

Regionalized Protein Localization Domains in the Zebrafish Hair Cell Kinocilium
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Husbandry
2.2. Transgenic Fish Lines
2.3. Thiouracil (TU)-Tagging and Bioinformatics
2.4. Whole Mount mRNA In Situ Hybridization
2.5. Confocal Microscopy, Image Analysis, and Statistics
3. Results
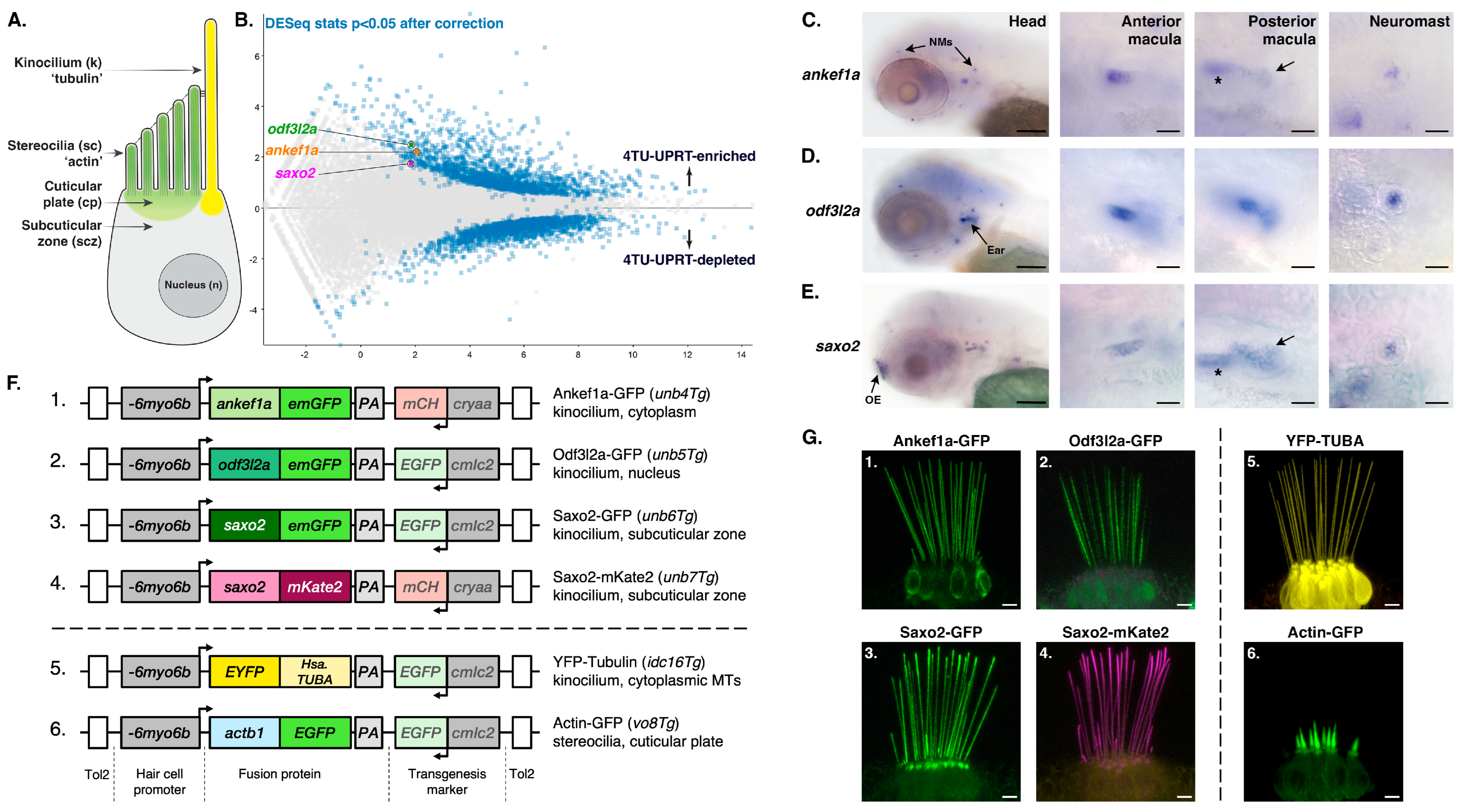
3.1. Identification of Putative Kinocilia-Associated, Hair Cell-Enriched Transcripts
3.2. Evaluation of ANKEF1, ODF3L2, and SAXO2 as Potential Human Deafness Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Gene | Cytogenetic Location | GRCh38.p13 Coordinates | Proposed Deafness Locus | Critical Region Markers | Locus Coordinates | Causative Gene | Reference |
|---|---|---|---|---|---|---|---|
| ANKEF1 | 20p12.2 | 20:10034987-10058303 | n/a | n/a | n/a | n/a | n/a |
| ODF3L2 | 19p13.3 | 19:463346-474880 | DFNB72 | D19S216–D19S1034 | 19:4949401-6113471 | GIPC3 | Ain et al. [76] |
| DFNA57 | D19S912–D19S212 | 19:7806762-18232460 | unknown | Bönsch et al. [75] | |||
| SAXO2 | 15q25.2 | 15:82262818-82284927 | DFNA30 | D15S151–D15S130 | 15:87053920-94168232 | unknown | Mangino et al. [77] |
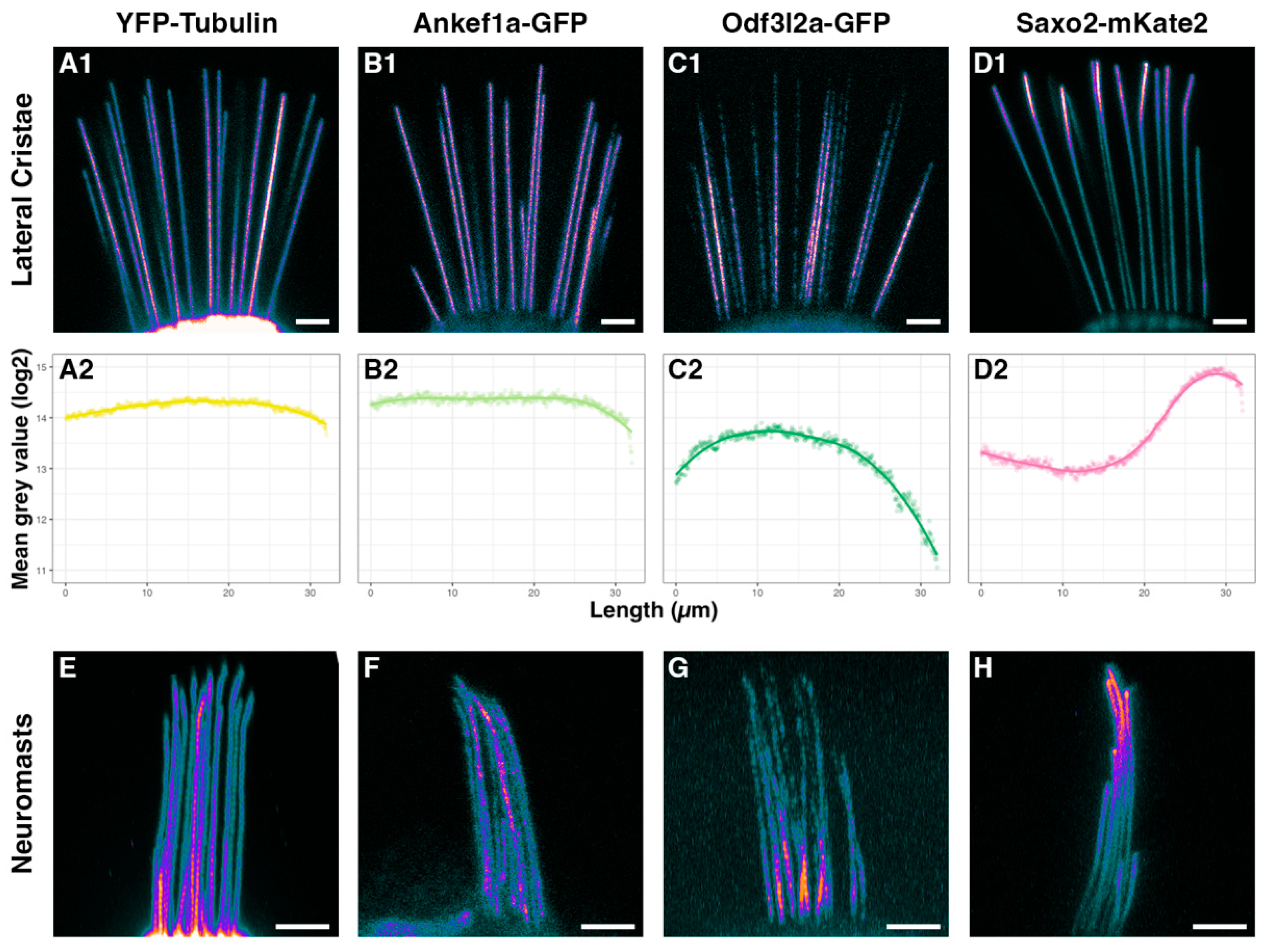
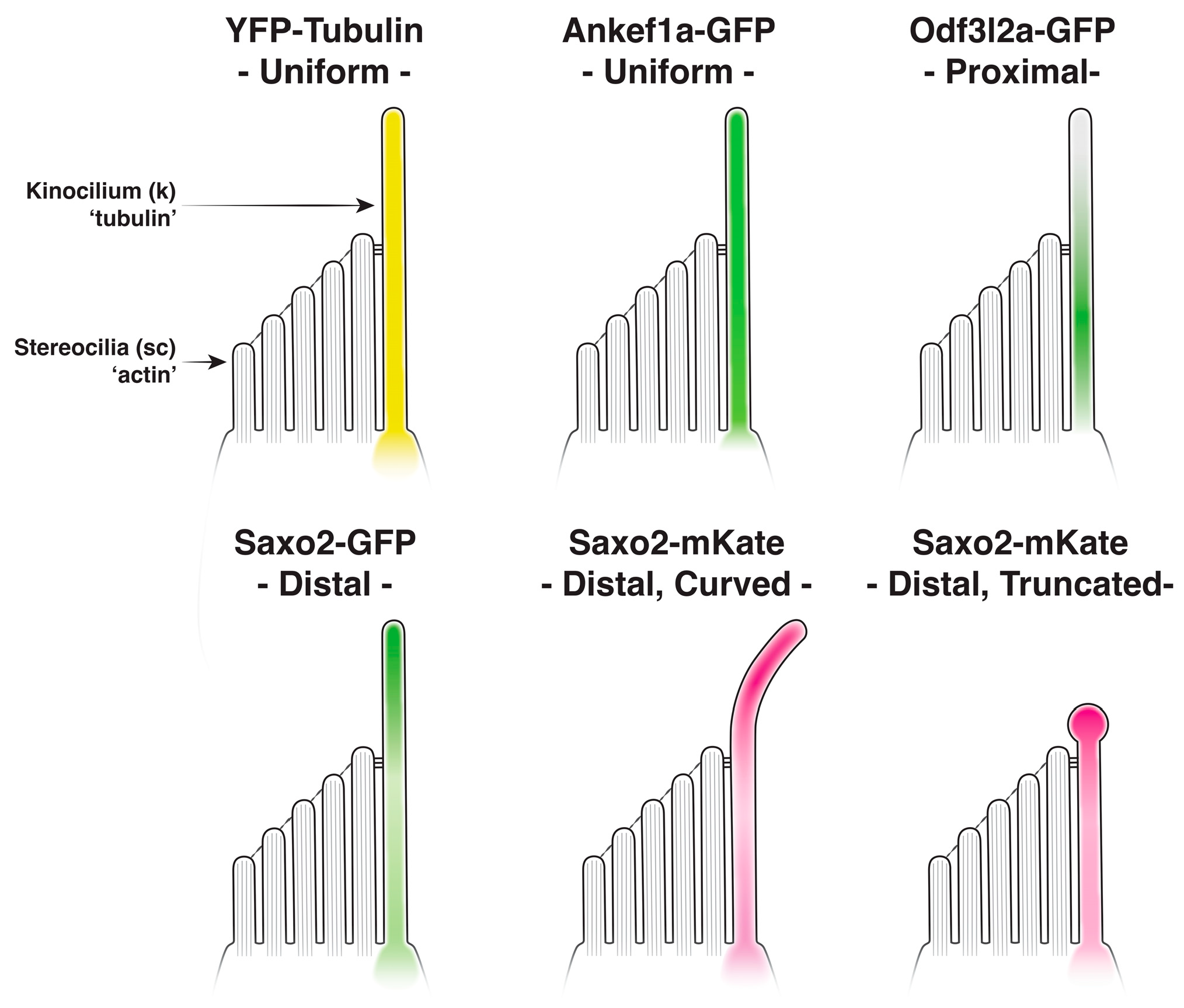
3.3. Distinct Localization Patterns of Ankef1a, Odf3l2a, and Saxo2 in the Kinocilium
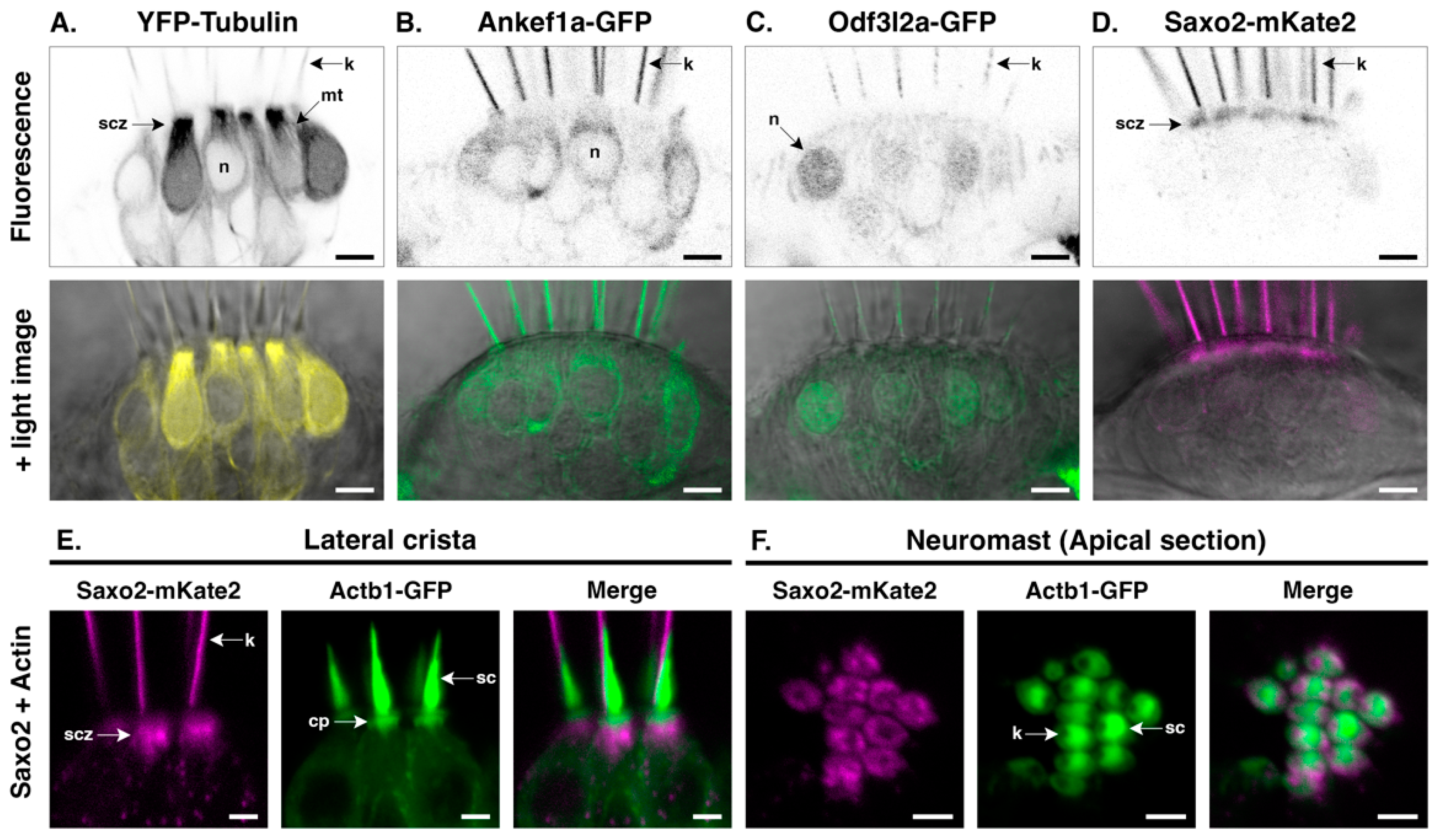
3.4. Ankef1a, Odf3l2a, and Saxo2 Exhibit Distinct Localization Patterns in the Hair Cell Soma That Differ from Cytoplasmic Tubulin
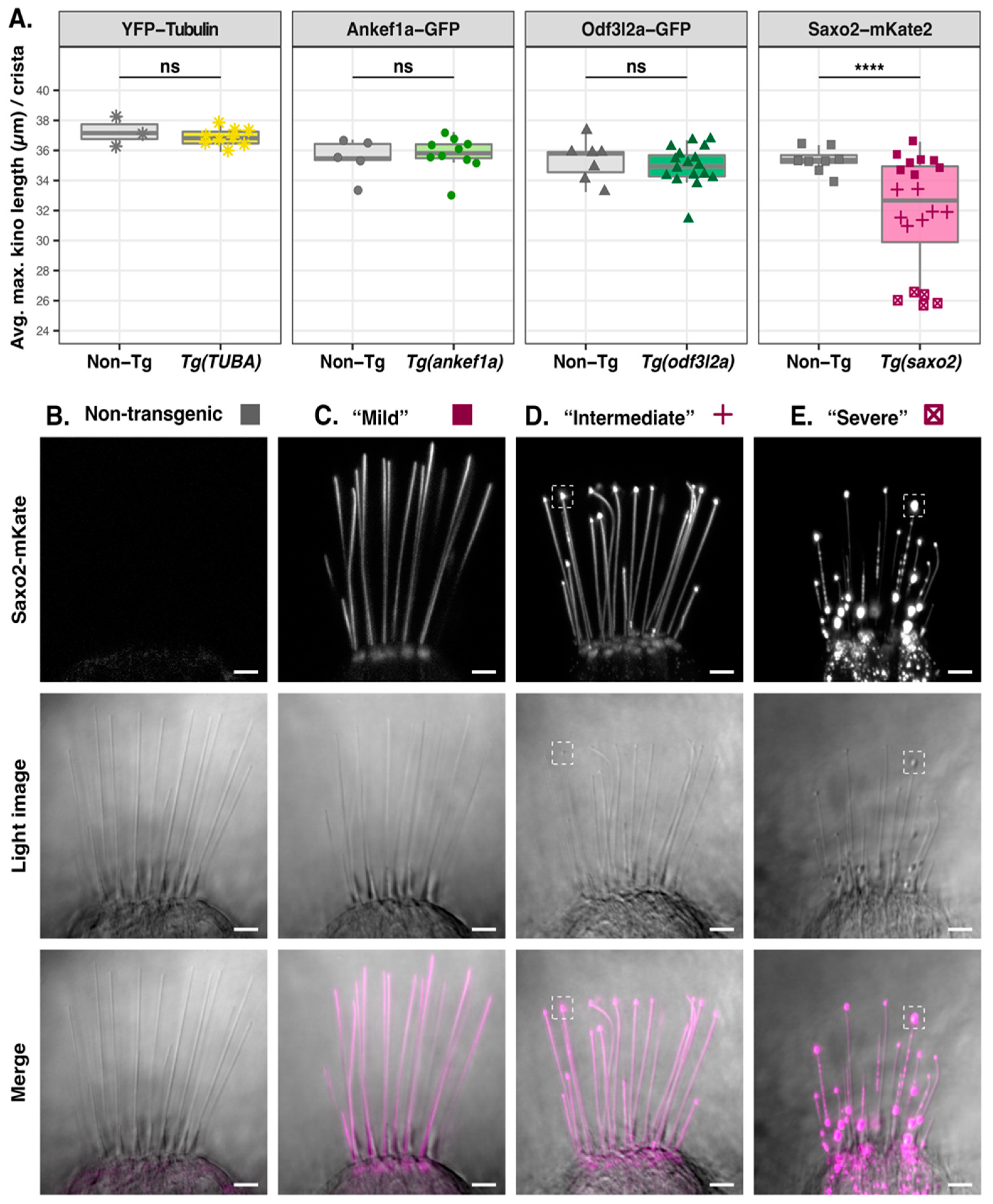
3.5. Saxo2-mKate2 Overexpression Results in Curved, Truncated, and ‘Bulbed’ Kinocilia
3.6. Ankef1a, Odf3l2a, and Tubulin Distribution Patterns Are Not Influenced by Saxo2
4. Discussion
4.1. Utility of TU-Tagging to Identify Hair Cell Transcripts
4.2. Determining the Subcellular Localization for Putative Kinocilial Proteins
4.3. Regionalization of the Hair Cell Kinocilium
4.4. Saxo2 Promotes Formation of Curved and Truncated Kinocilia
4.5. Evaluation of ANKEF1, ODF3L2, and SAXO2 as Putative Hearing Loss Genes
4.6. Benefits and Limitations of This Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bezares-Calderón, L.A.; Berger, J.; Jékely, G. Diversity of Cilia-Based Mechanosensory Systems and Their Functions in Marine Animal Behaviour. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190376. [Google Scholar] [CrossRef] [PubMed]
- Sharkova, M.; Chow, E.; Erickson, T.; Hocking, J.C. The Morphological and Functional Diversity of Apical Microvilli. J. Anat. 2022, 242, 327–353. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Roper, V.C.; Foucher, I.; Qian, D.; Banizs, B.; Petit, C.; Yoder, B.K.; Chen, P. Ciliary Proteins Link Basal Body Polarization to Planar Cell Polarity Regulation. Nat. Genet. 2008, 40, 69–77. [Google Scholar] [CrossRef]
