
Comparison of Pronase versus Manual Dechorionation of Zebrafish Embryos for Small Molecule Treatments


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solutions
| ICS Water | ||
| Reagent | Quantity | Final Concentration |
| Instant Ocean Salt | 5.97 g | 300 mg/L |
| CaCl2 | 1.26 g | 0.56 mM |
| NaHCO3 | 1.9 g | 1.2 mM |
| RO water | Up to 20 L | |
| Pronase Stock Solution | |||
| Reagent | Source | Quantity | Final Concentration |
| Pronase from Streptomyces griseus | Roche Applied Science | 1 g | 20 mg/mL |
| RO water | Up to 50 mL | ||
| Make 1 mL aliquots and store at −20 °C. | |||
| Embryo Medium 20X Stock Solution | |||
| Reagent | Quantity | Final Concentration | |
| NaCl | 17.5 g | 15 mM | |
| KCl | 0.75 g | 0.50 mM | |
| CaCl2·2H2O | 2.9 g | 1 mM | |
| KH2PO4 | 0.41 g | 0.15 mM | |
| Na2HPO4 | 0.142 g | 0.05 mM | |
| MgSO4·7H2O | 4.9 g | 1 mM | |
| RO water | Up to 1 L | ||
| Vacuum filter and store at 4 °C. | |||
| Sodium Bicarbonate 1000X Stock Solution | |||
| Reagent | Quantity | Final Concentration | |
| NaHCO3 | 0.70 g | 0.83 M | |
| RO water | Up to 10 mL | ||
| Make 1 mL aliquots and store at −20 °C. | |||
| Embryo Medium 1X Stock Solution | |||
| Reagent | Quantity | Final Concentration | |
| 20X Embryo medium | 50 mL | 1X | |
| 1000X Sodium bicarbonate | 1 mL | 0.83 mM | |
| RO water | Up to 1 L | ||
| Store at room temperature and use within 1 week. | |||
| TSA Treatment Solution | |||
| Reagent | Source | Quantity | Final Concentration |
| TSA, 1 mM in DMSO | Sigma-Aldrich | 12 μL | 200 nM |
| DMSO | Sigma-Aldrich | 48 μL | 0.0704 M (0.5%) |
| 1X Embryo medium | Up to 12 mL | ||
| Make fresh each day. | |||
| Salermide + Oxamflatin Treatment Solution | |||
| Reagent | Source | Quantity | Final Concentration |
| Salermide + oxamflatin, 1 mM each in DMSO | Cayman Chemical | 60 μL | 5 μM each |
| 1X Embryo medium | 12 mL | ||
| Make fresh each day. | |||
| Doxorubicin treatment solution | |||
| Reagent | Source | Quantity | Final concentration |
| Doxorubicin, 10 mM in DMSO | Sigma-Aldrich | 10 μL | 100 μM |
| 1X Embryo medium | 1 mL | ||
| Make fresh each day. | |||
| DMSO Control Treatment Solution | |||
| Reagent | Source | Quantity | Final Concentration |
| DMSO | Sigma-Aldrich | 60 μL | 0.0704 M (0.5%) |
| 1X Embryo medium | Up to 12 mL | ||
| Make fresh each day. | |||
2.2. Zebrafish Husbandry
2.3. Dechorionation
- To make 0.5 mg/mL pronase solution, mix 125 µL of 20 mg/mL pronase stock solution in 5 mL RO water in a 15 mL tube.
- Using a Pasteur pipette, transfer up to 100 24 hpf embryos to a 60 mm × 15 mm Petri dish (Figure 1B). Remove excess ICS water.
- Pour in 5 mL of 0.5 mg/mL pronase solution per dish and immediately start a timer set for 14 min.
- Swirl the dish 3−4X during the treatment incubation. A few chorions may begin coming off during the last 2–3 min of incubation.
- Immediately after the timer goes off, carefully pour off most of the liquid into a waste container. Be sure to not lose any embryos. Some liquid can be left in the dish.
- Rinse the embryos by immediately adding about 10 mL of ICS water to the dish. A plastic wash bottle may be used.
- Using a plastic transfer pipette (Fisher Scientific 13-711-7M), very gently pipette the ICS water and embryos up and down about 5X to remove any remaining chorions.
- Repeat the ICS water rinse two more times to remove chorions and residual pronase.
