
Patterning of the Vertebrate Head in Time and Space by BMP Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microinjection
2.2. Whole Mount In Situ Hybridization
3. Results
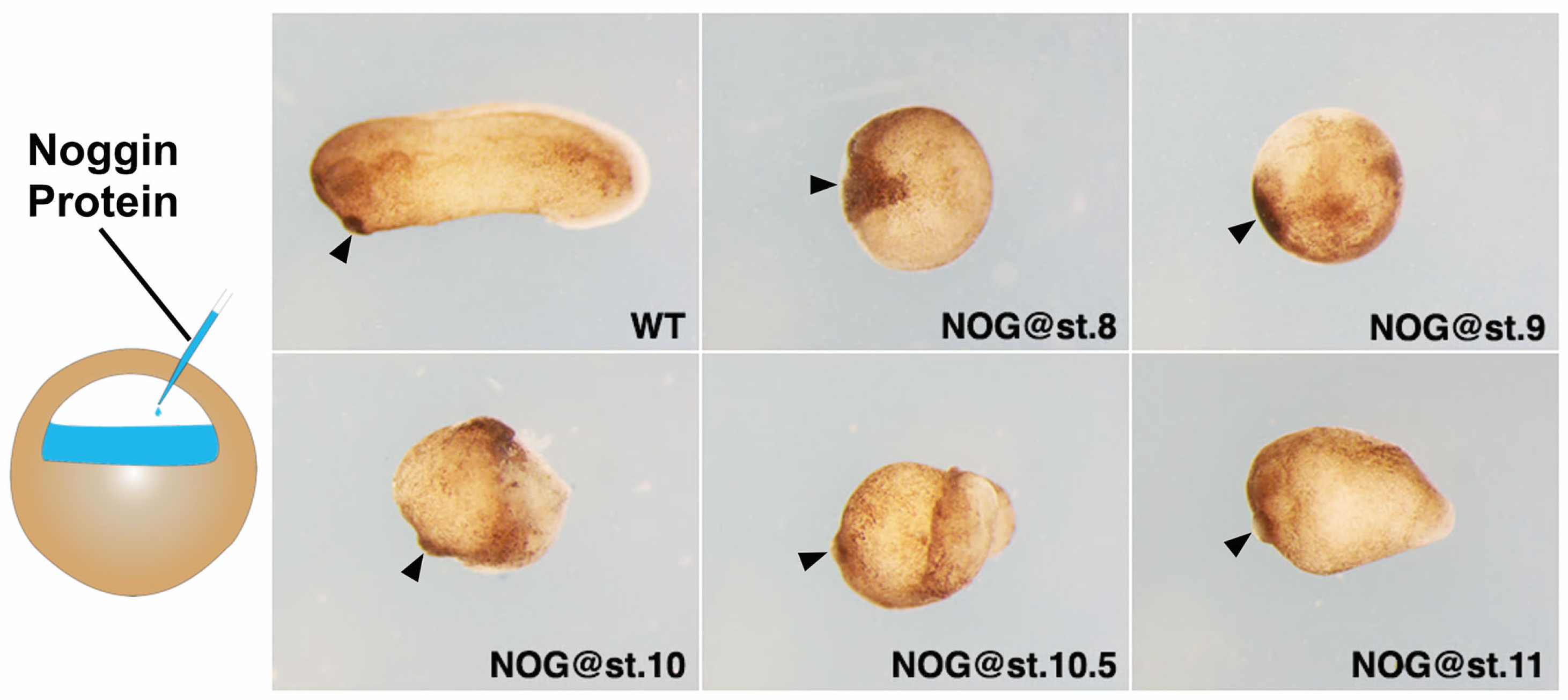
3.1. Timed Anti-BMP Treatment Arrests Head Patterning at Different Positions
3.2. Timed Anti-BMP Treatment in Ventralized Embryos Rescued Different Portions of the Head
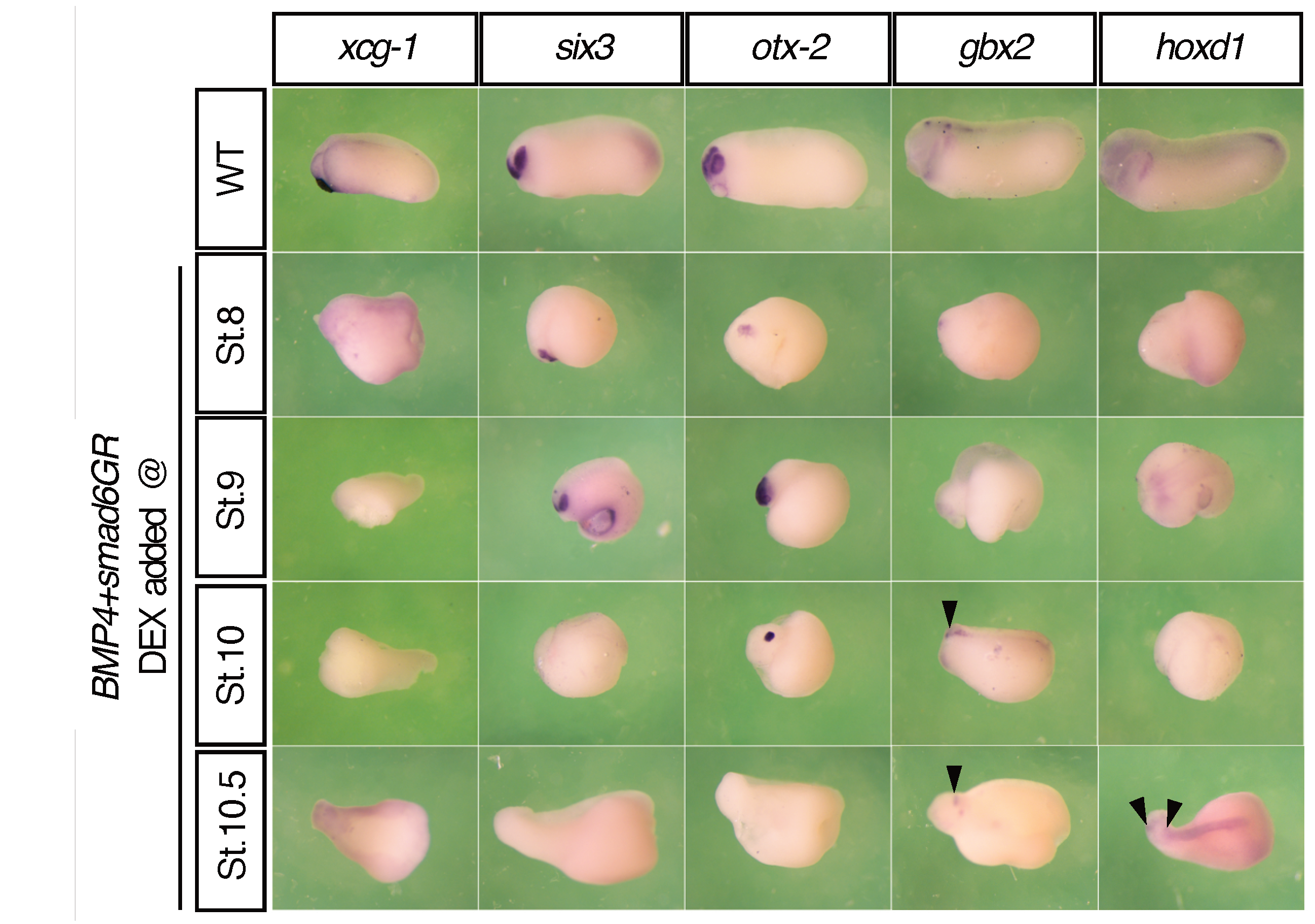
3.3. The Timing of A–P Markers Is Disrupted in smad6-Injected Embryos
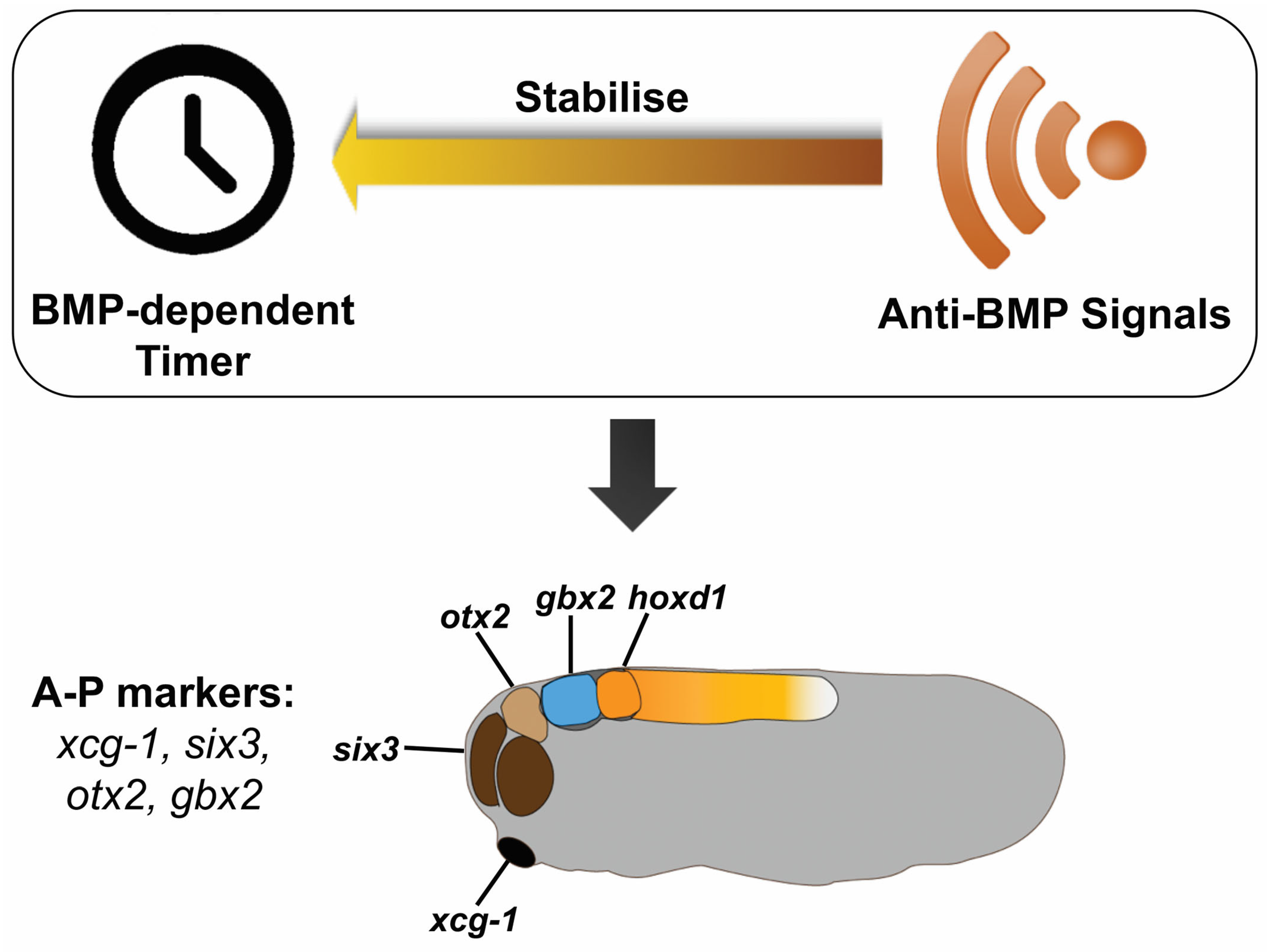
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eyal-Giladi, H. Dynamic Aspects of Neural Induction in Amphibia. Arch. Biol. 1954, 65, 179–259. [Google Scholar]
- Gamse, J.; Sive, H. Vertebrate Anteroposterior Patterning: The Xenopus Neurectoderm as a Paradigm. Bioessays 2000, 22, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Gamse, J.T.; Sive, H. Early Anteroposterior Division of the Presumptive Neurectoderm in Xenopus. Mech. Dev. 2001, 104, 21–36. [Google Scholar] [CrossRef]
- Nieuwkoop, P.D. Activation and Organization of the Central Nervous System in Amphibians. Part III. Synthesis of a New Working Hypothesis. J. Exp. Zool. 1952, 120, 83–108. [Google Scholar] [CrossRef]
- Stern, C.D.; Charite, J.; Deschamps, J.; Duboule, D.; Durston, A.J.; Kmita, M.; Nicolas, J.F.; Palmeirim, I.; Smith, J.C.; Wolpert, L. Head-Tail Patterning of the Vertebrate Embryo: One, Two or Many Unresolved Problems? Int. J. Dev. Biol. 2006, 50, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Durston, A.J.; Zhu, K. A Time Space Translation Hypothesis for Vertebrate Axial Patterning. Semin. Cell Dev. Biol. 2015, 42, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Wacker, S.A.; Jansen, H.J.; McNulty, C.L.; Houtzager, E.; Durston, A.J. Timed Interactions between the Hox Expressing Non-Organiser Mesoderm and the Spemann Organiser Generate Positional Information During Vertebrate Gastrulation. Dev. Biol. 2004, 268, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacker, S.; McNulty, C.; Durston, A. The