- Moon, K.-H.; Ma, J.-H.; Min, H.; Koo, H.; Kim, H.; Ko, H.W.; Bok, J. Dysregulation of Sonic Hedgehog Signaling Causes Hearing Loss in Ciliopathy Mouse Models. Elife 2020, 9, e56551. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, H.; Zhang, C.; Lu, Y.; Yao, J.; Chen, Z.; Xing, G.; Wei, Q.; Cao, X. Mutations in OSBPL2 Cause Hearing Loss Associated with Primary Cilia Defects via Sonic Hedgehog Signaling. JCI Insight 2022, 7, e149626. [Google Scholar] [CrossRef]
- Baird, R.A. Comparative Transduction Mechanisms of Hair Cells in the Bullfrog Utriculus. II. Sensitivity and Response Dynamics to Hair Bundle Displacement. J. Neurophysiol. 1994, 71, 685–705. [Google Scholar] [CrossRef]
- Spoon, C.; Grant, W. Biomechanics of Hair Cell Kinocilia: Experimental Measurement of Kinocilium Shaft Stiffness and Base Rotational Stiffness with Euler–Bernoulli and Timoshenko Beam Analysis. J. Exp. Biol. 2011, 214, 862–870. [Google Scholar] [CrossRef] [Green Version]
- Kindt, K.S.; Finch, G.; Nicolson, T. Kinocilia Mediate Mechanosensitivity in Developing Zebrafish Hair Cells. Dev. Cell. 2012, 23, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Stooke-Vaughan, G.A.; Huang, P.; Hammond, K.L.; Schier, A.F.; Whitfield, T.T. The Role of Hair Cells, Cilia and Ciliary Motility in Otolith Formation in the Zebrafish Otic Vesicle. Development 2012, 139, 1777–1787. [Google Scholar] [CrossRef] [Green Version]
- Sobkowicz, H.M.; Slapnick, S.M.; August, B.K. The Kinocilium of Auditory Hair Cells and Evidence for Its Morphogenetic Role during the Regeneration of Stereocilia and Cuticular Plates. J. Neurocytol. 1995, 24, 633–653. [Google Scholar] [CrossRef]
- Flock, A.; Duvall, A.J. The Ultrastructure of the Kinocilium of the Sensory Cells in the Inner Ear and Lateral Line Organs. J. Cell Biol. 1965, 25, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, T.; Takasaka, T.; Tonosaki, A.; Watanabe, H. Fine Structure of Guinea Pig Vestibular Kinocilium. Acta Otolaryngol. 1989, 108, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Burton, P.R. Ultrastructural Studies of Microtubules and Microtubule Organizing Centers of the Vertebrate Olfactory Neuron. Microsc. Res. Tech. 1992, 23, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Rüsch, A.; Thurm, U. Passive and Active Deflections of Ampullary Kinocilia Correlated with Changes in Transepithelial Voltage. ORL 1986, 48, 76–80. [Google Scholar] [CrossRef]
