
The Influence of Lipid Electric Charge on the Binding of Aβ(1–42) Amyloid Peptide to Bilayers in the Liquid-Ordered State

Abstract
:1. Introduction
2. Results
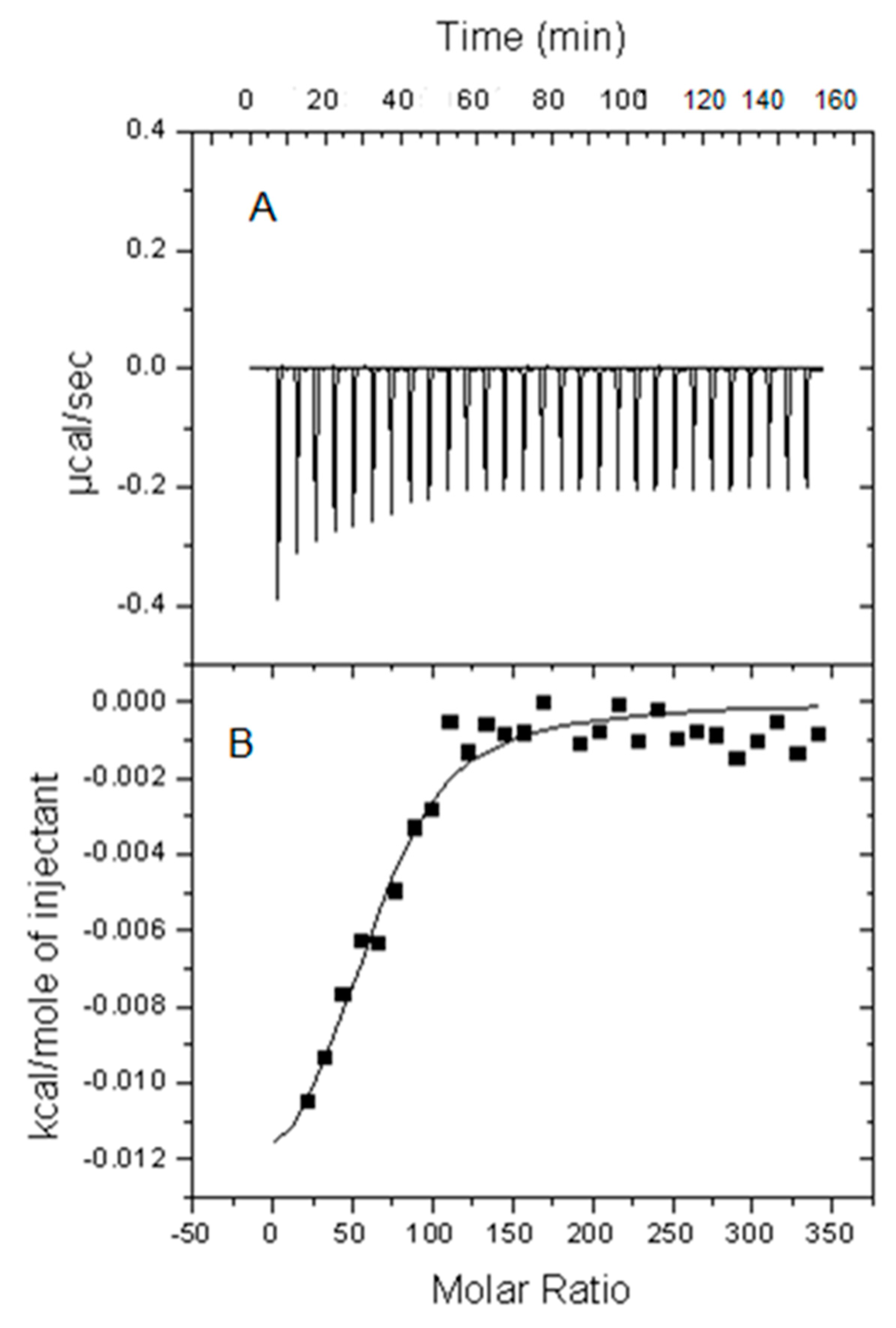
2.1. Calorimetric Studies
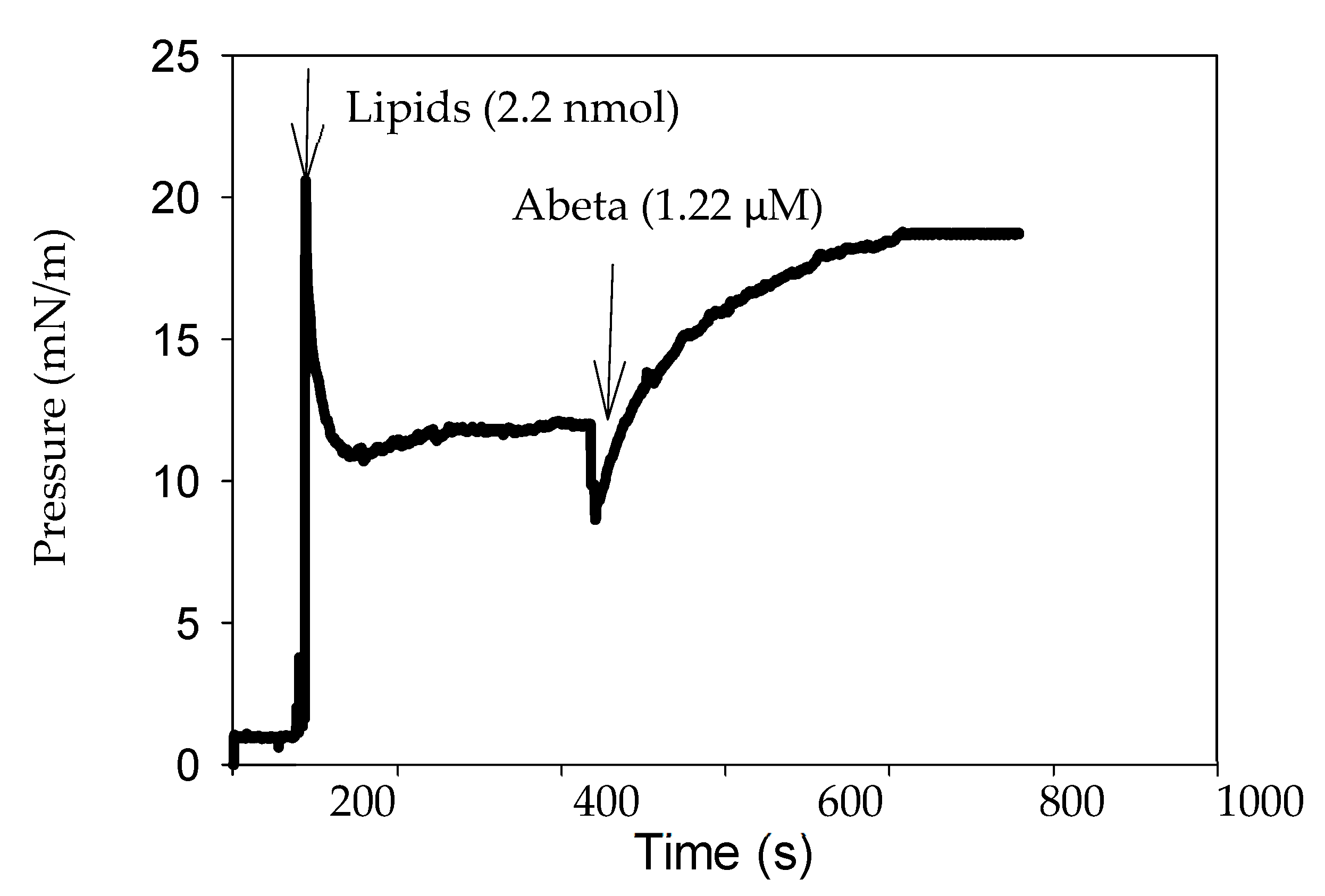
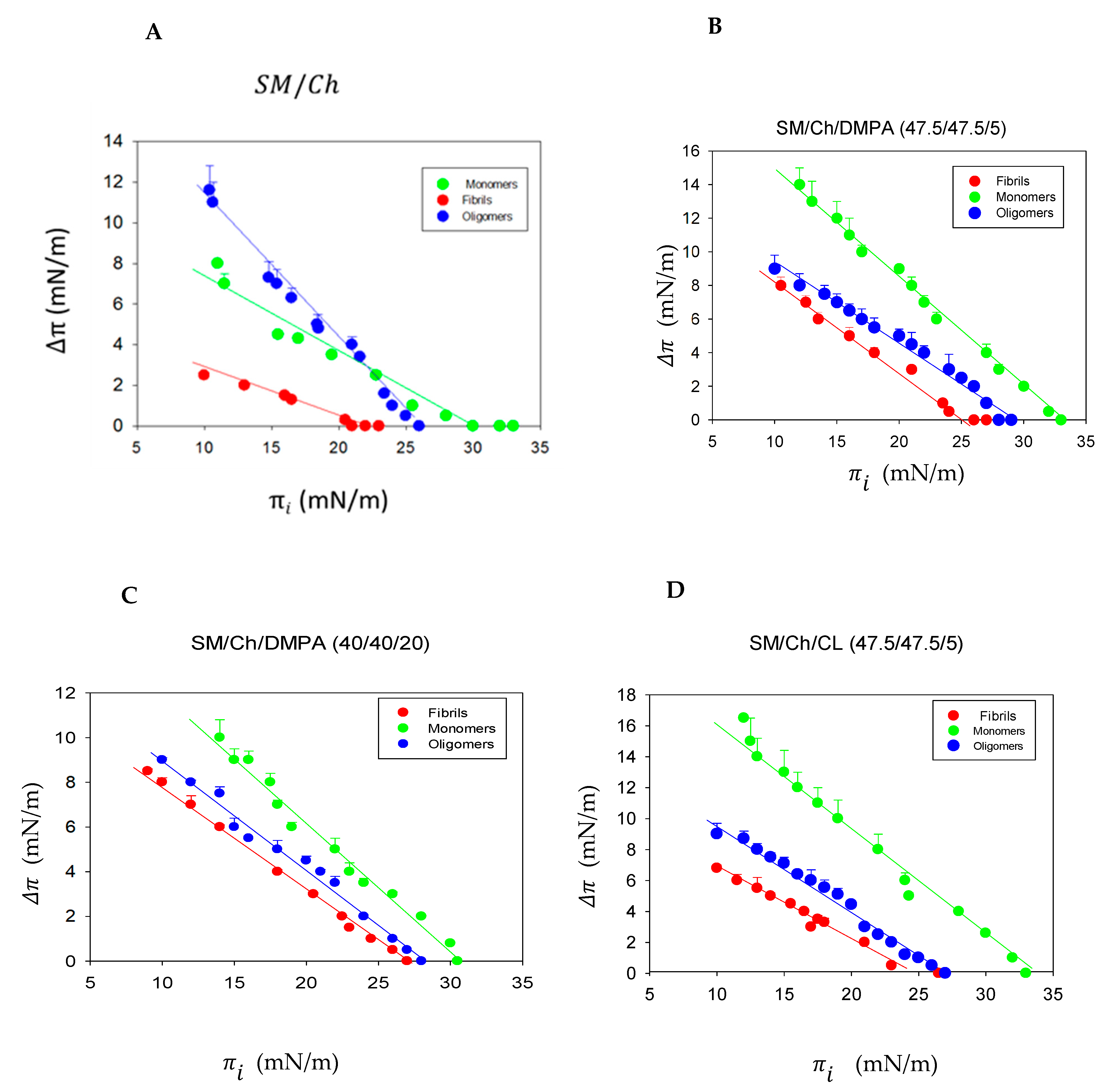
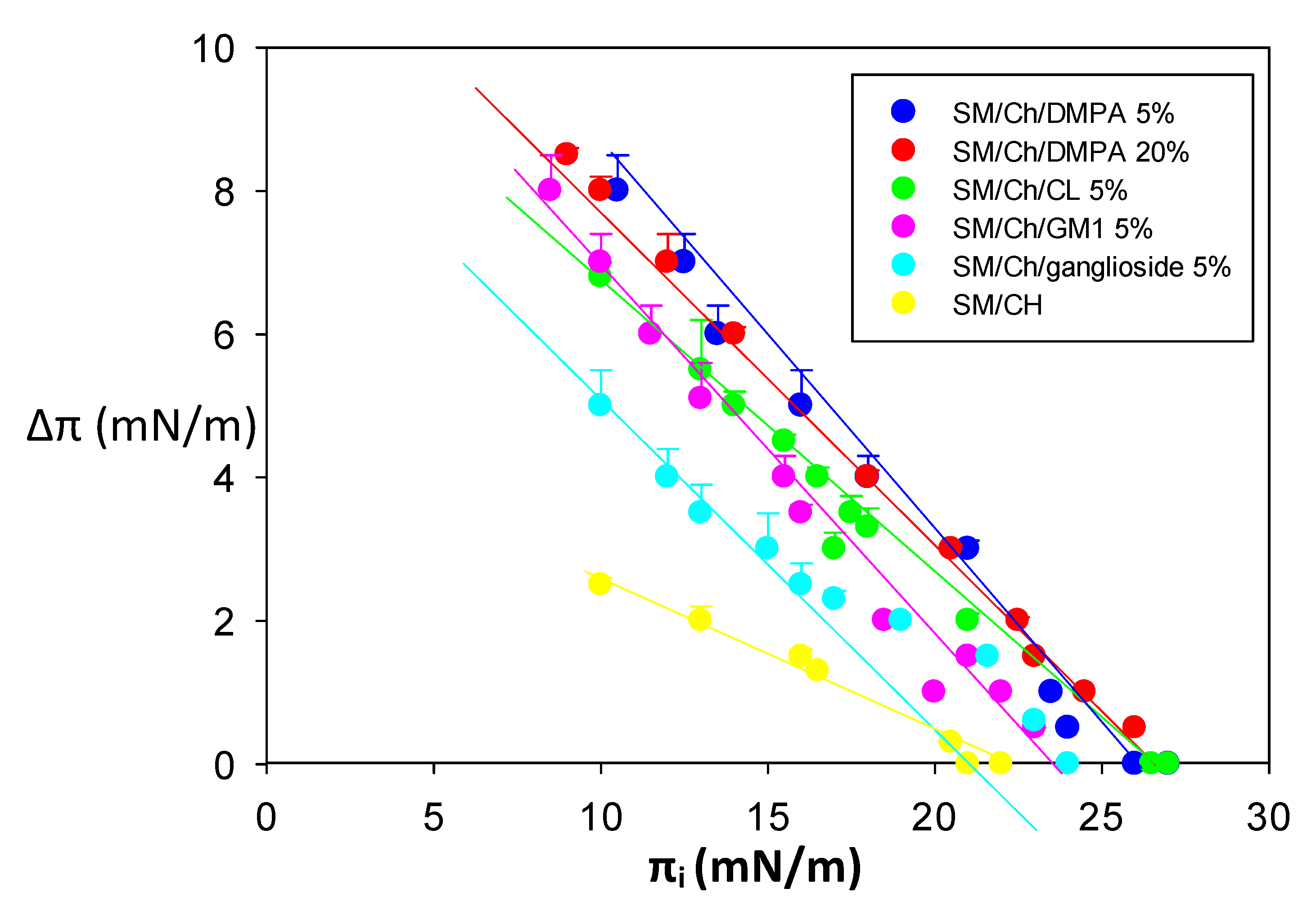
2.2. Monolayer Lateral Pressure Studies
3. Discussion
3.1. Two Techniques, One Phenomenon?
3.2. The Peptide Aggregation State
3.3. The Bilayer Physical Properties
3.4. From Model to Cell Membranes
4. Materials and Methods
4.1. Materials
4.2. Aβ(1-42) Sample Preparation
4.3. Monomers
4.4. Oligomers
4.5. Fibrils
4.6. Liposome Preparation
4.7. Isothermal Titration Calorimetry
4.8. Lipid Monolayer Measurements
5. Concluding Remarks
- (a)
- Interaction of the amyloidogenic Aβ(1-42) peptide with cell membranes can be mimicked using model lipid monolayers or bilayers. Interaction with bilayer membranes can adopt at least two different forms: adsorption onto the membrane surface or insertion into it. The expression ‘membrane binding’ is often used to encompass both situations. Adsorption and insertion should not be considered as two different end-points of a process: adsorption can be a reversible, intermediate step leading to either insertion or desorption, while insertion is usually an irreversible event. Of the two main techniques used in the present study