- Using a stereomicroscope, confirm that all of the embryos have been released from their chorions and are undamaged (Figure 1C).
2.4. Drug Treatments
2.5. Imaging and Quantitation of Muscle Lesions
3. Results
3.1. Determination of Pronase Treatment Specifications
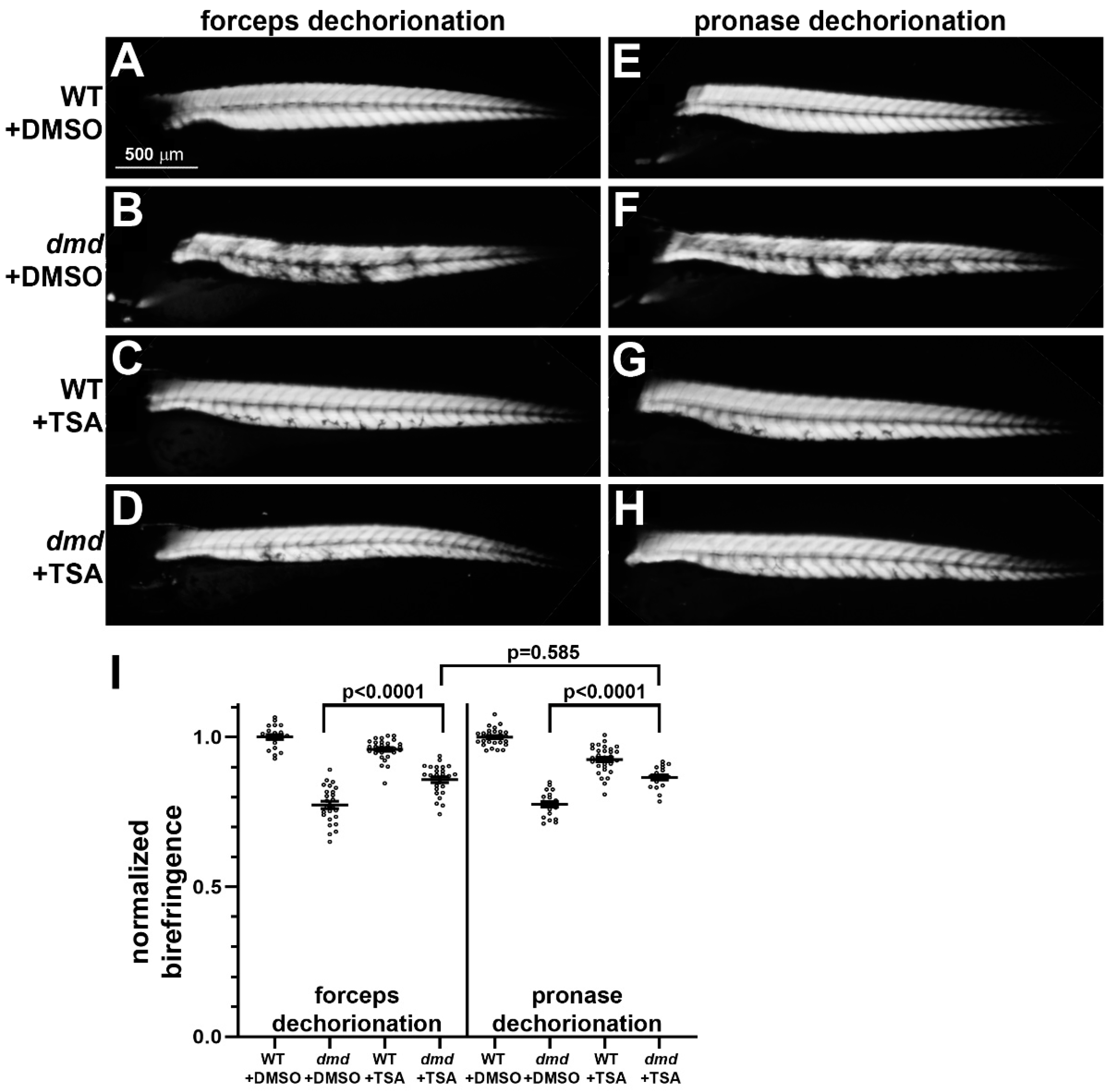
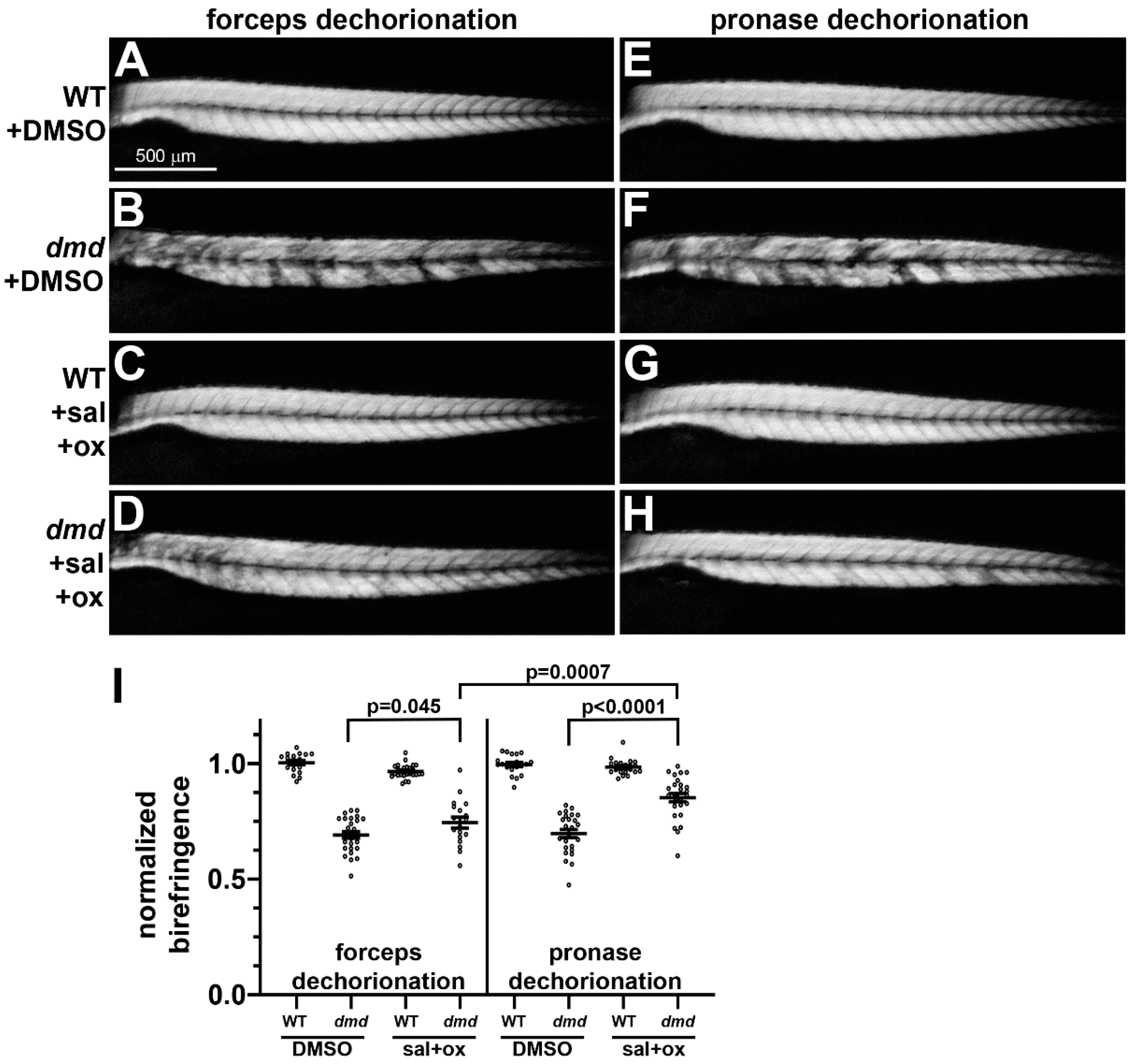
3.2. Small Molecule Treatments of dmd Embryos Exhibit Similar Rescue Following Either Manual or Pronase Dechorionation
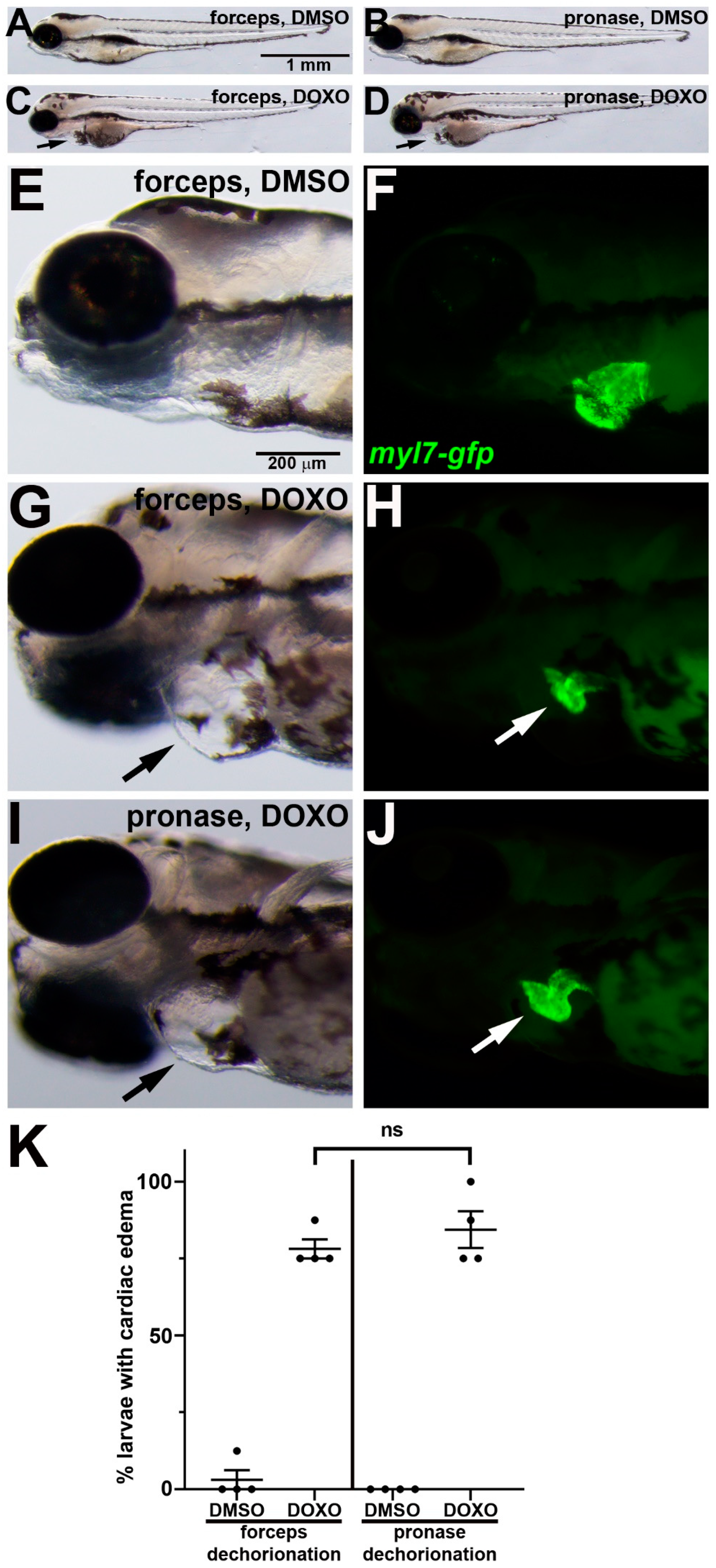
3.3. Doxorubicin Induces Cardiotoxicity Following Either Manual or Pronase Dechorionation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacRae, C.M.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Truong, L.; Bugel, S.M.; Chlebowski, A.; Usenko, C.Y.; Simonich, M.T.; Simonich, S.L.; Tanguay, R.L. Optimizing multi-dimensional high throughput screening using zebrafish. Reprod. Toxicol. 2016, 65, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horzmann, K.A.; Freeman, J.L. Making Waves: New Developments in Toxicology With the Zebrafish. Toxicol. Sci. 2018, 163, 5–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patton, E.E.; Zon, L.I.; Langenau, D.M. Zebrafish disease models in drug discovery: From preclinical modelling to clinical trials. Nat. Rev. Drug Discov. 2021, 20, 611–628. [Google Scholar] [CrossRef] [PubMed]
- Bonsignorio, D.; Perego, L.; Del Giacco, L.; Cotelli, F. Structure and macromolecular composition of the zebrafish egg chorion. Zygote 1996, 4, 101–108. [Google Scholar] [CrossRef]