- Rüsch, A.; Thurm, U. Spontaneous and Electrically Induced Movements of Ampullary Kinocilia and Stereovilli. Hear. Res. 1990, 48, 247–263. [Google Scholar] [CrossRef]
- Alstrom, C.H.; Hallgren, B.; Nilsson, L.; Asander, H. Retinal Degeneration Combined with Obesity, Diabetes Mellitus and Neurogenous Deafness: A Specific Syndrome (Not Hitherto Described) Distinct from the Laurence-Moon-Bardet-Biedl Syndrome: A Clinical, Endocrinological and Genetic Examination Based on a Large Pedigree. Acta Psychiatr. Neurol. Scand. Suppl. 1959, 129, 1–35. [Google Scholar]
- Ross, A.J.; May-Simera, H.; Eichers, E.R.; Kai, M.; Hill, J.; Jagger, D.J.; Leitch, C.C.; Chapple, J.P.; Munro, P.M.; Fisher, S.; et al. Disruption of Bardet-Biedl Syndrome Ciliary Proteins Perturbs Planar Cell Polarity in Vertebrates. Nat. Genet. 2005, 37, 1135–1140. [Google Scholar] [CrossRef]
- Jagger, D.; Collin, G.; Kelly, J.; Towers, E.; Nevill, G.; Longo-Guess, C.; Benson, J.; Halsey, K.; Dolan, D.; Marshall, J.; et al. Alström Syndrome Protein ALMS1 Localizes to Basal Bodies of Cochlear Hair Cells and Regulates Cilium-Dependent Planar Cell Polarity. Hum. Mol. Genet. 2011, 20, 466–481. [Google Scholar] [CrossRef] [Green Version]
- Modamio-Høybjør, S.; Mencía, Á.; Goodyear, R.; del Castillo, I.; Richardson, G.; Moreno, F.; Moreno-Pelayo, M.Á. A Mutation in CCDC50, a Gene Encoding an Effector of Epidermal Growth Factor-Mediated Cell Signaling, Causes Progressive Hearing Loss. Am. J. Hum. Genet. 2007, 80, 1076–1089. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Zheng, J.; Zhao, Q.; Chen, J.-R.; Liu, H.; Peng, G.; Wu, Y.; Chen, C.; He, Q.; Shi, H.; et al. Mutations of MAP1B Encoding a Microtubule-Associated Phosphoprotein Cause Sensorineural Hearing Loss. JCI Insight 2020, 5, e136046. [Google Scholar] [CrossRef]
- Grati, M.; Chakchouk, I.; Ma, Q.; Bensaid, M.; Desmidt, A.; Turki, N.; Yan, D.; Baanannou, A.; Mittal, R.; Driss, N.; et al. A Missense Mutation in DCDC2 Causes Human Recessive Deafness DFNB66, Likely by Interfering with Sensory Hair Cell and Supporting Cell Cilia Length Regulation. Hum. Mol. Genet. 2015, 24, 2482–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bork, J.M.; Peters, L.M.; Riazuddin, S.; Ahmed, Z.M.; Li, X.C.; Griffith, A.J.; Wilcox, E.R.; Friedman, T.B.; Morell, R.J.; Griffith, A.J.; et al. Usher Syndrome 1D and Nonsyndromic Autosomal Recessive Deafness DFNB12 Are Caused by Allelic Mutations of the Novel Cadherin-like Gene CDH23. Am. J. Hum. Genet. 2001, 68, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, Z.M. PCDH15 Is Expressed in the Neurosensory Epithelium of the Eye and Ear and Mutant Alleles Are Responsible for Both USH1F and DFNB23. Hum. Mol. Genet. 2003, 12, 3215–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, L.; Zallocchi, M. Integrin A8 and Pcdh15 Act as a Complex to Regulate Cilia Biogenesis in Sensory Cells. J. Cell Sci. 2017, 130, 3698–3712. [Google Scholar] [CrossRef] [Green Version]
