarack (Larix laricina) seedlings exposed to flooding. BMC Plant Biol. 2012, 12, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, K.W.; Allen, J.A.; Cahoon, D.R. Differential rates of vertical accretion and elevation change among aerial root types in Micronesian mangrove forests. Estuar. Coast. Shelf Sci. 2003, 56, 251–259. [Google Scholar] [CrossRef]
- Kipriotisa, E.; Xiong, H.P.; Vafeiadakisc, T.; Kipriotia, M.; Alexopoulou, E. Ramie and kenaf as feed crops. Ind. Crop Prod. 2015, 68, 126–130. [Google Scholar] [CrossRef]
- Zhu, Q.H.; Huang, D.Y.; Liu, S.L.; Rao, Z.X.; Cao, X.L.; Ren, X.F. Accumulation and subcellular distribution of cadmium in ramie (Boehmeria nivea L. gaud) planted on elevated soil cadmium contents. Plant Soil Environ. 2013, 59, 57–61. [Google Scholar]
- Luan, M.-B.; Jian, J.-B.; Chen, P.; Chen, J.-H.; Chen, J.-H.; Gao, Q.; Gao, G.; Zhou, J.-H.; Chen, K.-M.; Guang, X.-M.; et al. Draft genome sequence of ramie, Boehmeria nivea (L.) Gaudich. Mol. Ecol. Resour. 2018, 18, 639–645. [Google Scholar] [CrossRef]
- Mori, Y.; Miyahara, F.; Tsutsumi, Y.; Kondo, R. Effects of combinational treatment with ethephon and indole-3-butyric acid on adventitious rooting of Pinus thunbergii cuttings. Plant Growth Regul. 2011, 63, 271–278. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.-Y.; Wu, L.-Y.; Zhang, C.-C.; Li, H.-L.; Tan, L.-Q.; Cao, H.-L.; Cheng, H. Transcriptome Analysis of Indole-3-Butyric Acid-Induced Adventitious Root Formation in Nodal Cuttings of Camellia sinensis (L.). PLoS ONE 2014, 9, e107201. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.S.; Alam, M.J.; Mollick, A.S.; Khan, M.N.I. Rooting of cuttings of the wild Indian almond tree (Sterculia foetida) enhanced by the application of indole-3-butyric acid (IBA) under leafy and non-leafy conditions. Rhizosphere 2018, 5, 8–15. [Google Scholar] [CrossRef]
- Pacurar, D.I.; Perrone, I.; Bellini, C. Auxin is a central player in the hormone cross-talks that control adventitious rooting. Physiol. Plantarum. 2014, 151, 83–96. [Google Scholar] [CrossRef]
- Li, X.-P.; Xu, Q.-Q.; Liao, W.-B.; Ma, Z.-J.; Xu, X.-T.; Wang, M.; Ren, P.-J.; Niu, L.-J.; Jin, X.; Zhu, Y.-C. Hydrogen peroxide is involved in abscisic acid-induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Biol. 2016, 59, 536–548. [Google Scholar] [CrossRef]
- Negi, S.; Sukumar, P.; Liu, X.; Cohen, J.D.; Muday, G.K. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010, 61, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druege, U.; Franken, P.; Lischewski, S.; Ahkami, A.H.; Zerche, S.; Hause, B.; Hajirezaei, M.R. Transcriptomic analysis reveals ethylene as stimulator and auxin as regulator of adventitious root formation in petunia cuttings. Front. Plant Sci. 2014, 5, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauriat, M.; Petterle, A.; Bellini, C.; Moritz, T. Gibberellins inhibit adventitious rooting in hybrid aspen and Arabidopsis by affecting auxin transport. Plant J. 2014, 78, 372–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baque, M.A.; Hahn, E.J.; Paek, K.Y. Growth, secondary metabolite production and antioxidant enzyme response of Morinda citrifolia adventitious root as affected by auxin and cytokinin. Plant Biotechnol. Rep. 2010, 4, 109–116. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, D.; Meng, Y.; Li, K.; Wang, H.; Han, M. Inhibition of adventitious root development in apple rootstocks by cytokinin is based on its suppression of adventitious root primordia formation. Physiol. Plantarum. 2019, 166, 663–676. [Google Scholar] [CrossRef]
- Rasmussen, A.; Mason, M.G.; De Cuyper, C.; Brewer, P.B.; Herold, S.; Agusti, J.; Geelen, D.; Greb, T.; Goormachtig, S.; Beeckman, T.; et al. Strigolactones Suppress Adventitious Rooting in Arabidopsis and Pea. Plant Physiol. 2012, 158, 1976–1987. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Tao, J.; Hou, M.; Huang, S.; Chen, S.; Liang, Z.; Xie, T.; Wei, Y.; Xie, X.; Yoneyama, K.; et al. A strigolactone signal is required for adventitious root formation in rice. Ann. Bot. 2015, 115, 1155–1162. [Google Scholar] [CrossRef]