- Iconomidou, V.A.; Chryssikos, D.G.; Gionis, V.; Pavlidis, M.A.; Paipetis, A.; Hamodrakas, S.J. Secondary structure of chorion proteins of the teleostean fish Dentex dentex by ATR FT-IR and FT-Raman spectroscopy. J. Struct. Biol. 2000, 132, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Bar-Ilan, O.; Louis, K.M.; Yang, S.P.; Pedersen, J.A.; Hamers, R.J.; Peterson, R.E.; Heideman, W. Titanium dioxide nanoparticles produce phototoxicity in the developing zebrafish. Nanotoxicology 2012, 6, 670–679. [Google Scholar] [CrossRef]
- Creton, R. The Calcium Pump of the Endoplasmic Reticulum Plays a Role in Midline Signaling during Early Zebrafish Development. Dev. Brain Res. 2004, 151, 33–41. [Google Scholar] [CrossRef]
- Henn, K.; Braunbeck, T. Dechorionation as a Tool to Improve the Fish Embryo Toxicity Test (FET) with the Zebrafish (Danio Rerio). Comp. Biochem. Physiol. Part-C: Toxicol. Pharmacol. 2011, 153, 91–98. [Google Scholar] [CrossRef]
- Kim, K.-T.; Tanguay, R.L. The role of chorion on toxicity of silver nanoparticles in the embryonic zebrafish assay. Environ. Anal. Health Toxicol. 2014, 29, e2014021. [Google Scholar] [CrossRef]
- Pelka, K.E.; Henn, K.; Keck, A.; Sapel, B.; Braunbeck, T. Size does matter-Determination of the critical molecular size for the uptake of chemicals across the chorion of zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2017, 185, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio), 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Kim, D.H.; Hwang, C.N.; Sun, Y.; Lee, S.H.; Kim, B.; Nelson, B.J. Mechanical Analysis of Chorion Softening in Prehatching Stages of Zebrafish Embryos. IEEE Trans. Nanobiosci. 2006, 5, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Mandrell, D.; Truong, L.; Jephson, C.; Sarker, M.R.; Moore, A.; Lang, C.; Simonich, M.T.; Tanguay, R.L. Automated Zebrafish Chorion Removal and Single Embryo Placement: Optimizing Throughput of Zebrafish Developmental Toxicity Screens. SLAS Technol. 2012, 17, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieger, S. Dechorionation of Zebrafish Embryos with Pronase for Metronidazole-Mediated β-Cell Ablation; The Diabetic Complications Consortium: Bethesda, MD, USA, 2013. [Google Scholar]