Initiation of Hox Gene Expression in Xenopus Laevis Is Controlled by Brachyury and Bmp-4. Dev. Biol. 2004, 266, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.C.; Harland, R.M. Expression Cloning of Noggin, a New Dorsalizing Factor Localized to the Spemann Organizer in Xenopus Embryos. Cell 1992, 70, 829–840. [Google Scholar] [CrossRef]
- Sasai, Y.; Lu, B.; Steinbeisser, H.; Geissert, D.; Gont, L.K.; De Robertis, E. Xenopus Chordin: A Novel Dorsalizing Factor. Activated by Organizer-Specific Homeobox Genes. Cell 1994, 79, 779–790. [Google Scholar]
- Smith, W.C.; Knecht, A.K.; Wu, M.; Harland, R.M. Secreted Noggin Protein Mimics the Spemann Organizer in Dorsalizing Xenopus Mesoderm. Nature 1993, 361, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Khokha, M.K.; Yeh, J.; Grammer, T.C.; Harland, R.M. Depletion of Three Bmp Antagonists from Spemann’s Organizer Leads to a Catastrophic Loss of Dorsal Structures. Dev. Cell 2005, 8, 401–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Marikawa, Y.; Elinson, R.P. Ectopic Expression of Xenopus Noggin Rna Induces Complete Secondary Body Axes in Embryos of the Direct Developing Frog Eleutherodactylus Coqui. Dev. Genes Evol. 2000, 210, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Spemann, H.; Mangold, H. Über Induktion Von Embryonalagen Durch Implantation Artfremder Organisatoren. Roux’s Arch. F. Entw. Mech. 1924, 100, 599–638. [Google Scholar]
- Dias, A.S.; de Almeida, I.; Belmonte, J.M.; Glazier, J.A.; Stern, C.D. Somites without a Clock. Science 2014, 343, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Toyama, R.; Takeda, H.; Dawid, I.B.; Kawakami, K. Overexpression of the Forebrain-Specific Homeobox Gene Six3 Induces Rostral Forebrain Enlargement in Zebrafish. Development 1998, 125, 2973–2982. [Google Scholar] [CrossRef]
- Li, Y.; Allende, M.L.; Finkelstein, R.; Weinberg, E.S. Expression of Two Zebrafish Orthodenticle-Related Genes in the Embryonic Brain. Mech. Dev. 1994, 48, 229–244. [Google Scholar] [CrossRef]