to measure lipid–protein interaction, increased surface pressures, as detected in the Langmuir balance, are usually interpreted in terms of peptide insertion into the monolayer. However, insertion into a monolayer is not equivalent to insertion in a bilayer; the former can occur without the latter. The calorimetric assessment of the interaction does not allow, in turn, to distinguish between adsorption and insertion; thus, our observations are globally referred to as peptide binding.
- (b)
- It is generally accepted that Aβ monomers associate among themselves, ultimately giving rise to micrometer-sized amyloid plaques, monomers giving rise to oligomers, then to fibrils. These early aggregation steps can occur in aqueous media, although they might be facilitated/catalyzed by a primer consisting of a membrane-bound peptide molecule. Some of the above experimental results appear to indicate that preparations highly enriched in either monomers, oligomers, or fibrils interact differentially with membranes; this does not exclude that multiple equilibria (monomers, oligomers, fibrils) are simultaneously occurring. Thus, any quantitative analysis of Aβ amyloid formation in membranes must take into account these complex inter-peptide and peptide–lipid interactions.
- (c)
- Model membrane bilayers can be prepared, among others, in the liquid-disordered and the liquid-ordered states that could be respectively represented, e.g., by the lamellar phases of egg phosphatidylcholine and of an equimolar sphingomyelin/cholesterol mixture. However, these are extreme examples that may or may not correlate with cell membranes. A realistic interpretation of results obtained with those kinds of compositions should keep in mind that natural membranes are not laterally homogeneous so that domains with different degrees of molecular order can coexist. Moreover, within a given domain, a gradual spectrum of molecular order may occur between the fully ordered and the fully disordered states. Peptides and proteins tend to insert more easily into more disordered domains/membranes or at the interfaces between ordered and disordered domains.
- (d)
- The above data with monolayers and bilayers based on equimolar sphingomyelin/cholesterol mixtures correspond to a hypothetical cell membrane situation in which Aβ(1-42) binding is very difficult. They would represent a basal or minimal binding that would increase in the presence of negatively charged lipids (at concentrations compatible with the liquid-ordered state), particularly for monomers.
- (e)
- A reasonable hypothetical scenario would contemplate that, even in highly ordered domains, Aβ(1-42) monomers would be able to bind in the presence of some negatively charged lipids, in turn interacting with basic amino acid residues in Aβ(1-42), e.g., Lys-28. The membrane-bound monomer would then act as a catalyst (or a primer) for β-sheet formation, oligomerization, fibril formation, and ultimately, plaque deposit.