
Evolutionary Change in Gut Specification in Caenorhabditis Centers on the GATA Factor ELT-3 in an Example of Developmental System Drift

Abstract
:1. Introduction

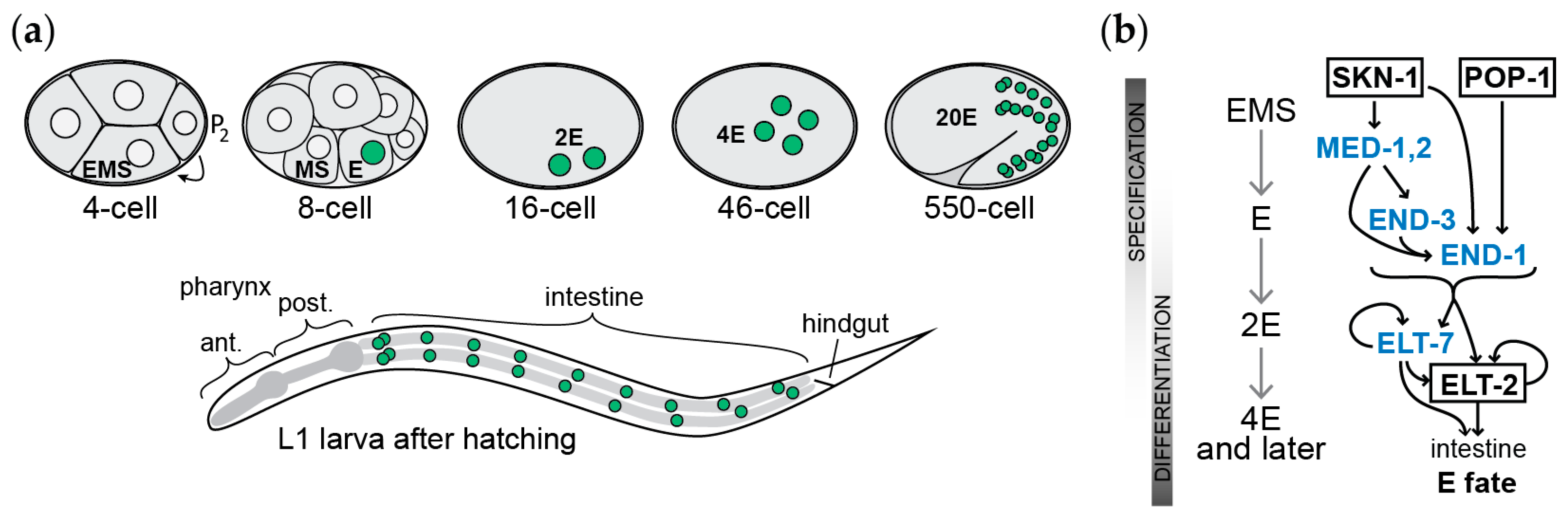
2. The Gut in C. elegans
3. Elucidation of the Gut Specification Network: Maternal Factors
4. Elucidation of the Gut Specification Network: Zygotic Factors
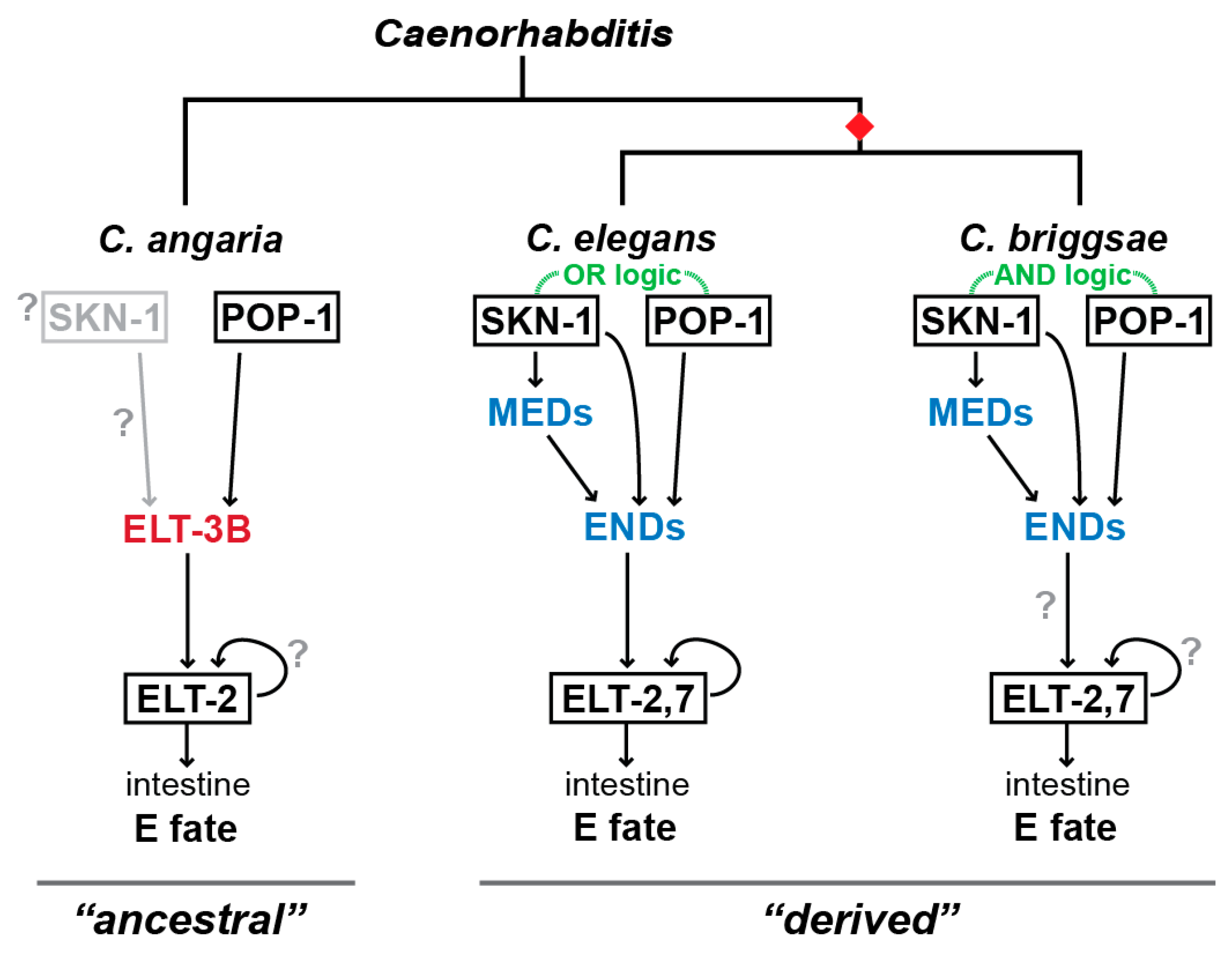
5. Evolution of Gut Specification in C. briggsae
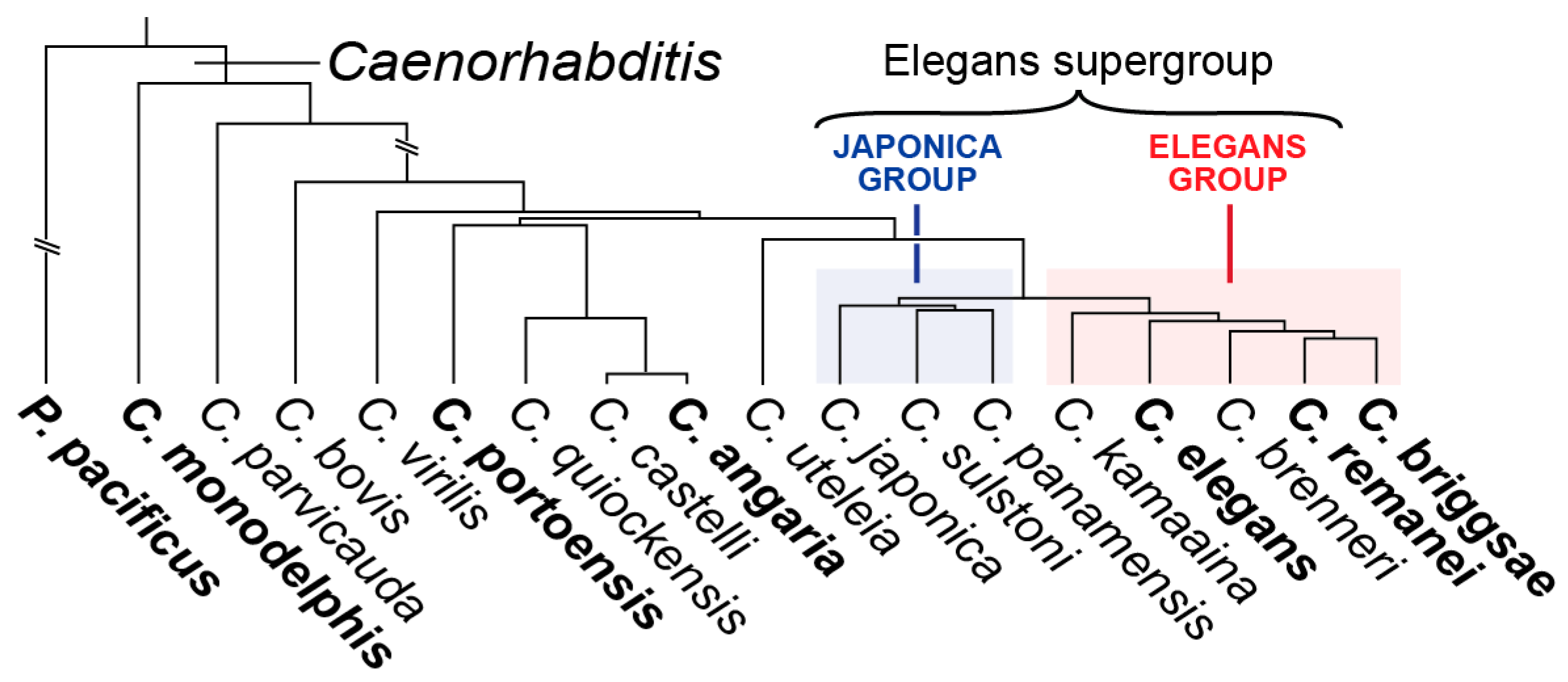
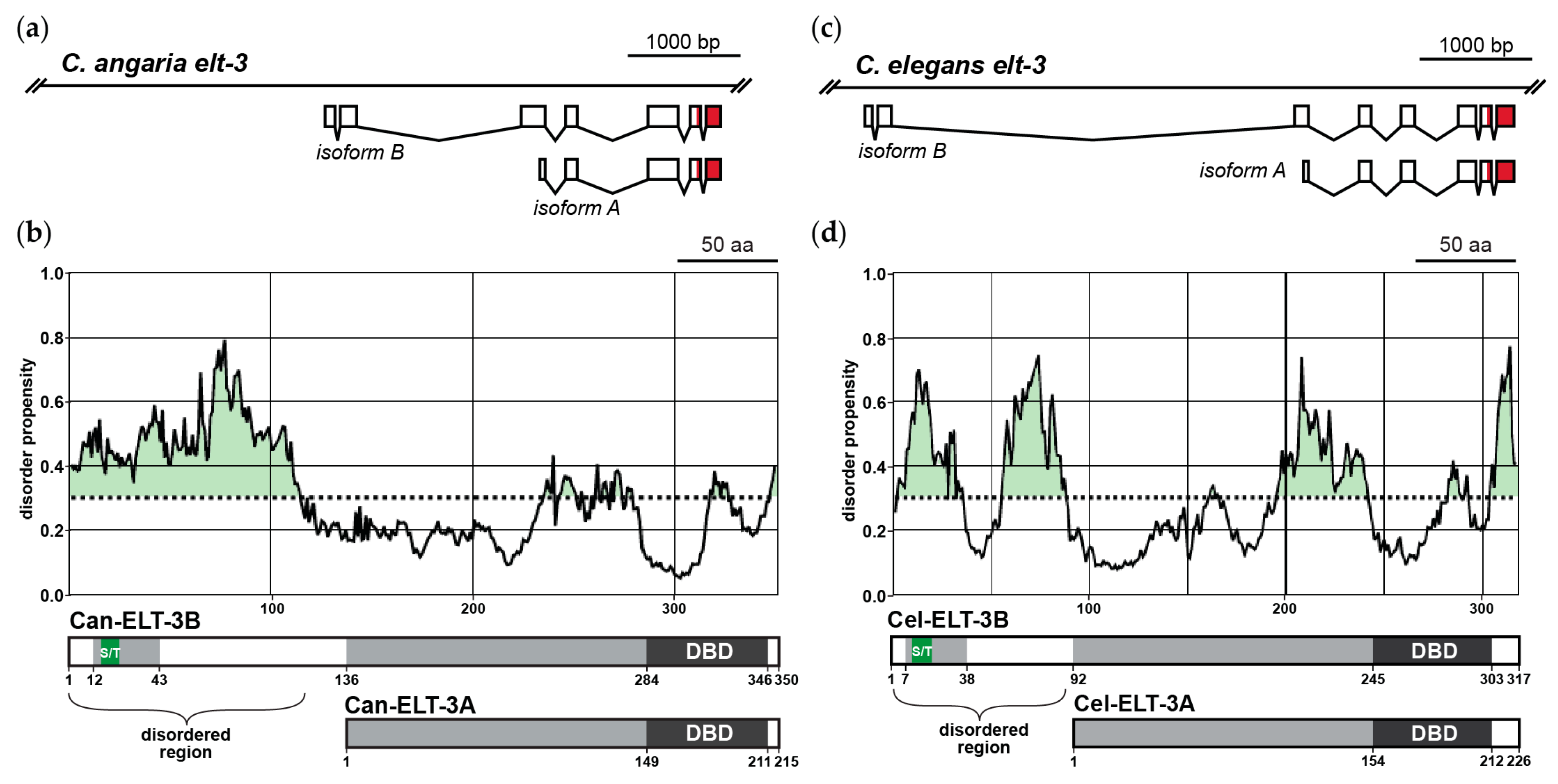
6. Gut Specification Outside of the Elegans Supergroup by the ELT-3 GATA Factor
7. Short and Long Isoforms of ELT-3 Have Different Functions
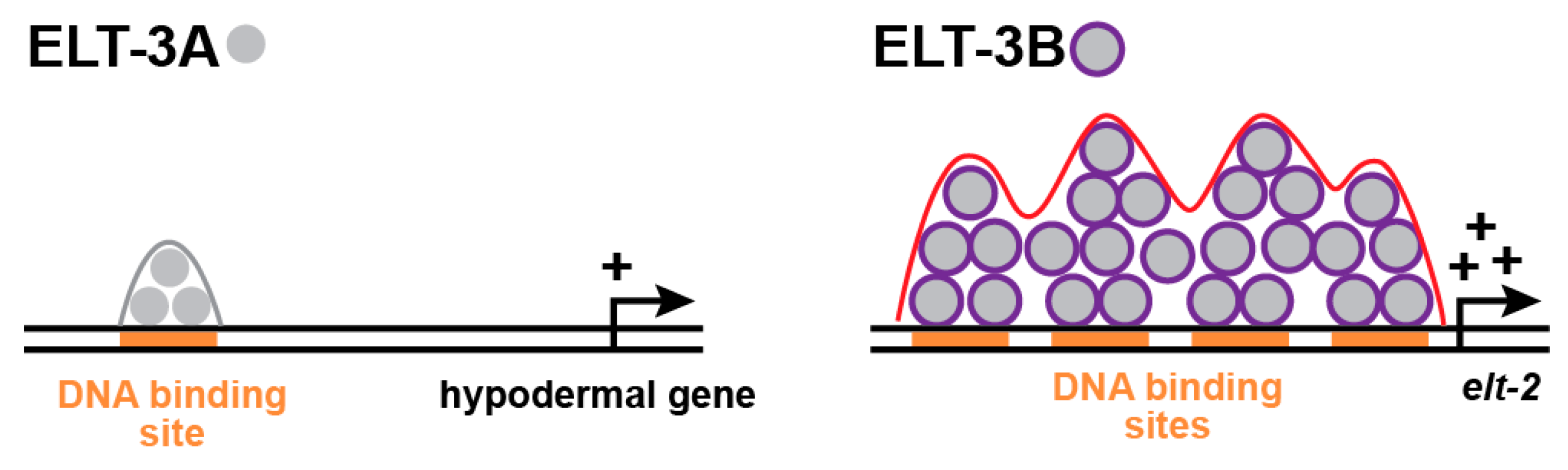
8. How Do ELT-3B and ELT-3A Regulate Different Target Genes?
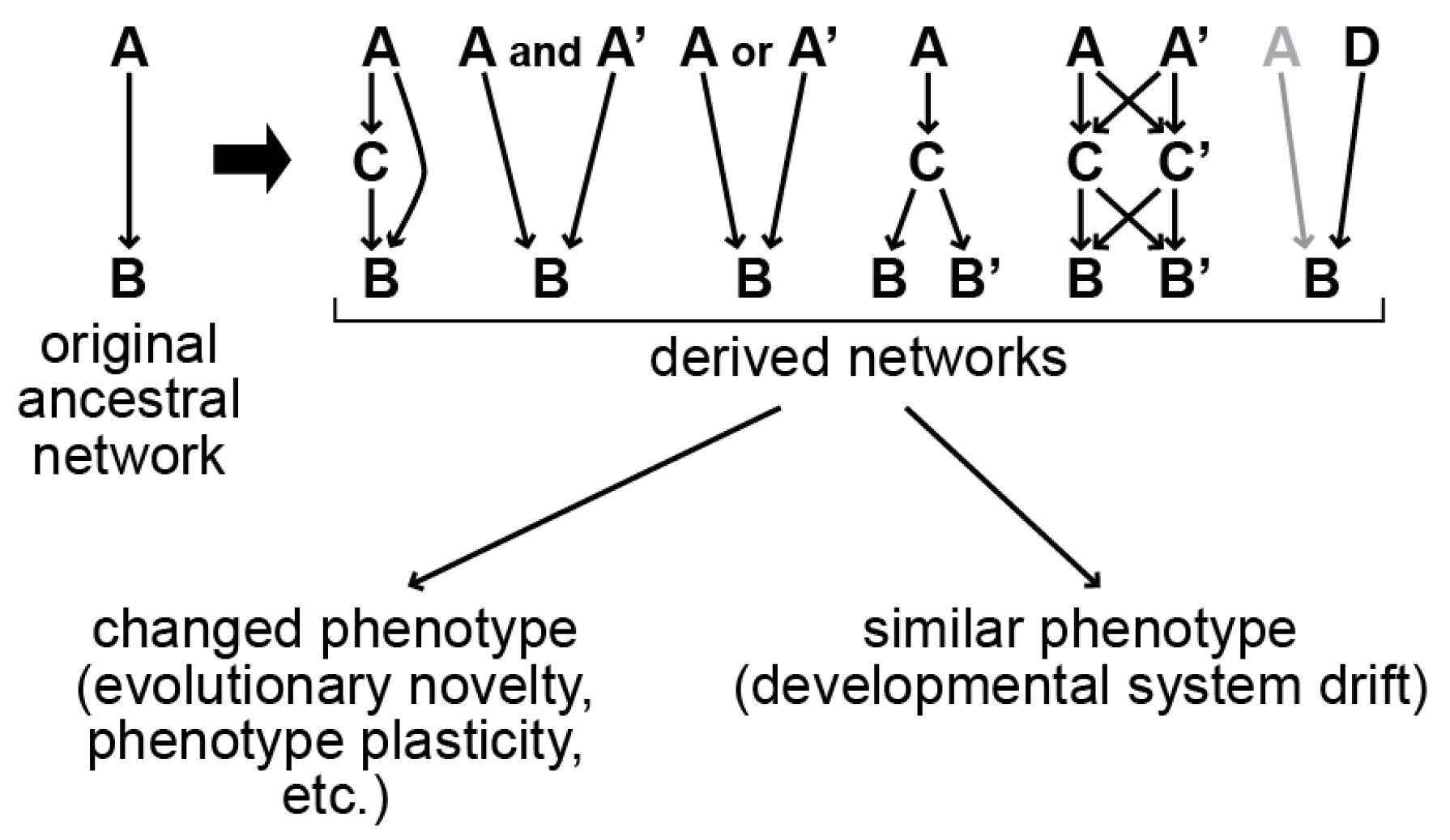
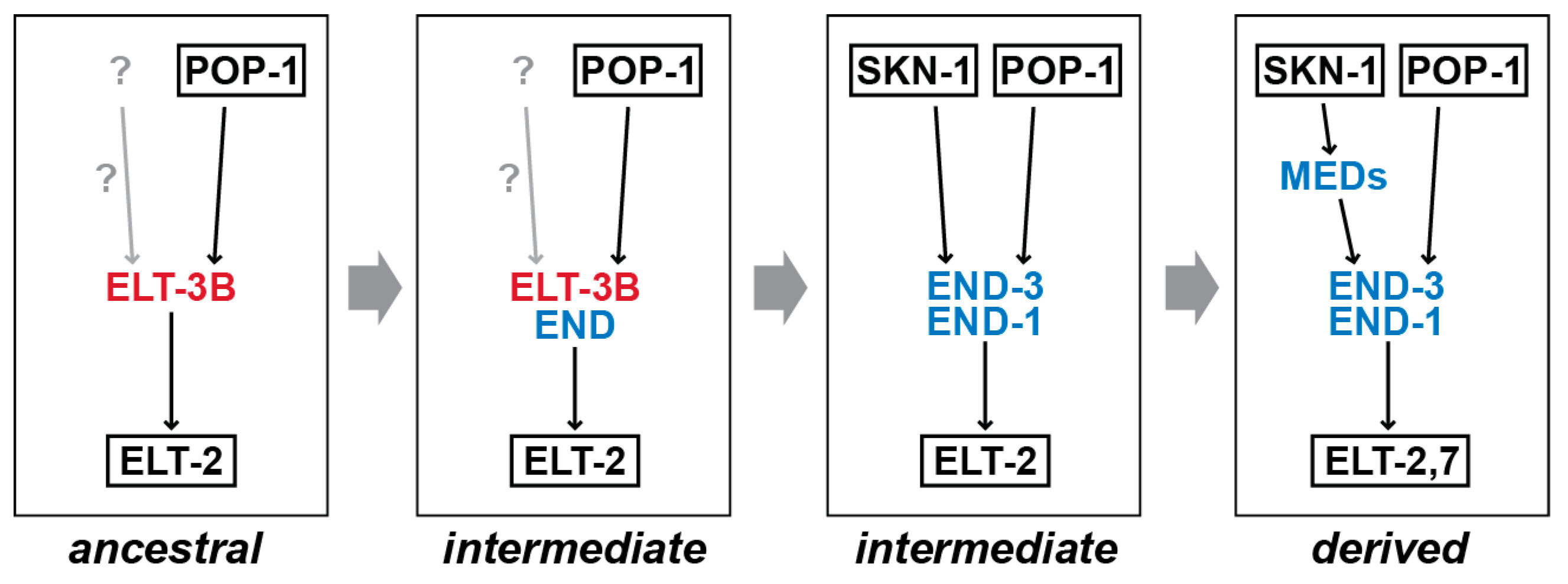
9. How Did the Gut Specification Network Change in Caenorhabditis?
10. Examples of DSD in Caenorhabditis and Other Animals
11. Additional Questions for Future Work
11.1. Gut Specification in Other Species in the Genus
11.2. Function of ELT-3B in C. Elegans and Other Species in the Elegans Supergroup
11.3. Evolution of ELT-3 Orthologues and Isoforms
11.4. Amenability of Gut Specification to Rewiring
11.5. What Caused the Rewiring of Endoderm Specification?
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davidson, E.H.; Levine, M.S. Properties of developmental gene regulatory networks. Proc. Natl. Acad. Sci. USA 2008, 105, 20063–20066. [Google Scholar] [CrossRef]
- Erwin, D.H.; Davidson, E.H. The evolution of hierarchical gene regulatory networks. Nat. Rev. Genet. 2009, 10, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittelmann, S.; Preger-Ben Noon, E.; McGregor, A.P.; Frankel, N. A complex gene regulatory architecture underlies the development and evolution of cuticle morphology in Drosophila. Curr. Opin. Genet. Dev. 2021, 69, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Chevin, L.M.; Leung, C.; Le Rouzic, A.; Uller, T. Using phenotypic plasticity to understand the structure and evolution of the genotype-phenotype map. Genetica 2022, 150, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Minelli, A. Phenotypic plasticity in development and evolution: Facts and concepts. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2010, 365, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- True, J.R.; Haag, E.S. Developmental system drift and flexibility in evolutionary trajectories. Evol. Dev. 2001, 3, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Haag, E.S. The same but different: Worms reveal the pervasiveness of developmental system drift. PLoS Genet. 2014, 10, e1004150. [Google Scholar] [CrossRef] [Green Version]
- Haag, E.S.; Fitch, D.H.A.; Delattre, M. From “the Worm” to “the Worms” and Back Again: The Evolutionary Developmental Biology of Nematodes. Genetics 2018, 210, 397–433. [Google Scholar] [CrossRef] [Green Version]
- Broitman-Maduro, G.; Sun, S.; Kikuchi, T.; Maduro, M.F. The GATA factor ELT-3 specifies endoderm in Caenorhabditis angaria in an ancestral gene network. Development 2022, 149, dev200984. [Google Scholar] [CrossRef]
- Coroian, C.; Broitman-Maduro, G.; Maduro, M.F. Med-type GATA factors and the evolution of mesendoderm specification in nematodes. Dev. Biol. 2005, 289, 444–455. [Google Scholar] [CrossRef] [Green Version]
- Eurmsirilerd, E.; Maduro, M.F. Evolution of Developmental GATA Factors in Nematodes. J. Dev. Biol. 2020, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.T.; Broitman-Maduro, G.; Hung, W.W.; Cervantes, S.; Maduro, M.F. Knockdown of SKN-1 and the Wnt effector TCF/POP-1 reveals differences in endomesoderm specification in C. briggsae as compared with C. elegans. Dev. Biol. 2009, 325, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maduro, M.; Hill, R.J.; Heid, P.J.; Newman-Smith, E.D.; Zhu, J.; Priess, J.; Rothman, J. Genetic redundancy in endoderm specification within the genus Caenorhabditis. Dev. Biol. 2005, 284, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Maduro, M.F. Structure and evolution of the C. elegans embryonic endomesoderm network. Biochim. Et Biophys. Acta 2009, 1789, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Maduro, M.F. Evolutionary Dynamics of the SKN-1→MED→END-1,3 Regulatory Gene Cascade in Caenorhabditis Endoderm Specification. G3 2020, 10, 333–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owraghi, M.; Broitman-Maduro, G.; Luu, T.; Roberson, H.; Maduro, M.F. Roles of the Wnt effector POP-1/TCF in the C. elegans endomesoderm specification gene network. Dev. Biol. 2010, 340, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Sulston, J.E.; Schierenberg, E.; White, J.G.; Thomson, J.N. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 1983, 100, 64–119. [Google Scholar] [CrossRef]