- Mori, H.; Miyazaki, Y.; Morita, T.; Nitta, H.; Mishina, M. Different Spatio-Temporal Expressions of Three Otx Homeoprotein Transcripts During Zebrafish Embryogenesis. Brain Res. Mol. Brain Res. 1994, 27, 221–231. [Google Scholar] [CrossRef]
- Rhinn, M.; Lun, K.; Amores, A.; Yan, Y.L.; Postlethwait, J.H.; Brand, M. Cloning, Expression and Relationship of Zebrafish Gbx1 and Gbx2 Genes to Fgf Signaling. Mech. Dev. 2003, 120, 919–936. [Google Scholar] [CrossRef]
- Alexandre, D.; Clarke, J.D.; Oxtoby, E.; Yan, Y.L.; Jowett, T.; Holder, N. Ectopic Expression of Hoxa-1 in the Zebrafish Alters the Fate of the Mandibular Arch Neural Crest and Phenocopies a Retinoic Acid-Induced Phenotype. Development 1996, 122, 735–746. [Google Scholar] [CrossRef]
- Hashiguchi, M.; Mullins, M.C. Anteroposterior and Dorsoventral Patterning Are Coordinated by an Identical Patterning Clock. Development 2013, 140, 1970–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, J.A.; Mintzer, K.A.; Mullins, M.C. The Bmp Signaling Gradient Patterns Dorsoventral Tissues in a Temporally Progressive Manner Along the Anteroposterior Axis. Dev. Cell 2008, 14, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacox, L.; Sindelka, R.; Chen, J.; Rothman, A.; Dickinson, A.; Sive, H. The Extreme Anterior Domain Is an Essential Craniofacial Organizer Acting through Kinin-Kallikrein Signaling. Cell Rep. 2014, 8, 596–609. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, A.; Sive, H. Positioning the Extreme Anterior in Xenopus: Cement Gland, Primary Mouth and Anterior Pituitary. Semin. Cell Dev. Biol. 2007, 18, 525–533. [Google Scholar]
- Sive, H.; Hattori, K.; Weintraub, H. Progressive Determination During Formation of the Anteroposterior Axis in Xenopus Laevis. Cell 1989, 58, 171–180. [Google Scholar] [CrossRef]
- Scharf, S.R.; Gerhart, J.C. Axis Determination in Eggs of Xenopus Laevis: A Critical Period before First Cleavage, Identified by the Common Effects of Cold, Pressure and Ultraviolet Irradiation. Dev. Biol. 1983, 99, 75–87. [Google Scholar] [CrossRef]