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lage, J.M. 100 Years of Alzheimer’s disease (1906–2006). J. Alzheimers Dis. 2006, 9 (Suppl. S3), 15–26. [Google Scholar] [CrossRef] [PubMed]
- Glenner, G.G.; Wong, C.W. Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984, 120, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, A.; Debbabi, M.; Bezine, M.; Karym, E.M.; Badreddine, A.; Rouaud, O.; Moreau, T.; Cherkaoui-Malki, M.; El Ayeb, M.; Nasser, B.; et al. Lipid Biomarkers in Alzheimer’s Disease. Curr. Alzheimer Res. 2018, 15, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Campos-Peña, V.; Pichardo-Rojas, P.; Sánchez-Barbosa, T.; Ortíz-Islas, E.; Rodríguez-Pérez, C.E.; Montes, P.; Ramos-Palacios, G.; Silva-Adaya, D.; Valencia-Quintana, R.; Cerna-Cortes, J.F.; et al. Amyloid β, Lipid Metabolism, Basal Cholinergic System, and Therapeutics in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 12092. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.B.; Rodríguez, P.E.A.; Fidelio, G.D. Interfacial Aβ fibril formation is modulated by the disorder-order state of the lipids: The concept of the physical environment as amyloid inductor in biomembranes. Biochim. Biophys. Acta Biomembr. 2023, 1866, 184234. [Google Scholar] [CrossRef] [PubMed]