- Rho, S.S.; Oguri-Nakamura, E.; Ando, K.; Yamamoto, K.; Takagi, Y.; Fukuhara, S. Protocol for analysis of integrin-mediated cell adhesion of lateral plate mesoderm cells isolated from zebrafish embryos. STAR Protoc. 2021, 2, 100428. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.M.; Pohajdak, B.; Henry, M.; Wright, J.R., Jr. Structure and Enzymatic Removal of the Chorion of Embryos of the Nile Tilapia: Chorion of Oreochromis Niloticus. J. Fish Biol. 2003, 63, 1439–1453. [Google Scholar] [CrossRef]
- Martinez-Sales, M.; García- Ximénez, F.; Espinós, F.J. Zebrafish (Danio rerio) as a Possible Bioindicator of Epigenetic Factors Present in Drinking Water That May Affect Reproductive Function: Is Chorion an Issue? Zygote 2015, 23, 447–452. [Google Scholar] [CrossRef]
- Johnson, N.M.; Farr, G.H., III; Maves, L. The HDAC Inhibitor TSA Ameliorates a Zebrafish Model of Duchenne Muscular Dystrophy. PLoS Curr. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Bajanca, F.; Vandel, L. Epigenetic regulators modulate muscle damage in Duchenne muscular dystrophy model. PLoS Curr. 2017, 9. [Google Scholar] [CrossRef]
- Farr, G.H., III; Morris, M.; Gomez, A.; Pham, T.; Kilroy, E.; Parker, E.U.; Said, S.; Henry, C.; Maves, L. A novel chemical-combination screen in zebrafish identifies epigenetic small molecule candidates for the treatment of Duchenne muscular dystrophy. Skelet. Muscle. 2020, 10, 29. [Google Scholar] [CrossRef]