- Kao, K.R.; Elinson, R.P. The Entire Mesodermal Mantle Behaves as Spemann’s Organizer in Dorsoanterior Enhanced Xenopus Laevis Embryos. Dev. Biol. 1988, 127, 64–77. [Google Scholar] [CrossRef]
- Faber, J.; Nieuwkoop, P.D. Normal Table of Xenopus Laevis (Daudin): A Systematical and Chronological Survey of the Development from the Fertilized Egg Till the End of Metamorphosis; Garland Pub: New York, NY, USA, 1994. [Google Scholar]
- Cooke, J.; Smith, J. Gastrulation and Larval Pattern in Xenopus after Blastocoelic Injection of a Xenopus-Derived Inducing Factor: Experiments Testing Models for the Normal Organization of Mesoderm. Dev. Biol. 1989, 131, 383–400. [Google Scholar] [CrossRef]
- Nishimatsu, S.I.; Suzuki, A.; Shoda, A.; Murakami, K.; Ueno, N. Genes for Bone Morphogenetic Proteins Are Differentially Transcribed in Early Amphibian Embryos. Biochem. Biophys. Res. Commun. 1992, 186, 1487–1495. [Google Scholar] [CrossRef]
- Marom, K.; Levy, V.; Pillemer, G.; Fainsod, A. Temporal Analysis of the Early Bmp Functions Identifies Distinct Anti-Organizer and Mesoderm Patterning Phases. Dev. Biol. 2005, 282, 442–454. [Google Scholar] [CrossRef] [Green Version]
- Gammill, L.; Sive, H. Coincidence of Otx2 and Bmp4 Signaling Correlates with Xenopus Cement Gland Formation. Mech. Dev. 2000, 92, 217–226. [Google Scholar] [CrossRef]
- Kenyon, K.L.; Moody, S.A.; Jamrich, M. A Novel Fork Head Gene Mediates Early Steps During Xenopus Lens Formation. Development 1999, 126, 5107–5116. [Google Scholar] [CrossRef]
- Blitz, I.L.; Cho, K.W. Anterior Neurectoderm Is Progressively Induced During Gastrulation—The Role of the Xenopus Homeobox Gene Orthodenticle. Development 1995, 121, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Von Bubnoff, A.; Schmidt, J.E.; Kimelman, D. The Xenopus Laevis Homeobox Gene Xgbx-2 Is an Early Marker of Anteroposterior Patterning in the Ectoderm. Mech. Dev. 1996, 54, 149–160. [Google Scholar] [CrossRef]