- Krasnobaev, V.D.; Bershatsky, Y.V.; Bocharova, O.V.; Bocharov, E.V.; Batishchev, O.V. Amyloid Precursor Protein Changes Arrangement in a Membrane and Its Structure Depending on the Cholesterol Content. Membranes 2023, 13, 706. [Google Scholar] [CrossRef]
- Saha, J.; Bose, P.; Dhakal, S.; Ghosh, P.; Rangachari, V. Ganglioside-Enriched Phospholipid Vesicles Induce Cooperative Aβ Oligomerization and Membrane Disruption. Biochemistry 2022, 61, 2206–2220. [Google Scholar] [CrossRef]
- Kumar, M.; Ivanova, M.I.; Ramamoorthy, A. Non-micellar ganglioside GM1 induces an instantaneous conformational change in Aβ42 leading to the modulation of the peptide amyloid-fibril pathway. Biophys. Chem. 2023, 301, 107091. [Google Scholar] [CrossRef]
- Kumar, M.; Ivanova, M.I.; Ramamoorthy, A. Ganglioside GM1 produces stable, short, and cytotoxic Aβ40 protofibrils. Chem. Commun. 2023, 59, 7040–7043. [Google Scholar] [CrossRef]
- Yagi-Utsumi, M.; Itoh, S.G.; Okumura, H.; Yanagisawa, K.; Kato, K.; Nishimura, K. The Double-Layered Structure of Amyloid-β Assemblage on GM1-Containing Membranes Catalytically Promotes Fibrillization. ACS Chem. Neurosci. 2023, 14, 2648–2657. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.T.; Tang, T.Y.C.; Kuo, C.T.; Yu, Y.T.; Chen, E.H.L.; Lee, M.T.; Tsai, R.F.; Chen, H.Y.; Chiang, Y.W.; Chen, R.P.Y. Cholesterol twists the transmembrane Di-Gly region of amyloid-precursor protein. PNAS Nexus 2023, 2, pgad162. [Google Scholar] [CrossRef] [PubMed]