- Bowerman, B. Maternal control of pattern formation in early Caenorhabditis elegans embryos. Curr. Top. Dev. Biol. 1998, 39, 73–117. [Google Scholar]
- Maduro, M.F. Cell fate specification in the C. elegans embryo. Dev. Dyn. 2010, 239, 1315–1329. [Google Scholar] [CrossRef]
- Goldstein, B. Induction of gut in Caenorhabditis elegans embryos. Nature 1992, 357, 255–257. [Google Scholar] [CrossRef]
- Asan, A.; Raiders, S.A.; Priess, J.R. Morphogenesis of the C. elegans Intestine Involves Axon Guidance Genes. PLoS Genet. 2016, 12, e1005950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Broitman-Maduro, G.; Maduro, M.F. Partially compromised specification causes stochastic effects on gut development in C. elegans. Dev. Biol. 2017, 427, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.U.; Son, M.; Kim, J.; Shim, Y.H.; Kawasaki, I. CDC-25.2, a C. elegans ortholog of cdc25, is essential for the progression of intestinal divisions. Cell Cycle 2016, 15, 654–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, P.A.; Levine, M. Mosaic and regulative development: Two faces of one coin. Curr. Biol. CB 2006, 16, R236–R239. [Google Scholar] [CrossRef] [Green Version]
- Bowerman, B.; Eaton, B.A.; Priess, J.R. skn-1, a maternally expressed gene required to specify the fate of ventral blastomeres in the early C. elegans embryo. Cell 1992, 68, 1061–1075. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Thompson, S.; Priess, J.R. pop-1 encodes an HMG box protein required for the specification of a mesoderm precursor in early C. elegans embryos. Cell 1995, 83, 599–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowerman, B.; Draper, B.W.; Mello, C.C.; Priess, J.R. The maternal gene skn-1 encodes a protein that is distributed unequally in early C. elegans embryos. Cell 1993, 74, 443–452. [Google Scholar] [CrossRef]
- Shetty, P.; Lo, M.C.; Robertson, S.M.; Lin, R. C. elegans TCF protein, POP-1, converts from repressor to activator as a result of Wnt-induced lowering of nuclear levels. Dev. Biol. 2005, 285, 584–592. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, T.K.; Steinbaugh, M.J.; Hourihan, J.M.; Ewald, C.Y.; Isik, M. SKN-1/Nrf, stress responses, and aging in Caenorhabditis elegans. Free. Radic. Biol. Med. 2015, 88, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocheleau, C.E.; Downs, W.D.; Lin, R.; Wittmann, C.; Bei, Y.; Cha, Y.H.; Ali, M.; Priess, J.R.; Mello, C.C. Wnt signaling and an APC-related gene specify endoderm in early C. elegans embryos. Cell 1997, 90, 707–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocheleau, C.E.; Yasuda, J.; Shin, T.H.; Lin, R.; Sawa, H.; Okano, H.; Priess, J.R.; Davis, R.J.; Mello, C.C. WRM-1 activates the LIT-1 protein kinase to transduce anterior/posterior polarity signals in C. elegans. Cell 1999, 97, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, A.; Shelton, C.A.; Maloof, J.N.; Meneghini, M.; Bowerman, B. Wnt pathway components orient a mitotic spindle in the early Caenorhabditis elegans embryo without requiring gene transcription in the responding cell. Genes Dev. 1999, 13, 2028–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, T.H.; Yasuda, J.; Rocheleau, C.E.; Lin, R.; Soto, M.; Bei, Y.; Davis, R.J.; Mello, C.C. MOM-4, a MAP kinase kinase kinase-related protein, activates WRM-1/LIT-1 kinase to transduce anterior/posterior polarity signals in C. elegans. Mol. Cell. 1999, 4, 275–280. [Google Scholar] [CrossRef]
- Thorpe, C.J.; Schlesinger, A.; Carter, J.C.; Bowerman, B. Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm. Cell 1997, 90, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Bhambhani, C.; Ravindranath, A.J.; Mentink, R.A.; Chang, M.V.; Betist, M.C.; Yang, Y.X.; Koushika, S.P.; Korswagen, H.C.; Cadigan, K.M. Distinct DNA binding sites contribute to the TCF transcriptional switch in C. elegans and Drosophila. PLoS Genet. 2014, 10, e1004133. [Google Scholar] [CrossRef] [Green Version]