- Lane, S.; More, L.A.; Asnani, A. Zebrafish Models of Cancer Therapy-Induced Cardiovascular Toxicity. J. Cardiovasc. Dev. Dis. 2021, 8, 8. [Google Scholar] [CrossRef]
- Liu, Y.; Asnani, A.; Zou, L.; Bentley, V.L.; Yu, M.; Wang, Y.; Dellaire, G.; Sarkar, K.S.; Dai, M.; Chen, H.H.; et al. Visnagin protects against doxorubicin-induced cardiomyopathy through modulation of mitochondrial malate dehydrogenase. Sci. Transl. Med. 2014, 6, 266ra170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amgalan, D.; Garner, T.P.; Pekson, R.; Jia, X.F.; Yanamandala, M.; Paulino, V.; Liang, F.G.; Corbalan, J.J.; Lee, J.; Chen, Y.; et al. A small-molecule allosteric inhibitor of BAX protects against doxorubicin-induced cardiomyopathy. Nat. Cancer 2020, 1, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Maciag, M.; Wnorowski, A.; Mierzejewska, M.; Plazinska, A. Pharmacological assessment of zebrafish-based cardiotoxicity models. Biomed. Pharmacother. 2022, 148, 112695. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.I.; Bryson-Richardson, R.J.; Daggett, D.F.; Gautier, P.; Keenan, D.G.; Currie, P.D. Dystrophin is required for the formation of stable muscle attachments in the zebrafish embryo. Development 2003, 130, 5851–5860. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Van Eeden, F.J.; Schach, U.; Trowe, T.; Brand, M.; Furutani-Seiki, M.; Haffter, P.; Hammerschmidt, M.; Heisenberg, C.P.; Jiang, Y.J.; et al. Genes controlling and mediating locomotion behavior of the zebrafish embryo and larva. Development 1996, 123, 399–413. [Google Scholar] [CrossRef]
- Berger, J.; Sztal, T.; Currie, P.D. Quantification of Birefringence Readily Measures the Level of Muscle Damage in Zebrafish. Biochem. Biophys. Res. Commun. 2012, 423, 785–788. [Google Scholar] [CrossRef]