- Pickett, S.B.; Raible, D.W. Water Waves to Sound Waves: Using Zebrafish to Explore Hair Cell Biology. J. Assoc. Res. Otolaryngol. 2019, 20, 1–19. [Google Scholar] [CrossRef]
- Gunn, R.K.; Keenan, M.-E.; Brown, R.E. Analysis of Sensory, Motor and Cognitive Functions of the Coloboma (C3Sn.Cg-Cm/J) Mutant Mouse. Genes. Brain Behav. 2011, 10, 579–588. [Google Scholar] [CrossRef]
- Kiernan, A.E.; Ahituv, N.; Fuchs, H.; Balling, R.; Avraham, K.B.; Steel, K.P.; Hrabé de Angelis, M. The Notch Ligand Jagged1 Is Required for Inner Ear Sensory Development. Proc. Natl. Acad. Sci. USA 2001, 98, 3873–3878. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Scheffer, D.I.; Kwan, K.Y.; Corey, D.P. SHIELD: An Integrative Gene Expression Database for Inner Ear Research. Database 2015, 2015, bav071. [Google Scholar] [CrossRef] [Green Version]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 4th ed.; University of Oregon: Eugene OR, USA, 2000. [Google Scholar]
- Ohta, S.; Ji, Y.R.; Martin, D.; Wu, D.K. Emx2 Regulates Hair Cell Rearrangement but Not Positional Identity within Neuromasts. Elife 2020, 9, e60432. [Google Scholar] [CrossRef]
- Erickson, T.; Pacentine, I.V.; Venuto, A.; Clemens, R.; Nicolson, T. The Lhfpl5 Ohnologs Lhfpl5a and Lhfpl5b Are Required for Mechanotransduction in Distinct Populations of Sensory Hair Cells in Zebrafish. Front. Mol. Neurosci. 2020, 12, 320. [Google Scholar] [CrossRef] [Green Version]
- Erickson, T.; Nicolson, T. Identification of Sensory Hair-Cell Transcripts by Thiouracil-Tagging in Zebrafish. BMC Genom. 2015, 16, 842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.S.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C.-B. The Tol2kit: A Multisite Gateway-Based Construction Kit for Tol2 Transposon Transgenesis Constructs. Dev. Dyn. 2007, 236, 3088–3099. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2016 Update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.-P.; Mi, H. PANTHER: Making Genome-Scale Phylogenetics Accessible to All. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Huang, X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol Update for Large-Scale Genome and Gene Function Analysis with the PANTHER Classification System (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef] [PubMed]
- Van Camp, G.; Smith, R.J.H. Hereditary Hearing Loss Homepage. Available online: https://hereditaryhearingloss.org (accessed on 29 December 2022).