- Maduro, M.F.; Kasmir, J.J.; Zhu, J.; Rothman, J.H. The Wnt effector POP-1 and the PAL-1/Caudal homeoprotein collaborate with SKN-1 to activate C. elegans endoderm development. Dev. Biol. 2005, 285, 510–523. [Google Scholar] [CrossRef] [Green Version]
- Sullivan-Brown, J.L.; Tandon, P.; Bird, K.E.; Dickinson, D.J.; Tintori, S.C.; Heppert, J.K.; Meserve, J.H.; Trogden, K.P.; Orlowski, S.K.; Conlon, F.L.; et al. Identifying Regulators of Morphogenesis Common to Vertebrate Neural Tube Closure and Caenorhabditis elegans Gastrulation. Genetics 2016, 202, 123–139. [Google Scholar] [CrossRef]
- Calvo, D.; Victor, M.; Gay, F.; Sui, G.; Luke, M.P.; Dufourcq, P.; Wen, G.; Maduro, M.; Rothman, J.; Shi, Y. A POP-1 repressor complex restricts inappropriate cell type-specific gene transcription during Caenorhabditis elegans embryogenesis. Embo. J. 2001, 20, 7197–7208. [Google Scholar] [CrossRef] [Green Version]
- Witze, E.S.; Field, E.D.; Hunt, D.F.; Rothman, J.H. C. elegans pur alpha, an activator of end-1, synergizes with the Wnt pathway to specify endoderm. Dev. Biol. 2009, 327, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Wiesenfahrt, T.; Duanmu, J.; Snider, F.; Moerman, D.; Au, V.; Li-Leger, E.; Flibotte, S.; Parker, D.M.; Marshall, C.J.; Nishimura, E.O.; et al. A Strategy To Isolate Modifiers of Caenorhabditis elegans Lethal Mutations: Investigating the Endoderm Specifying Ability of the Intestinal Differentiation GATA Factor ELT-2. G3 2018, 8, 1425–1437. [Google Scholar] [CrossRef]
- Ewe, C.K.; Torres Cleuren, Y.N.; Flowers, S.E.; Alok, G.; Snell, R.G.; Rothman, J.H. Natural cryptic variation in epigenetic modulation of an embryonic gene regulatory network. Proc. Natl. Acad. Sci. USA 2020, 117, 13637–13646. [Google Scholar] [CrossRef] [PubMed]
- Mello, C.C.; Draper, B.W.; Krause, M.; Weintraub, H.; Priess, J.R. The pie-1 and mex-1 genes and maternal control of blastomere identity in early C. elegans embryos. Cell 1992, 70, 163–176. [Google Scholar] [CrossRef]
- Mello, C.C.; Schubert, C.; Draper, B.; Zhang, W.; Lobel, R.; Priess, J.R. The PIE-1 protein and germline specification in C. elegans embryos. Nature 1996, 382, 710–712. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, R.; Weigner, C.; Hutter, H.; Feichtinger, R.; Schnabel, H. mex-1 and the general partitioning of cell fate in the early C. elegans embryo. Mech. Dev. 1996, 54, 133–147. [Google Scholar] [CrossRef]
- Maduro, M.F.; Meneghini, M.D.; Bowerman, B.; Broitman-Maduro, G.; Rothman, J.H. Restriction of mesendoderm to a single blastomere by the combined action of SKN-1 and a GSK-3beta homolog is mediated by MED-1 and -2 in C. elegans. Mol. Cell. 2001, 7, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Gilleard, J.S.; Shafi, Y.; Barry, J.D.; McGhee, J.D. ELT-3: A Caenorhabditis elegans GATA factor expressed in the embryonic epidermis during morphogenesis. Dev. Biol. 1999, 208, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Sommermann, E.M.; Strohmaier, K.R.; Maduro, M.F.; Rothman, J.H. Endoderm development in Caenorhabditis elegans: The synergistic action of ELT-2 and -7 mediates the specification→differentiation transition. Dev. Biol. 2010, 347, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Fukushige, T.; Hawkins, M.G.; McGhee, J.D. The GATA-factor elt-2 is essential for formation of the Caenorhabditis elegans intestine. Dev. Biol. 1998, 198, 286–302. [Google Scholar] [CrossRef] [Green Version]
- Baugh, L.R.; Hill, A.A.; Slonim, D.K.; Brown, E.L.; Hunter, C.P. Composition and dynamics of the Caenorhabditis elegans early embryonic transcriptome. Development 2003, 130, 889–900. [Google Scholar] [CrossRef] [Green Version]
- Tintori, S.C.; Osborne Nishimura, E.; Golden, P.; Lieb, J.D.; Goldstein, B. A Transcriptional Lineage of the Early C. elegans Embryo. Dev. Cell 2016, 38, 430–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimshony, T.; Feder, M.; Levin, M.; Hall, B.K.; Yanai, I. Spatiotemporal transcriptomics reveals the evolutionary history of the endoderm germ layer. Nature 2015, 519, 219–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Hill, R.J.; Heid, P.J.; Fukuyama, M.; Sugimoto, A.; Priess, J.R.; Rothman, J.H. end-1 encodes an apparent GATA factor that specifies the endoderm precursor in Caenorhabditis elegans embryos. Genes Dev. 1997, 11, 2883–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maduro, M.F.; Broitman-Maduro, G.; Mengarelli, I.; Rothman, J.H. Maternal deployment of the embryonic SKN-1→MED-1,2 cell specification pathway in C. elegans. Dev. Biol. 2007, 301, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, M.G.; McGhee, J.D. elt-2, a second GATA factor from the nematode Caenorhabditis elegans. J. Biol. Chem. 1995, 270, 14666–14671. [Google Scholar] [CrossRef] [Green Version]