- Xu, X.W.; Ji, J.; Ma, X.T.; Xu, Q.; Qi, X.H.; Chen, X.H. Comparative proteomic analysis provides insight into the key proteins involved in cucumber (Cucumis sativus L.) adventitious root emergence under waterlogging stress. Front. Plant Sci. 2016, 7, 1515. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Zhang, H.N.; Wu, Q.S.; Muday, G.K. Transcriptional sequencing and analysis of major genes involved in the adventitious root formation of mango cotyledon segments. Planta 2017, 245, 1193–1213. [Google Scholar] [CrossRef]
- Li, K.; Liang, Y.; Xing, L.; Mao, J.; Liu, Z.; Dong, F.; Meng, Y.; Han, M.; Zhao, C.; Bao, L.; et al. Transcriptome Analysis Reveals Multiple Hormones, Wounding and Sugar Signaling Pathways Mediate Adventitious Root Formation in Apple Rootstock. Int. J. Mol. Sci. 2018, 19, 2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.K.; Feng, Z.X.; Wang, X.; Wang, X.W.; Zhang, X.G. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Luan, M.; Xiong, H.; Chen, P.; Chen, J.; Gao, G.; Huang, K.; Zhu, A.; Yu, C. Genome-wide association study discovered favorable single nucleotide polymorphisms and candidate genes associated with ramet number in ramie (Boehmeria nivea L.). BMC Plant Biol. 2018, 18, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402e408. [Google Scholar] [CrossRef]
- Carpenter, W.J.; Cornell, J.A. Auxin application duration and concentration govern rooting of hibiscus stem cuttings. J. Am. Soc. Hort. Sci. 1992, 117, 68–74. [Google Scholar] [CrossRef]
- Roycewicz, P.S.; Malamy, J.E. Cell wall properties play an important role in the emergence of lateral root primordia from the parent root. J. Exp. Bot. 2014, 65, 2057–2069. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.D.; Dennison, W.C.; Alberte, R.S. Role of Seagrass Photosynthesis in Root Aerobic Processes. Plant Physiol. 1984, 74, 1055–1058. [Google Scholar] [CrossRef] [Green Version]
- Rapaka, V.K.; Bessler, B.; Schreiner, M.; Druege, U. Interplay between initial carbohydrate availability, current photosynthesis, and adventitious root formation in Pelargonium cuttings. Plant Sci. 2005, 168, 1547–1560. [Google Scholar] [CrossRef]
- Brinker, M. Microarray Analyses of Gene Expression during adventitious root development in Pinus contort. Plant Physiol. 2004, 135, 1526–1539. [Google Scholar] [CrossRef] [Green Version]
- Negishi, N.; Nakahama, K.; Urata, N.; Kojima, M.; Sakakibara, H.; Kawaoka, A. Hormone level analysis on adventitious root formation in Eucalyptus globulus. New For. 2014, 45, 577–587. [Google Scholar] [CrossRef]
- Kurepin, L.; Haslam, T.; Lopez-Villalobos, A.; Oinam, G.; Yeung, E. Adventitious root formation in ornamental plants: II. The role of plant growth regulators. Propag. Ornam. Plants 2011, 11, 161–171. [Google Scholar]
- Strader, L.C.; Bartel, B. Transport and metabolism of the endogenous auxin precursor indole-3-butyric acid. Mol. Plant 2011, 4, 477–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlicht, M.; Ludwig-Müller, J.; Burbach, C.; Volkmann, D.; Baluska, F. Indole-3-butyric acid induces lateral root formation via peroxisome-derived indole-3-acetic acid and nitric oxide. New Phytol. 2013, 200, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Rӧnsch, H.; Adam, G.; Matschke, J.; Schachler, G. Influence of (22S,23S)-homobrassinolide on rooting capacity and survival of adult Norway spruce cuttings. Tree Physiol. 1993, 12, 71–80. [Google Scholar] [CrossRef]
- Yang, W.; Zhu, C.H.; Ma, X.L.; Li, G.J.; Gan, L.J.; Ng, D.; Xia, K. Hydrogen peroxide is a second messenger in the salicylic acid-triggered adventitious rooting process in mung bean seedlings. PLoS ONE 2013, 8, e84580. [Google Scholar] [CrossRef] [Green Version]