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]
- Daniel, J.G.; Panizzi, J.R. Spatiotemporal Expression Profile of Embryonic and Adult Ankyrin Repeat and EF-Hand Domain Containing Protein 1-Encoding Genes Ankef1a and Ankef1b in Zebrafish. Gene Expr. Patterns 2019, 34, 119069. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Arshadi, C.; Günther, U.; Eddison, M.; Harrington, K.I.S.; Ferreira, T.A. SNT: A Unifying Toolbox for Quantification of Neuronal Anatomy. Nat. Methods 2021, 18, 374–377. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio PBC: Boston, MA, USA, 2022. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Miller, M.R.; Robinson, K.J.; Cleary, M.D.; Doe, C.Q. TU-Tagging: Cell Type-Specific RNA Isolation from Intact Complex Tissues. Nat. Methods 2009, 6, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.K.; Mason, L.; Arrenberg, A.B.; Ziv, L.; Gosse, N.J.; Xiao, T.; Chi, N.C.; Asakawa, K.; Kawakami, K.; Baier, H. Targeting Neural Circuitry in Zebrafish Using GAL4 Enhancer Trapping. Nat. Methods 2007, 4, 323–326. [Google Scholar] [CrossRef]
- Chung, H.A.; Yamamoto, T.S.; Ueno, N. ANR5, an FGF Target Gene Product, Regulates Gastrulation in Xenopus. Curr. Biol. 2007, 17, 932–939. [Google Scholar] [CrossRef] [Green Version]
- Di Domenico, F. The Role of Ankef1 in Male Fertility. Ph.D. Thesis, Icahn School of Medicine at Mount Sinai, New York, NY, USA, 2021. [Google Scholar]
- Jan, T.A.; Eltawil, Y.; Ling, A.H.; Chen, L.; Ellwanger, D.C.; Heller, S.; Cheng, A.G. Spatiotemporal Dynamics of Inner Ear Sensory and Non-Sensory Cells Revealed by Single-Cell Transcriptomics. Cell Rep. 2021, 36, 109358. [Google Scholar] [CrossRef] [PubMed]
- Hoh, R.A.; Stowe, T.R.; Turk, E.; Stearns, T. Transcriptional Program of Ciliated Epithelial Cells Reveals New Cilium and Centrosome Components and Links to Human Disease. PLoS ONE 2012, 7, e52166. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.A.; Hetherington, L.; Reeves, G.M.; Aitken, R.J. The Mouse Sperm Proteome Characterizedvia IPG Strip Prefractionation and LC-MS/MS Identification. Proteomics 2008, 8, 1720–1730. [Google Scholar] [CrossRef]
- Baker, M.A.; Hetherington, L.; Reeves, G.; Müller, J.; Aitken, R.J. The Rat Sperm Proteome Characterized via IPG Strip Prefractionation and LC-MS/MS Identification. Proteomics 2008, 8, 2312–2321. [Google Scholar] [CrossRef]
- Blackburn, K.; Bustamante-Marin, X.; Yin, W.; Goshe, M.B.; Ostrowski, L.E. Quantitative Proteomic Analysis of Human Airway Cilia Identifies Previously Uncharacterized Proteins of High Abundance. J. Proteome Res. 2017, 16, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Elkon, R.; Milon, B.; Morrison, L.; Shah, M.; Vijayakumar, S.; Racherla, M.; Leitch, C.C.; Silipino, L.; Hadi, S.; Weiss-Gayet, M.; et al. RFX Transcription Factors Are Essential for Hearing in Mice. Nat. Commun. 2015, 6, 8549. [Google Scholar] [CrossRef] [Green Version]
- Lush, M.E.; Diaz, D.C.; Koenecke, N.; Baek, S.; Boldt, H.; St Peter, M.K.; Gaitan-Escudero, T.; Romero-Carvajal, A.; Busch-Nentwich, E.M.; Perera, A.G.; et al. ScRNA-Seq Reveals Distinct Stem Cell Populations That Drive Hair Cell Regeneration after Loss of Fgf and Notch Signaling. Elife 2019, 8, e44431. [Google Scholar] [CrossRef] [PubMed]
- Barta, C.L.; Liu, H.; Chen, L.; Giffen, K.P.; Li, Y.; Kramer, K.L.; Beisel, K.W.; He, D.Z. RNA-Seq Transcriptomic Analysis of Adult Zebrafish Inner Ear Hair Cells. Sci. Data 2018, 5, 180005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, E.J.; Collins, K.A.; Copeland, N.G.; Jenkins, N.A.; Wilson, M.C. Deletion Map of the Coloboma (Cm) Locus on Mouse Chromosome 2. Genomics 1994, 21, 257–261. [Google Scholar] [CrossRef]
- Blake, J.A.; Baldarelli, R.; Kadin, J.A.; Richardson, J.E.; Smith, C.L.; Bult, C.J.; Anagnostopoulos, A.v.; Beal, J.S.; Bello, S.M.; Blodgett, O.; et al. Mouse Genome Database (MGD): Knowledgebase for Mouse–Human Comparative Biology. Nucleic Acids Res. 2021, 49, D981–D987. [Google Scholar] [CrossRef] [PubMed]
- Hess, E.; Jinnah, H.; Kozak, C.; Wilson, M. Spontaneous Locomotor Hyperactivity in a Mouse Mutant with a Deletion Including the Snap Gene on Chromosome 2. J. Neurosci. 1992, 12, 2865–2874. [Google Scholar] [CrossRef] [Green Version]
- Egydio de Carvalho, C.; Tanaka, H.; Iguchi, N.; Ventelä, S.; Nojima, H.; Nishimune, Y. Molecular Cloning and Characterization of a Complementary DNA Encoding Sperm Tail Protein SHIPPO 11. Biol. Reprod. 2002, 66, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Dorus, S.; Busby, S.A.; Gerike, U.; Shabanowitz, J.; Hunt, D.F.; Karr, T.L. Genomic and Functional Evolution of the Drosophila melanogaster Sperm Proteome. Nat. Genet. 2006, 38, 1440–1445. [Google Scholar] [CrossRef]
- Konno, A.; Padma, P.; Ushimaru, Y.; Inaba, K. Multidimensional Analysis of Uncharacterized Sperm Proteins in Ciona intestinalis: EST-Based Analysis and Functional Immunoscreening of Testis-Expressed Genes. Zoolog. Sci. 2010, 27, 204–215. [Google Scholar] [CrossRef]
- Konno, A.; Shiba, K.; Cai, C.; Inaba, K. Branchial Cilia and Sperm Flagella Recruit Distinct Axonemal Components. PLoS ONE 2015, 10, e0126005. [Google Scholar] [CrossRef] [Green Version]
- White-Cooper, H. Studying How Flies Make Sperm—Investigating Gene Function in Drosophila Testes. Mol. Cell Endocrinol. 2009, 306, 66–74. [Google Scholar] [CrossRef]
- Bastin, B.R. Evolution of Tektin and ODF3 Family Genes and the Role of Gene Duplication in the Specialization of Motile Ciliary Structures in the Polychaete Platynereis dumerilii. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2018. [Google Scholar]