- Fukushige, T.; Hendzel, M.J.; Bazett-Jones, D.P.; McGhee, J.D. Direct visualization of the elt-2 gut-specific GATA factor binding to a target promoter inside the living Caenorhabditis elegans embryo. Proc. Natl. Acad. Sci. USA 1999, 96, 11883–11888. [Google Scholar] [CrossRef]
- Du, L.; Tracy, S.; Rifkin, S.A. Mutagenesis of GATA motifs controlling the endoderm regulator elt-2 reveals distinct dominant and secondary cis-regulatory elements. Dev. Biol. 2016, 412, 160–170. [Google Scholar] [CrossRef]
- Wiesenfahrt, T.; Berg, J.Y.; Nishimura, E.O.; Robinson, A.G.; Goszczynski, B.; Lieb, J.D.; McGhee, J.D. The Function and Regulation of the GATA Factor ELT-2 in the C. elegans Endoderm. Development 2015, 143, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Fukushige, T.; Goszczynski, B.; Tian, H.; McGhee, J.D. The Evolutionary Duplication and Probable Demise of an Endodermal GATA Factor in Caenorhabditis elegans. Genetics 2003, 165, 575–588. [Google Scholar] [CrossRef]
- Lowry, J.A.; Atchley, W.R. Molecular evolution of the GATA family of transcription factors: Conservation within the DNA-binding domain. J. Mol. Evol. 2000, 50, 103–115. [Google Scholar] [CrossRef]
- Weirauch, M.T.; Yang, A.; Albu, M.; Cote, A.G.; Montenegro-Montero, A.; Drewe, P.; Najafabadi, H.S.; Lambert, S.A.; Mann, I.; Cook, K.; et al. Determination and inference of eukaryotic transcription factor sequence specificity. Cell 2014, 158, 1431–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Fukushige, T.; McGhee, J.D.; Rothman, J.H. Reprogramming of early embryonic blastomeres into endodermal progenitors by a Caenorhabditis elegans GATA factor. Genes Dev. 1998, 12, 3809–3814. [Google Scholar] [CrossRef] [Green Version]
- Riddle, M.R.; Weintraub, A.; Nguyen, K.C.; Hall, D.H.; Rothman, J.H. Transdifferentiation and remodeling of post-embryonic C. elegans cells by a single transcription factor. Development 2013, 140, 4844–4849. [Google Scholar] [CrossRef] [Green Version]
- Dineen, A.; Osborne Nishimura, E.; Goszczynski, B.; Rothman, J.H.; McGhee, J.D. Quantitating transcription factor redundancy: The relative roles of the ELT-2 and ELT-7 GATA factors in the C. elegans endoderm. Dev. Biol. 2018, 435, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Gilleard, J.S.; McGhee, J.D. Activation of hypodermal differentiation in the Caenorhabditis elegans embryo by GATA transcription factors ELT-1 and ELT-3. Mol. Cell. Biol. 2001, 21, 2533–2544. [Google Scholar] [CrossRef] [Green Version]
- Broitman-Maduro, G.; Maduro, M.F. The long isoform of the C. elegans ELT-3 GATA factor can specify endoderm when overexpressed. Micropublication Biol. 2023, 2023. [Google Scholar] [CrossRef]
- Broitman-Maduro, G.; Maduro, M.F.; Rothman, J.H. The noncanonical binding site of the MED-1 GATA factor defines differentially regulated target genes in the C. elegans mesendoderm. Dev. Cell 2005, 8, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Lowry, J.A.; Gamsjaeger, R.; Thong, S.Y.; Hung, W.; Kwan, A.H.; Broitman-Maduro, G.; Matthews, J.M.; Maduro, M.; Mackay, J.P. Structural analysis of MED-1 reveals unexpected diversity in the mechanism of DNA recognition by GATA-type zinc finger domains. J. Biol. Chem. 2009, 284, 5827–5835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broitman-Maduro, G.; Owraghi, M.; Hung, W.; Kuntz, S.; Sternberg, P.W.; Maduro, M. The NK-2 class homeodomain factor CEH-51 and the T-box factor TBX-35 have overlapping function in C. elegans mesoderm development. Development 2009, 176, 2735–2746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maduro, M.F.; Lin, R.; Rothman, J.H. Dynamics of a developmental switch: Recursive intracellular and intranuclear redistribution of Caenorhabditis elegans POP-1 parallels Wnt-inhibited transcriptional repression. Dev. Biol. 2002, 248, 