- Sive, H.L.; Cheng, P.F. Retinoic Acid Perturbs the Expression of Xhox.Lab Genes and Alters Mesodermal Determination in Xenopus Laevis. Genes Dev. 1991, 5, 1321–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, L.B.; De Jesús-Escobar, J.M.; Harland, R.M. The Spemann Organizer Signal Noggin Binds and Inactivates Bone Morphogenetic Protein 4. Cell 1996, 86, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Clement, J.H.; Fettes, P.; Knöchel, S.; Lef, J.; Knöchel, W. Bone Morphogenetic Protein-2 in the Early Development of Xenopus-Laevis. Mech. Dev. 1995, 3, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Dale, L.; Howes, G.; Price, B.M.; Smith, J.C. Bone Morphogenetic Protein 4: A Ventralizing Factor in Early Xenopus Development. Development 1992, 115, 573–585. [Google Scholar] [CrossRef]
- Jones, C.M.; Lyons, K.M.; Lapan, P.M.; Wright, C.V.; Hogan, B.L. Dvr-4 (Bone Morphogenetic Protein-4) as a Posterior-Ventralizing Factor in Xenopus Mesoderm Induction. Development 1992, 115, 639–647. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Suzuki, A.; Ueno, N.; Kimelman, D. Localized Bmp-4 Mediates Dorsal/Ventral Patterning in the Early Xenopus Embryo. Dev. Biol. 1995, 169, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Spaink, H.P.; Durston, A.J. Collinear Hox-Hox Interactions Are Involved in Patterning the Vertebrate Anteroposterior (a-P) Axis. PLoS ONE 2017, 12, e0175287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, K.; Kamiya, Y.; Imamura, T.; Miyazono, K.; Miyazawa, K. Selective Inhibitory Effects of Smad6 on Bone Morphogenetic Protein Type I Receptors. J. Biol. Chem. 2007, 282, 20603–20611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hata, A.; Lagna, G.; Massagué, J.; Hemmati-Brivanlou, A. Smad6 Inhibits Bmp/Smad1 Signaling by Specifically Competing with the Smad4 Tumor Suppressor. Genes Dev. 1998, 12, 186–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, T.; Takase, M.; Nishihara, A.; Oeda, E.; Hanai, J.; Kawabata, M.; Miyazono, K. Smad6 Inhibits Signalling by the Tgf-Beta Superfamily. Nature 1997, 389, 622–626. [Google Scholar] [CrossRef]