- Steffens, B.; Wang, J.X.; Sauter, M. Interactions between ethylene, gibberellin and abscisic acid regulate emergence and growth rate of adventitious roots in deepwater rice. Planta 2006, 223, 604–612. [Google Scholar] [CrossRef]
- Laplaze, L.; Benkova, E.; Casimiro, I.; Maes, L.; Vanneste, S.; Swarup, R.; Weijers, D.; Calvo, V.; Parizot, B.; Herrera-Rodriguez, M.B.; et al. Cytokinins Act Directly on Lateral Root Founder Cells to Inhibit Root Initiation. Plant Cell 2007, 19, 3889–3900. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.D.; Chen, L.J.; Yu, S.M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar]
- Guan, J.C.; Koch, K.E.; Suzuki, M.; Wu, S.; Latshaw, S.; Petruff, T.; Goulet, C.; Klee, H.J.; McCarty, D.R. Diverse Roles of Strigolactone Signaling in Maize Architecture and the Uncoupling of a Branching-Specific Subnetwork. Plant Physiol. 2012, 160, 1303–1317. [Google Scholar] [CrossRef] [Green Version]
- Ravnikar, M.; Vilhar, B.; Gogala, N. Stimulatory effects of jasmonic acid on potato stem node and protoplast culture. J. Plant Growth Regul. 1992, 11, 29–33. [Google Scholar] [CrossRef]
- Lischewski, S.; Muchow, A.; Guthörl, D.; Hause, B. Jasmonates act positively in adventitious root formation in petunia cuttings. BMC Plant Biol. 2015, 15, 229. [Google Scholar]
- Gutierrez, L.; Mongelard, G.; Floková, K.; Păcurar, D.I.; Novák, O.; Staswick, P.; Kowalczyk, M.; Păcurar, M.; Demailly, H.; Geiss, G.; et al. Auxin Controls Arabidopsis Adventitious Root Initiation by Regulating Jasmonic Acid Homeostasis. Plant Cell 2012, 24, 2515–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onkokesung, N.; Ivan, G.; von Dahl, C.C.; Matsuoka, K.; Saluz, H.P.; Baldwin, I.T. Jasmonic acid and ethylene modulate local responses to wounding and simulated herbivory in Nicotiana attenuata leaves. Plant Physiol. 2010, 153, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Raw Read (M) | Total Clean Read (M) | Total Clean Base (Gb) | Clean Read q20 (%) | Clean Read q30 (%) | Total Mapping Genome Ratio (%) | Uniquely Mapping Genome Ratio (%) |
|---|---|---|---|---|---|---|---|
| H4T1 | 41.10 | 36.03 | 5.44 | 97.78 | 93.58 | 90.51 | 63.70 |
| H4T2 | 47.88 | 42.30 | 6.39 | 97.84 | 93.73 | 90.35 | 63.64 |
| H4T3 | 39.33 | 34.14 | 5.15 | 97.70 | 93.39 | 90.23 | 63.27 |
| H4CK1 | 45.76 | 40.17 | 6.06 | 97.81 | 93.70 | 90.37 | 65.14 |
| H4CK2 | 50.74 | 44.39 | 6.70 | 97.78 | 93.59 | 90.80 | 63.98 |
| H4CK3 | 45.36 | 39.87 | 6.02 | 97.80 | 93.62 | 90.67 | 64.03 |
| Z2T1 | 47.07 | 40.79 | 6.16 | 97.69 | 93.37 | 90.92 | 63.61 |
| Z2T2 | 44.86 | 39.44 | 5.96 | 97.81 | 93.64 | 90.47 | 65.14 |
| Z2T3 | 44.13 | 38.43 | 5.80 | 97.73 | 93.47 | 91.33 | 65.72 |
| Z2CK1 | 38.17 | 33.09 | 5.00 | 97.67 | 93.31 | 90.87 | 65.43 |
| Z2CK2 | 43.06 | 37.19 | 5.62 | 97.65 | 93.27 | 91.00 | 65.50 |
| Z2CK3 | 40.74 | 35.38 | 5.34 | 97.70 | 93.41 | 91.43 | 66.06 |
| Mean | 44.02 | 38.44 | 5.80 | 97.75 | 93.51 | 90.75 | 64.60 |
| Total | 528.20 | 461.22 | 69.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Guo, B.; Yu, C.; Chen, P.; Chen, J.; Gao, G.; Wang, X.; Zhu, A. Comparative Transcriptome Analysis Provides New Insights into the Molecular Regulatory Mechanism of Adventitious Root Formation in Ramie (Boehmeria nivea L.). Plants 2021, 10, 160. https://doi.org/10.3390/plants10010160
Chen K, Guo B, Yu C, Chen P, Chen J, Gao G, Wang X, Zhu A. Comparative Transcriptome Analysis Provides New Insights into the Molecular Regulatory Mechanism of Adventitious Root Formation in Ramie (Boehmeria nivea L.). Plants. 2021; 10(1):160. https://doi.org/10.3390/plants10010160
Chicago/Turabian StyleChen, Kunmei, Bing Guo, Chunming Yu, Ping Chen, Jikang Chen, Gang Gao, Xiaofei Wang, and Aiguo Zhu. 2021. "Comparative Transcriptome Analysis Provides New Insights into the Molecular Regulatory Mechanism of Adventitious Root Formation in Ramie (Boehmeria nivea L.)" Plants 10, no. 1: 160. https://doi.org/10.3390/plants10010160