128–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGhee, J.D.; Fukushige, T.; Krause, M.W.; Minnema, S.E.; Goszczynski, B.; Gaudet, J.; Kohara, Y.; Bossinger, O.; Zhao, Y.; Khattra, J.; et al. ELT-2 is the predominant transcription factor controlling differentiation and function of the C. elegans intestine, from embryo to adult. Dev. Biol. 2009, 327, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Ewe, C.K.; Sommermann, E.M.; Kenchel, J.; Flowers, S.E.; Maduro, M.F.; Joshi, P.M.; Rothman, J.H. Feedforward regulatory logic controls the specification-to-differentiation transition and terminal cell fate during Caenorhabditis elegans endoderm development. Development 2022, 149, dev200337. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Rifkin, S.A.; Andersen, E.; van Oudenaarden, A. Variability in gene expression underlies incomplete penetrance. Nature 2010, 463, 913–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, L.; Felix, M.A.; Beltran, T.; Braendle, C.; Caurcel, C.; Fausett, S.; Fitch, D.; Frezal, L.; Gosse, C.; Kaur, T.; et al. Comparative genomics of 10 new Caenorhabditis species. Evol. Lett. 2019, 3, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Kiontke, K.C.; Felix, M.A.; Ailion, M.; Rockman, M.V.; Braendle, C.; Penigault, J.B.; Fitch, D.H. A phylogeny and molecular barcodes for Caenorhabditis, with numerous new species from rotting fruits. BMC Evol. Biol. 2011, 11, 339. [Google Scholar] [CrossRef]
- Stevens, L.; Rooke, S.; Falzon, L.C.; Machuka, E.M.; Momanyi, K.; Murungi, M.K.; Njoroge, S.M.; Odinga, C.O.; Ogendo, A.; Ogola, J.; et al. The Genome of Caenorhabditis bovis. Curr. Biol. CB 2020, 30, 1023–1031.e4. [Google Scholar] [CrossRef]
- Stein, L.D.; Bao, Z.; Blasiar, D.; Blumenthal, T.; Brent, M.R.; Chen, N.; Chinwalla, A.; Clarke, L.; Clee, C.; Coghlan, A.; et al. The Genome Sequence of Caenorhabditis briggsae: A Platform for Comparative Genomics. PLoS Biol. 2003, 1, E45. [Google Scholar] [CrossRef]
- Consortium, T.C.e.S. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar]
- Zhao, Z.; Boyle, T.J.; Bao, Z.; Murray, J.I.; Mericle, B.; Waterston, R.H. Comparative analysis of embryonic cell lineage between Caenorhabditis briggsae and Caenorhabditis elegans. Dev. Biol. 2008, 314, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Memar, N.; Schiemann, S.; Hennig, C.; Findeis, D.; Conradt, B.; Schnabel, R. Twenty million years of evolution: The embryogenesis of four Caenorhabditis species are indistinguishable despite extensive genome divergence. Dev. Biol. 2019, 447, 182–199. [Google Scholar] [CrossRef] [PubMed]
- Winston, W.M.; Sutherlin, M.; Wright, A.J.; Feinberg, E.H.; Hunter, C.P. Caenorhabditis elegans SID-2 is required for environmental RNA interference. Proc. Natl. Acad. Sci. USA 2007, 104, 10565–10570. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.P.; Johnsen, R.; Chen, N. Genomics and biology of the nematode Caenorhabditis briggsae. In WormBook: The Online Review of C. elegans Biology; 2007; pp. 1–16. Available online: https://pubmed.ncbi.nlm.nih.gov/18050493/ (accessed on 1 June 2023). [CrossRef]
- Slos, D.; Sudhaus, W.; Stevens, L.; Bert, W.; Blaxter, M. Caenorhabditis monodelphis sp. n.: Defining the stem morphology and genomics of the genus Caenorhabditis. BMC Zool. 2017, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Flibotte, S.; Murray, J.I.; Blick, D.; Boyle, T.J.; Gupta, B.; Moerman, D.G.; Waterston, R.H. New tools for investigating the comparative biology of Caenorhabditis briggsae and C. elegans. Genetics 2010, 184, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Maduro, M.F.; Broitman-Maduro, G.; Choi, H.; Carranza, F.; Chia-Yi Wu, A.; Rifkin, S.A. MED GATA factors promote robust development of the C. elegans endoderm. Dev. Biol. 2015, 404, 66–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesenfahrt, T.; Osborne Nishimura, E.; Berg, J.Y.; McGhee, J.D. Probing and rearranging the transcription factor network controlling the C. elegans endoderm. Worm 2016, 5, e1198869. [Google Scholar] [CrossRef]
- Koh, K.; Rothman, J.H. ELT-5 and ELT-6 are required continuously to regulate epidermal seam cell differentiation and cell fusion in C. elegans. Development 2001, 128, 2867–2880. [Google Scholar] [CrossRef]
- Page, B.D.; Zhang, W.; Steward, K.; Blumenthal, T.; Priess, J.R. ELT-1, a GATA-like transcription factor, is required for epidermal cell fates in Caenorhabditis elegans embryos. Genes Dev. 1997, 11, 1651–1661. [Google Scholar] [CrossRef] [Green Version]