- Duboule, D.; Dollé, P. The Structural and Functional-Organization of the Murine Hox Gene Family Resembles That of Drosophila Homeotic Genes. EMBO J. 1989, 8, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Graham, A.; Papalopulu, N.; Krumlauf, R. The Murine and Drosophila Homeobox Gene Complexes Have Common Features of Organization and Expression. Cell 1989, 57, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.B. A Gene Complex Controlling Segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Mailhos, A.; Wehr, R.; Copeland, N.G.; Jenkins, N.A.; Gruss, P. Six3, a Murine Homologue of the Sine Oculis Gene, Demarcates the Most Anterior Border of the Developing Neural Plate and Is Expressed During Eye Development. Development 1995, 121, 4045–4055. [Google Scholar] [CrossRef]
- Kessel, M.; Gruss, P. Homeotic Transformations of Murine Vertebrae and Concomitant Alteration of Hox Codes Induced by Retinoic Acid. Cell 1991, 67, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Wellik, D.M.; Capecchi, M.R. Hox10 and Hox11 Genes Are Required to Globally Pattern the Mammalian Skeleton. Science 2003, 301, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Tour, E.; Pillemer, G.; Gruenbaum, Y.; Fainsod, A. Gbx2 Interacts with Otx2 and Patterns the Anterior-Posterior Axis During Gastrulation in Xenopus. Mech. Dev. 2002, 2, 141–151. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.; Spaink, H.P.; Durston, A.J. Patterning of the Vertebrate Head in Time and Space by BMP Signaling. J. Dev. Biol. 2023, 11, 31. https://doi.org/10.3390/jdb11030031
Zhu K, Spaink HP, Durston AJ. Patterning of the Vertebrate Head in Time and Space by BMP Signaling. Journal of Developmental Biology. 2023; 11(3):31. https://doi.org/10.3390/jdb11030031
Chicago/Turabian StyleZhu, Kongju, Herman P. Spaink, and Antony J. Durston. 2023. "Patterning of the Vertebrate Head in Time and Space by BMP Signaling" Journal of Developmental Biology 11, no. 3: 31. https://doi.org/10.3390/jdb11030031