- Lauwaet, T.; Smith, A.J.; Reiner, D.S.; Romijn, E.P.; Wong, C.C.L.; Davids, B.J.; Shah, S.A.; Yates, J.R.; Gillin, F.D. Mining the Giardia Genome and Proteome for Conserved and Unique Basal Body Proteins. Int. J. Parasitol. 2011, 41, 1079–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, K.; Leung, J.W.; Li, Y.; Wang, W.; Feng, L.; Ma, W.; Liu, D.; Songyang, Z.; Chen, J. MTR120/KIAA1383, a Novel Microtubule-Associated Protein, Promotes Microtubule Stability and Ensures Cytokinesis. J. Cell Sci. 2013, 126, 825–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, S.; Black, C.S.; Joachimiak, E.; Yang, S.K.; Legal, T.; Peri, K.; Abdelzaher, A.; Khalifa, Z.; Ghanaeian, A.; Mccafferty, C.L.; et al. Native Doublet Microtubules from Tetrahymena thermophila Reveal the Importance of Outer Junction Proteins. Nat. Commun. 2023, 14, 2168. [Google Scholar] [CrossRef]
- Bowl, M.R.; Simon, M.M.; Ingham, N.J.; Greenaway, S.; Santos, L.; Cater, H.; Taylor, S.; Mason, J.; Kurbatova, N.; Pearson, S.; et al. A Large Scale Hearing Loss Screen Reveals an Extensive Unexplored Genetic Landscape for Auditory Dysfunction. Nat. Commun. 2017, 8, 886. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.U.; Gul, K.; Morell, R.J.; Lee, K.; Ahmed, Z.M.; Riazuddin, S.; Ali, R.A.; Shahzad, M.; Jaleel, A.; Andrade, P.B.; et al. Mutations of GIPC3 Cause Nonsyndromic Hearing Loss DFNB72 but Not DFNB81 That Also Maps to Chromosome 19p. Hum. Genet. 2011, 130, 759–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charizopoulou, N.; Lelli, A.; Schraders, M.; Ray, K.; Hildebrand, M.S.; Ramesh, A.; Srisailapathy, C.R.S.; Oostrik, J.; Admiraal, R.J.C.; Neely, H.R.; et al. Gipc3 Mutations Associated with Audiogenic Seizures and Sensorineural Hearing Loss in Mouse and Human. Nat. Commun. 2011, 2, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Cortez, R.L.P.; Faridi, R.; Rehman, A.U.; Lee, K.; Ansar, M.; Wang, X.; Morell, R.J.; Isaacson, R.; Belyantseva, I.A.; Dai, H.; et al. Autosomal-Recessive Hearing Impairment Due to Rare Missense Variants within S1PR2. Am. J. Hum. Genet. 2016, 98, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, E.M.; Rehman, A.U.; Walsh, T.; Clayton-Smith, J.; Lee, K.; Morell, R.J.; Drummond, M.C.; Khan, S.N.; Naeem, M.A.; Rauf, B.; et al. Perrault Syndrome Is Caused by Recessive Mutations in CLPP, Encoding a Mitochondrial ATP-Dependent Chambered Protease. Am. J. Hum. Genet. 2013, 92, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Bönsch, D.; Schmidt, C.M.; Scheer, P.; Bohlender, J.; Neumann, C.; am Zehnhoff-Dinnesen, A.; Deufel, T. Ein Neuer Genort Für Eine Autosomal Dominante, Nichtsyndromale Hörstörung (DFNA57) Kartiert Auf Chromosom 19p13.2 Und Überlappt Mit DFNB15. HNO 2008, 56, 177–182. [Google Scholar] [CrossRef]
- Ain, Q.; Nazli, S.; Riazuddin, S.; Jaleel, A.; Riazuddin, S.A.; Zafar, A.U.; Khan, S.N.; Husnain, T.; Griffith, A.J.; Ahmed, Z.M.; et al. The Autosomal Recessive Nonsyndromic Deafness Locus DFNB72 Is Located on Chromosome 19p13.3. Hum. Genet. 2007, 122, 445–450. [Google Scholar] [CrossRef]
- Mangino, M.; Flex, E.; Capon, F.; Sangiuolo, F.; Carraro, E.; Gualandi, F.; Mazzoli, M.; Martini, A.; Novelli, G.; Dallapiccola, B. Mapping of a New Autosomal Dominant Nonsyndromic Hearing Loss Locus (DFNA30) to Chromosome 15q25-26. Eur. J. Human. Genet. 2001, 9, 667–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuveillier, C.; Boulan, B.; Ravanello, C.; Denarier, E.; Deloulme, J.-C.; Gory-Fauré, S.; Delphin, C.; Bosc, C.; Arnal, I.; Andrieux, A. Beyond Neuronal Microtubule Stabilization: MAP6 and CRMPS, Two Converging Stories. Front. Mol. Neurosci. 2021, 14, 82. [Google Scholar] [CrossRef]
- Dacheux, D.; Landrein, N.; Thonnus, M.; Gilbert, G.; Sahin, A.; Wodrich, H.; Robinson, D.R.; Bonhivers, M. A MAP6-Related Protein Is Present in Protozoa and Is Involved in Flagellum Motility. PLoS ONE 2012, 7, e31344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacheux, D.; Roger, B.; Bosc, C.; Landrein, N.; Roche, E.; Chansel, L.; Trian, T.; Andrieux, A.; Papaxanthos-Roche, A.; Marthan, R.; et al. Human FAM154A (SAXO1) Is a Microtubule-Stabilizing Protein Specific to Cilia and Related Structures. J. Cell Sci. 2015, 128, 1294–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyeneho, I.A. Microtubule Stability Mediated by New Mammalian STOP Family Members. Ph.D. Thesis, Stanford University, Stanford, CA, USA, 2014. [Google Scholar]
- Kim, K.; Gibboney, S.; Razy-Krajka, F.; Lowe, E.K.; Wang, W.; Stolfi, A. Regulation of Neurogenesis by FGF Signaling and Neurogenin in the Invertebrate Chordate Ciona. Front. Cell Dev. Biol. 2020, 8, 477. [Google Scholar] [CrossRef]
- Leung, M.R.; Roelofs, M.C.; Chiozzi, R.Z.; Hevler, J.F.; Heck, A.J.R.; Zeev-Ben-Mordehai, T. Unraveling the Intricate Microtubule Inner Protein Networks That Reinforce Mammalian Sperm Flagella. bioRxiv 2022. [Google Scholar] [CrossRef]
- Gui, M.; Wang, X.; Dutcher, S.K.; Brown, A.; Zhang, R. Ciliary Central Apparatus Structure Reveals Mechanisms of Microtubule Patterning. Nat. Struct. Mol. Biol. 2022, 29, 483–492. [Google Scholar] [CrossRef]
- Gui, M.; Croft, J.T.; Zabeo, D.; Acharya, V.; Kollman, J.M.; Burgoyne, T.; Höög, J.L.; Brown, A. SPACA9 Is a Lumenal Protein of Human Ciliary Singlet and Doublet Microtubules. Proc. Natl. Acad. Sci. USA 2022, 119, e2207605119. [Google Scholar] [CrossRef]
- Ichikawa, M.; Bui, K.H. Microtubule Inner Proteins: A Meshwork of Luminal Proteins Stabilizing the Doublet Microtubule. BioEssays 2018, 40, 1700209. [Google Scholar] [CrossRef]