- Tsanov, N.; Samacoits, A.; Chouaib, R.; Traboulsi, A.M.; Gostan, T.; Weber, C.; Zimmer, C.; Zibara, K.; Walter, T.; Peter, M.; et al. smiFISH and FISH-quant—A flexible single RNA detection approach with super-resolution capability. Nucleic Acids Res. 2016, 44, e165. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; He, K.; Wang, H.; Ho, W.S.; Ren, X.; An, X.; Wong, M.K.; Yan, B.; Xie, D.; Stamatoyannopoulos, J.; et al. Collaborative regulation of development but independent control of metabolism by two epidermis-specific transcription factors in Caenorhabditis elegans. J. Biol. Chem. 2013, 288, 33411–33426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; D’Amora, D.R.; MacNeil, L.T.; Walhout, A.J.M.; Kubiseski, T.J. The Oxidative Stress Response in Caenorhabditis elegans Requires the GATA Transcription Factor ELT-3 and SKN-1/Nrf2. Genetics 2017, 206, 1909–1922. [Google Scholar] [CrossRef] [Green Version]
- Mesbahi, H.; Pho, K.B.; Tench, A.J.; Leon Guerrero, V.L.; MacNeil, L.T. Cuticle Collagen Expression Is Regulated in Response to Environmental Stimuli by the GATA Transcription Factor ELT-3 in Caenorhabditis elegans. Genetics 2020, 215, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Tabarraei, H.; Waddell, B.M.; Raymond, K.; Murray, S.M.; Wang, Y.; Choe, K.P.; Wu, C.W. CCR4-NOT subunit CCF-1/CNOT7 promotes transcriptional activation to multiple stress responses in Caenorhabditis elegans. Aging Cell 2023, 22, e13795. [Google Scholar] [CrossRef]
- Li, R.; Ren, X.; Ding, Q.; Bi, Y.; Xie, D.; Zhao, Z. Direct full-length RNA sequencing reveals unexpected transcriptome complexity during Caenorhabditis elegans development. Genome Res. 2020, 30, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.; Hirsh, D. A trans-spliced leader sequence on actin mRNA in C. elegans. Cell 1987, 49, 753–761. [Google Scholar] [CrossRef]
- Hu, G.; Katuwawala, A.; Wang, K.; Wu, Z.; Ghadermarzi, S.; Gao, J.; Kurgan, L. flDPnn: Accurate intrinsic disorder prediction with putative propensities of disorder functions. Nat. Commun. 2021, 12, 4438. [Google Scholar] [CrossRef]
- Romano, O.; Miccio, A. GATA factor transcriptional activity: Insights from genome-wide binding profiles. IUBMB Life 2020, 72, 10–26. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, L.; Dai, T.; Qin, Z.; Lu, H.; Zhou, F. Liquid-liquid phase separation in human health and diseases. Signal Transduct. Target. Ther. 2021, 6, 290. [Google Scholar] [CrossRef]
- Hnisz, D.; Shrinivas, K.; Young, R.A.; Chakraborty, A.K.; Sharp, P.A. A Phase Separation Model for Transcriptional Control. Cell 2017, 169, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, I.; Han, S.K.; Bowie, J.U.; Kim, S. Network rewiring is an important mechanism of gene essentiality change. Sci. Rep. 2012, 2, 900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, E.S.; True, J.R. Developmental System Drift. In Evolutionary Developmental Biology: A Reference Guide; Nuño de la Rosa, L., Müller, G.B., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 99–110. [Google Scholar] [CrossRef]
- Wilkins, A.S. Moving up the hierarchy: A hypothesis on the evolution of a genetic sex determination pathway. BioEssays News Rev. Mol. Cell. Dev. Biol. 1995, 17, 71–77. [Google Scholar] [CrossRef]
- Lin, R.; Hill, R.J.; Priess, J.R. POP-1 and anterior-posterior fate decisions in C. elegans embryos. Cell 1998, 92, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couthier, A.; Smith, J.; McGarr, P.; Craig, B.; Gilleard, J.S. Ectopic expression of a Haemonchus contortus GATA transcription factor in Caenorhabditis elegans reveals conserved function in spite of extensive sequence divergence. Mol. Biochem. Parasitol. 2004, 133, 241–253. [Google Scholar] [CrossRef]
- Stevens, L.; Moya, N.D.; Tanny, R.E.; Gibson, S.B.; Tracey, A.; Na, H.; Chitrakar, R.; Dekker, J.; Walhout, A.J.M.; Baugh, L.R.; et al. Chromosome-Level Reference Genomes for Two Strains of Caenorhabditis briggsae: An Improved Platform for Comparative Genomics. Genome Biol. Evol. 2022, 14, evac042. [Google Scholar] [CrossRef]
- Sommer, R.J. As good as they get: Cells in nematode vulva development and evolution. Curr. Opin. Cell. Biol. 2001, 13, 715–720. [Google Scholar] [CrossRef]