- Huang, C.J.; Tu, C.T.; Hsiao, C.D.; Hsieh, F.J.; Tsai, H.J. Germ-line transmission of a myocardium-specific GFP transgene reveals critical regulatory elements in the cardiac myosin light chain 2 promoter of zebrafish. Dev. Dyn. 2003, 228, 30–40. [Google Scholar] [CrossRef]
- Westhoff, J.H.; Steenbergen, P.J.; Thomas, L.S.V.; Heigwer, J.; Bruckner, T.; Cooper, L.; Tönshoff, B.; Hoffmann, G.F.; Gehrig, J. In Vivo High-Content Screening in Zebrafish for Developmental Nephrotoxicity of Approved Drugs. Front. Cell Dev. Biol. 2020, 8, 583. [Google Scholar] [CrossRef]
- Marguerie, A.; Gustafson, A.L.; Panter, G.; Stewart, J.; Hutchinson, T.; Alderton, W.; Oskarsson, A. A Pilot Study on the Effects of Mechanical Dechorionation on Developmental Toxicity in Zebrafish Embryos; 2007 HESI Workshop on Alternative Assay for Development: Paris, France, 2007. [Google Scholar]
- Craig, M.P.; Gilday, S.D.; Hove, J.R. Dose-dependent effects of chemical immobilization on the heart rate of embryonic zebrafish. Lab. Anim. 2006, 35, 41–47. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasegawa, E.H.; Farr, G.H., III; Maves, L. Comparison of Pronase versus Manual Dechorionation of Zebrafish Embryos for Small Molecule Treatments. J. Dev. Biol. 2023, 11, 16. https://doi.org/10.3390/jdb11020016
Hasegawa EH, Farr GH III, Maves L. Comparison of Pronase versus Manual Dechorionation of Zebrafish Embryos for Small Molecule Treatments. Journal of Developmental Biology. 2023; 11(2):16. https://doi.org/10.3390/jdb11020016
Chicago/Turabian StyleHasegawa, Eva H., Gist H. Farr, III, and Lisa Maves. 2023. "Comparison of Pronase versus Manual Dechorionation of Zebrafish Embryos for Small Molecule Treatments" Journal of Developmental Biology 11, no. 2: 16. https://doi.org/10.3390/jdb11020016