- Cuveillier, C.; Delaroche, J.; Seggio, M.; Gory-Fauré, S.; Bosc, C.; Denarier, E.; Bacia, M.; Schoehn, G.; Mohrbach, H.; Kulić, I.; et al. MAP6 Is an Intraluminal Protein That Induces Neuronal Microtubules to Coil. Sci. Adv. 2020, 6, eaaz4344. [Google Scholar] [CrossRef] [Green Version]
- Wilkerson, B.A.; Zebroski, H.L.; Finkbeiner, C.R.; Chitsazan, A.D.; Beach, K.E.; Sen, N.; Zhang, R.C.; Bermingham-McDonogh, O. Novel Cell Types and Developmental Lineages Revealed by Single-Cell RNA-Seq Analysis of the Mouse Crista ampullaris. Elife 2021, 10, e60108. [Google Scholar] [CrossRef]
- Azaiez, H.; Decker, A.R.; Booth, K.T.; Simpson, A.C.; Shearer, A.E.; Huygen, P.L.M.; Bu, F.; Hildebrand, M.S.; Ranum, P.T.; Shibata, S.B.; et al. HOMER2, a Stereociliary Scaffolding Protein, Is Essential for Normal Hearing in Humans and Mice. PLoS Genet. 2015, 11, e1005137. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Wang, Q.; Gu, H.; Zhang, X.; Qi, Y.; Liu, Y. Whole Exome Sequencing Identified a Second Pathogenic Variant in HOMER2 for Autosomal Dominant Non-Syndromic Deafness. Clin. Genet. 2018, 94, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, J.; Khan, S.N.; Khan, S.Y.; Ramzan, K.; Riazuddin, S.; Ahmed, Z.M.; Wilcox, E.R.; Friedman, T.B.; Riazuddin, S. DFNB48, a New Nonsyndromic Recessive Deafness Locus, Maps to Chromosome 15q23-Q25.1. Hum. Genet. 2005, 116, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Riazuddin, S.; Belyantseva, I.A.; Giese, A.P.J.; Lee, K.; Indzhykulian, A.A.; Nandamuri, S.P.; Yousaf, R.; Sinha, G.P.; Lee, S.; Terrell, D.; et al. Alterations of the CIB2 Calcium- and Integrin-Binding Protein Cause Usher Syndrome Type 1J and Nonsyndromic Deafness DFNB48. Nat. Genet. 2012, 44, 1265–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomek, M.S.; Brown, M.R.; Mani, S.R.; Ramesh, A.; Srisailapathy, C.R.; Coucke, P.; Zbar, R.I.; Bell, A.M.; McGuirt, W.T.; Fukushima, K.; et al. Localization of a Gene for Otosclerosis to Chromosome 15q25-Q26. Hum. Mol. Genet. 1998, 7, 285–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Hazelrigg, T. Implications for Bcd MRNA Localization from Spatial Distribution of Exu Protein in Drosophila oogenesis. Nature 1994, 369, 400–403. [Google Scholar] [CrossRef]
- Gerdes, H.-H.; Kaether, C. Green Fluorescent Protein: Applications in Cell Biology. FEBS Lett. 1996, 389, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Sheets, L.; He, X.J.; Olt, J.; Schreck, M.; Petralia, R.S.; Wang, Y.-X.; Zhang, Q.; Beirl, A.; Nicolson, T.; Marcotti, W.; et al. Enlargement of Ribbons in Zebrafish Hair Cells Increases Calcium Currents But Disrupts Afferent Spontaneous Activity and Timing of Stimulus Onset. J. Neurosci. 2017, 37, 6299–6313. [Google Scholar] [CrossRef]
- Maeda, R.; Pacentine, I.V.; Erickson, T.; Nicolson, T. Functional Analysis of the Transmembrane and Cytoplasmic Domains of Pcdh15a in Zebrafish Hair Cells. J. Neurosci. 2017, 37, 3231–3245. [Google Scholar] [CrossRef] [Green Version]
- Erickson, T.; Morgan, C.P.; Olt, J.; Hardy, K.; Busch-Nentwich, E.; Maeda, R.; Clemens, R.; Krey, J.F.; Nechiporuk, A.; Barr-Gillespie, P.G.; et al. Integration of Tmc1/2 into the Mechanotransduction Complex in Zebrafish Hair Cells Is Regulated by Transmembrane O-Methyltransferase (Tomt). Elife 2017, 6, e28474. [Google Scholar] [CrossRef] [PubMed]
- Pacentine, I.v.; Nicolson, T. Subunits of the Mechano-Electrical Transduction Channel, Tmc1/2b, Require Tmie to Localize in Zebrafish Sensory Hair Cells. PLoS Genet. 2019, 15, e1007635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopal, S.R.; Chen, D.H.-C.; Chou, S.-W.; Zang, J.; Neuhauss, S.C.F.; Stepanyan, R.; McDermott, B.M.; Alagramam, K.N. Zebrafish Models for the Mechanosensory Hair Cell Dysfunction in Usher Syndrome 3 Reveal That Clarin-1 Is an Essential Hair Bundle Protein. J. Neurosci. 2015, 35, 10188–10201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.T.; Pacentine, I.; Shipman, A.; Hill, M.; Nicolson, T. Disruption of Tmc1/2a/2b Genes in Zebrafish Reveals Subunit Requirements in Subtypes of Inner Ear Hair Cells. J. Neurosci. 2020, 40, 4457–4468. [Google Scholar] [CrossRef]
- Chen, Z.; Chou, S.-W.; McDermott, B.M. Ribeye Protein Is Intrinsically Dynamic but Is Stabilized in the Context of the Ribbon Synapse. J. Physiol. 2018, 596, 409–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.; Chou, S.-W.; Chen, Z.; McDermott, B.M. The Stereociliary Paracrystal Is a Dynamic Cytoskeletal Scaffold In Vivo. Cell Rep. 2015, 13, 1287–1294. [Google Scholar] [CrossRef] [Green Version]
- Sheets, L.; Hagen, M.W.; Nicolson, T. Characterization of Ribeye Subunits in Zebrafish Hair Cells Reveals That Exogenous Ribeye B-Domain and CtBP1 Localize to the Basal Ends of Synaptic Ribbons. PLoS ONE 2014, 9, e107256. [Google Scholar] [CrossRef] [Green Version]
- Choksi, S.P.; Babu, D.; Lau, D.; Yu, X.; Roy, S. Systematic Discovery of Novel Ciliary Genes through Functional Genomics in the Zebrafish. Development 2014, 141, 3410–3419. [Google Scholar] [CrossRef] [Green Version]
- Walentek, P.; Quigley, I.K.; Sun, D.I.; Sajjan, U.K.; Kintner, C.; Harland, R.M. Ciliary Transcription Factors and MiRNAs Precisely Regulate Cp110 Levels Required for Ciliary Adhesions and Ciliogenesis. Elife 2016, 5, e17557. [Google Scholar] [CrossRef]
- Stauber, M.; Weidemann, M.; Dittrich-Breiholz, O.; Lobschat, K.; Alten, L.; Mai, M.; Beckers, A.; Kracht, M.; Gossler, A. Identification of FOXJ1 Effectors during Ciliogenesis in the Foetal Respiratory Epithelium and Embryonic Left-Right Organiser of the Mouse. Dev. Biol. 2017, 423, 170–188. [Google Scholar] [CrossRef]