- Barkoulas, M.; Vargas Velazquez, A.M.; Peluffo, A.E.; Felix, M.A. Evolution of New cis-Regulatory Motifs Required for Cell-Specific Gene Expression in Caenorhabditis. PLoS Genet. 2016, 12, e1006278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, P.W. Vulval development. In WormBook: The Online Review of C. elegans Biology; 2005; pp. 1–28. Available online: https://pubmed.ncbi.nlm.nih.gov/18050418/ (accessed on 1 June 2023). [CrossRef] [Green Version]
- Felix, M.A. Cryptic quantitative evolution of the vulva intercellular signaling network in Caenorhabditis. Curr. Biol. CB 2007, 17, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bei, Y.; Hogan, J.; Berkowitz, L.A.; Soto, M.; Rocheleau, C.E.; Pang, K.M.; Collins, J.; Mello, C.C. SRC-1 and Wnt signaling act together to specify endoderm and to control cleavage orientation in early C. elegans embryos. Dev. Cell 2002, 3, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, C.J.; Schlesinger, A.; Bowerman, B. Wnt signalling in Caenorhabditis elegans: Regulating repressors and polarizing the cytoskeleton. Trends Cell Biol. 2000, 10, 10–17. [Google Scholar] [CrossRef]
- Davis, G.K.; Patel, N.H. Short, long, and beyond: Molecular and embryological approaches to insect segmentation. Annu. Rev. Entomol. 2002, 47, 669–699. [Google Scholar] [CrossRef] [Green Version]
- Lemke, S.; Stauber, M.; Shaw, P.J.; Rafiqi, A.M.; Prell, A.; Schmidt-Ott, U. Bicoid occurrence and Bicoid-dependent hunchback regulation in lower cyclorrhaphan flies. Evol. Dev. 2008, 10, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Stauber, M.; Prell, A.; Schmidt-Ott, U. A single Hox3 gene with composite bicoid and zerknullt expression characteristics in non-Cyclorrhaphan flies. Proc. Natl. Acad. Sci. USA 2002, 99, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.A.; Brent, A.E.; Leaf, D.S.; Pultz, M.A.; Desplan, C. Localized maternal orthodenticle patterns anterior and posterior in the long germ wasp Nasonia. Nature 2006, 439, 728–732. [Google Scholar] [CrossRef]
- McGregor, A.P. How to get ahead: The origin, evolution and function of bicoid. BioEssays News Rev. Mol. Cell. Dev. Biol. 2005, 27, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Schroder, R. The genes orthodenticle and hunchback substitute for bicoid in the beetle Tribolium. Nature 2003, 422, 621–625. [Google Scholar] [CrossRef]
- Kotkamp, K.; Klingler, M.; Schoppmeier, M. Apparent role of Tribolium orthodenticle in anteroposterior blastoderm patterning largely reflects novel functions in dorsoventral axis formation and cell survival. Development 2010, 137, 1853–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sherif, E.; Lynch, J.A.; Brown, S.J. Comparisons of the embryonic development of Drosophila, Nasonia, and Tribolium. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 16–39. [Google Scholar] [CrossRef] [Green Version]
- Rudolf, H.; Zellner, C.; El-Sherif, E. Speeding up anterior-posterior patterning of insects by differential initialization of the gap gene cascade. Dev. Biol. 2020, 460, 20–31. [Google Scholar] [CrossRef]
- Brauchle, M.; Kiontke, K.; MacMenamin, P.; Fitch, D.H.; Piano, F. Evolution of early embryogenesis in rhabditid nematodes. Dev. Biol. 2009, 335, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Lowe, E.K.; Stolfi, A. Developmental system drift in motor ganglion patterning between distantly related tunicates. EvoDevo 2018, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Stolfi, A.; Lowe, E.K.; Racioppi, C.; Ristoratore, F.; Brown, C.T.; Swalla, B.J.; Christiaen, L. Divergent mechanisms regulate conserved cardiopharyngeal development and gene expression in distantly related ascidians. eLife 2014, 3, e03728. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Levine, M. Neuronal subtype specification in the spinal cord of a protovertebrate. Development 2011, 138, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packer, J.S.; Zhu, Q.; Huynh, C.; Sivaramakrishnan, P.; Preston, E.; Dueck, H.; Stefanik, D.; Tan, K.; Trapnell, C.; Kim, J.; et al. A lineage-resolved molecular atlas of C. elegans embryogenesis at single-cell resolution. Science 2019, 365, eaax1971. [Google Scholar] [CrossRef] [PubMed]
- Nuez, I.; Felix, M.A. Evolution of susceptibility to ingested double-stranded RNAs in Caenorhabditis nematodes. PLoS ONE 2012, 7, e29811. [Google Scholar] [CrossRef] [Green Version]
- Tonsaker, T.; Pratt, R.M.; McGhee, J.D. Re-evaluating the role of ELT-3 in a GATA transcription factor circuit proposed to guide aging in C. elegans. Mech. Ageing Dev. 2012, 133, 50–53. [Google Scholar] [CrossRef]
- Budovskaya, Y.V.; Wu, K.; Southworth, L.K.; Jiang, M.; Tedesco, P.; Johnson, T.E.; Kim, S.K. An elt-3/elt-5/elt-6 GATA transcription circuit guides aging in C. elegans. Cell 2008, 134, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.L.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Schierenberg, E. Three sons of fortune: Early embryogenesis, evolution and ecology of nematodes. BioEssays News Rev. Mol. Cell. Dev. Biol. 2001, 23, 841–847. [Google Scholar] [CrossRef]
- Macchietto, M.; Angdembey, D.; Heidarpour, N.; Serra, L.; Rodriguez, B.; El-Ali, N.; Mortazavi, A. Comparative Transcriptomics of Steinernema and Caenorhabditis Single Embryos Reveals Orthologous Gene Expression Convergence during Late Embryogenesis. Genome Biol. Evol. 2017, 9, 2681–2696. [Google Scholar] [CrossRef]
- Johnson, N.A.; Porter, A.H. Evolution of branched regulatory genetic pathways: Directional selection on pleiotropic loci accelerates developmental system drift. Genetica 2007, 129, 57–70. [Google Scholar] [CrossRef]
- Pavlicev, M.; Wagner, G.P. A model of developmental evolution: Selection, pleiotropy and compensation. Trends Ecol. Evol. 2012, 27, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Nance, J.; Lee, J.Y.; Goldstein, B. Gastrulation in C. elegans. In WormBook: The Online Review of C. elegans Biology; 2005; pp. 1–13. Available online: https://pubmed.ncbi.nlm.nih.gov/18050409/ (accessed on 1 June 2023). [CrossRef] [Green Version]
- Robertson, S.M.; Medina, J.; Lin, R. Uncoupling different characteristics of the C. elegans E lineage from differentiation of intestinal markers. PLoS ONE 2014, 9, e106309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeck, M.E.; Boyle, T.; Bao, Z.; Murray, J.; Mericle, B.; Waterston, R. Specific roles for the GATA transcription factors end-1 and end-3 during C. elegans E-lineage development. Dev. Biol. 2011, 358, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Clucas, C.; Cabello, J.; Bussing, I.; Schnabel, R.; Johnstone, I.L. Oncogenic potential of a C. elegans cdc25 gene is demonstrated by a gain-of-function allele. Embo. J. 2002, 21, 665–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, I.; Roy, R. Organ-specific cell division abnormalities caused by mutation in a general cell cycle regulator in C. elegans. Development 2002, 129, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Hashimshony, T.; Wagner, F.; Sher, N.; Yanai, I. CEL-Seq: Single-cell RNA-Seq by multiplexed linear amplification. Cell Rep. 2012, 2, 666–673. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Loss-of-Function Phenotype in C. elegans | Loss-of-Function Phenotype in C. briggsae * | Loss-of-Function Phenotype in C. angaria * |
|---|---|---|---|
| Maternal factors | |||
| skn-1/Nrf | Loss of MS, E fates [27] | Loss of MS, E fates [12] | No phenotype [9] |
| pop-1/TCF | Excess gut from MS [28] | Loss of gut [12] | Loss of gut [9] |
| Zygotic GATA factors | |||
| med-1, med-2 | Loss of MS, E fates [47] | n.d. | n/a |
| end-1, end-3 | Loss of gut [13] | Loss of gut [13] | n/a |
| elt-3 | No phenotype [48] | n.d. | Loss of gut [9] |
| elt-7 | No phenotype; enhances elt-2(-) [49] | n.d. | n/a |
| elt-2 | Incomplete gut differentiation [50] | n.d. | Incomplete gut differentiation [9] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broitman-Maduro, G.; Maduro, M.F. Evolutionary Change in Gut Specification in Caenorhabditis Centers on the GATA Factor ELT-3 in an Example of Developmental System Drift. J. Dev. Biol. 2023, 11, 32. https://doi.org/10.3390/jdb11030032
Broitman-Maduro G, Maduro MF. Evolutionary Change in Gut Specification in Caenorhabditis Centers on the GATA Factor ELT-3 in an Example of Developmental System Drift. Journal of Developmental Biology. 2023; 11(3):32. https://doi.org/10.3390/jdb11030032
Chicago/Turabian StyleBroitman-Maduro, Gina, and Morris F. Maduro. 2023. "Evolutionary Change in Gut Specification in Caenorhabditis Centers on the GATA Factor ELT-3 in an Example of Developmental System Drift" Journal of Developmental Biology 11, no. 3: 32. https://doi.org/10.3390/jdb11030032