- Alsolami, M.; Kuhns, S.; Alsulami, M.; Blacque, O.E. ERICH3 in Primary Cilia Regulates Cilium Formation and the Localisations of Ciliary Transport and Sonic Hedgehog Signaling Proteins. Sci. Rep. 2019, 9, 16519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imtiaz, A.; Belyantseva, I.A.; Beirl, A.J.; Fenollar-Ferrer, C.; Bashir, R.; Bukhari, I.; Bouzid, A.; Shaukat, U.; Azaiez, H.; Booth, K.T.; et al. CDC14A Phosphatase Is Essential for Hearing and Male Fertility in Mouse and Human. Hum. Mol. Genet. 2018, 27, 780–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwenk, B.M.; Lang, C.M.; Hogl, S.; Tahirovic, S.; Orozco, D.; Rentzsch, K.; Lichtenthaler, S.F.; Hoogenraad, C.C.; Capell, A.; Haass, C.; et al. The FTLD Risk Factor TMEM106B and MAP6 Control Dendritic Trafficking of Lysosomes. EMBO J. 2013, 33, 450–467. [Google Scholar] [CrossRef] [PubMed]
- Gory-Fauré, S.; Windscheid, V.; Brocard, J.; Montessuit, S.; Tsutsumi, R.; Denarier, E.; Fukata, Y.; Bosc, C.; Delaroche, J.; Collomb, N.; et al. Non-Microtubular Localizations of Microtubule-Associated Protein 6 (MAP6). PLoS ONE 2014, 9, e114905. [Google Scholar] [CrossRef]
- Tortosa, E.; Adolfs, Y.; Fukata, M.; Pasterkamp, R.J.; Kapitein, L.C.; Hoogenraad, C.C. Dynamic Palmitoylation Targets MAP6 to the Axon to Promote Microtubule Stabilization during Neuronal Polarization. Neuron 2017, 94, 809–825.e7. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Insinna, C.; Ott, C.; Stauffer, J.; Pintado, P.A.; Rahajeng, J.; Baxa, U.; Walia, V.; Cuenca, A.; Hwang, Y.S.; et al. Early Steps in Primary Cilium Assembly Require EHD1/EHD3-Dependent Ciliary Vesicle Formation. Nat. Cell Biol. 2015, 17, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Chung, Y.D. Ciliary Subcompartments: How Are They Established and What Are Their Functions? BMB Rep. 2015, 48, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Soares, H.; Carmona, B.; Nolasco, S.; Melo, L.V.; Gonçalves, J. Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures. Cells 2019, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Gui, M.; Farley, H.; Anujan, P.; Anderson, J.R.; Maxwell, D.W.; Whitchurch, J.B.; Botsch, J.J.; Qiu, T.; Meleppattu, S.; Singh, S.K.; et al. De Novo Identification of Mammalian Ciliary Motility Proteins Using Cryo-EM. Cell 2021, 184, 5791–5806.e19. [Google Scholar] [CrossRef]
- Lagziel, A.; Ahmed, Z.M.; Schultz, J.M.; Morell, R.J.; Belyantseva, I.A.; Friedman, T.B. Spatiotemporal Pattern and Isoforms of Cadherin 23 in Wild Type and Waltzer Mice during Inner Ear Hair Cell Development. Dev. Biol. 2005, 280, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Nicolson, T.; Rüsch, A.; Friedrich, R.W.; Granato, M.; Ruppersberg, J.P.; Nüsslein-Volhard, C. Genetic Analysis of Vertebrate Sensory Hair Cell Mechanosensation: The Zebrafish Circler Mutants. Neuron 1998, 20, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Lanford, P.J.; Lan, Y.; Jiang, R.; Lindsell, C.; Weinmaster, G.; Gridley, T.; Kelley, M.W. Notch Signalling Pathway Mediates Hair Cell Development in Mammalian Cochlea. Nat. Genet. 1999, 21, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Zine, A.; van de Water, T.R.; de Ribaupierre, F. Notch Signaling Regulates the Pattern of Auditory Hair Cell Differentiation in Mammals. Development 2000, 127, 3373–3383. [Google Scholar] [CrossRef] [PubMed]
- Bowl, M.R.; Brown, S.D.M. Genetic Landscape of Auditory Dysfunction. Hum. Mol. Genet. 2018, 27, R130–R135. [Google Scholar] [CrossRef] [PubMed]
- Groza, T.; Gomez, F.L.; Mashhadi, H.H.; Muñoz-Fuentes, V.; Gunes, O.; Wilson, R.; Cacheiro, P.; Frost, A.; Keskivali-Bond, P.; Vardal, B.; et al. The International Mouse Phenotyping Consortium: Comprehensive Knockout Phenotyping Underpinning the Study of Human Disease. Nucleic Acids Res. 2022, 51, D1038–D1045. [Google Scholar] [CrossRef]
- Kolla, L.; Kelly, M.C.; Mann, Z.F.; Anaya-Rocha, A.; Ellis, K.; Lemons, A.; Palermo, A.T.; So, K.S.; Mays, J.C.; Orvis, J.; et al. Characterization of the Development of the Mouse Cochlear Epithelium at the Single Cell Level. Nat. Commun. 2020, 11, 2389. [Google Scholar] [CrossRef]
- Kubota, M.; Scheibinger, M.; Jan, T.A.; Heller, S. Greater Epithelial Ridge Cells Are the Principal Organoid-Forming Progenitors of the Mouse Cochlea. Cell Rep. 2021, 34, 108646. [Google Scholar] [CrossRef]
- Steiner, A.B.; Kim, T.; Cabot, V.; Hudspeth, A.J. Dynamic Gene Expression by Putative Hair-Cell Progenitors during Regeneration in the Zebrafish Lateral Line. Proc. Natl. Acad. Sci. USA 2014, 111, E1393–E1401. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Pathak, N.; Kramer-Zucker, A.; Drummond, I.A. Notch Signaling Controls the Differentiation of Transporting Epithelia and Multiciliated Cells in the Zebrafish Pronephros. Development 2007, 134, 1111–1122. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-C.; Hou, M.-F.; Hong, J.-R.; Wu, J.-L.; Her, G.M. Inducible Male Infertility by Targeted Cell Ablation in Zebrafish Testis. Mar. Biotechnol. 2010, 12, 466–478. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erickson, T.; Biggers, W.P., III; Williams, K.; Butland, S.E.; Venuto, A. Regionalized Protein Localization Domains in the Zebrafish Hair Cell Kinocilium. J. Dev. Biol. 2023, 11, 28. https://doi.org/10.3390/jdb11020028
Erickson T, Biggers WP III, Williams K, Butland SE, Venuto A. Regionalized Protein Localization Domains in the Zebrafish Hair Cell Kinocilium. Journal of Developmental Biology. 2023; 11(2):28. https://doi.org/10.3390/jdb11020028
Chicago/Turabian StyleErickson, Timothy, William Paul Biggers, III, Kevin Williams, Shyanne E. Butland, and Alexandra Venuto. 2023. "Regionalized Protein Localization Domains in the Zebrafish Hair Cell Kinocilium" Journal of Developmental Biology 11, no. 2: 28. https://doi.org/10.3390/jdb11020028