- Gamba, P.; Testa, G.; Gargiulo, S.; Staurenghi, E.; Poli, G.; Leonarduzzi, G. Oxidized cholesterol as the driving force behind the development of Alzheimer’s disease. Front. Aging Neurosci. 2015, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Iriondo, A.; García-Sebastian, M.; Arrospide, A.; Arriba, M.; Aurtenetxe, S.; Barandiaran, M.; Clerigue, M.; Ecay-Torres, M.; Estanga, A.; Gabilondo, A.; et al. Cerebrospinal Fluid 7-Ketocholesterol Level is Associated with Amyloid-β42 and White Matter Microstructure in Cognitively Healthy Adults. J. Alzheimers Dis. 2020, 76, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Ahyayauch, H.; Raab, M.; Busto, J.V.; Andraka, N.; Arrondo, J.R.; Masserini, M.; Tvaroska, I.; Goñi, F.M. Binding of β-amyloid (1–42) peptide to negatively charged phospholipid membranes in the liquid-ordered state: Modeling and experimental studies. Biophys. J. 2012, 103, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Ahyayauch, H.; García-Arribas, A.B.; Masserini, M.E.; Pantano, S.; Goñi, F.M.; Alonso, A. β-Amyloid (1–42) peptide adsorbs but does not insert into ganglioside-containing phospholipid membranes in the liquid-disordered state: Modelling and experimental studies. Int. J. Biol. Macromol. 2020, 164, 2651–2658. [Google Scholar] [CrossRef] [PubMed]
- Ahyayauch, H.; de la Arada, I.; Masserini, M.E.; Arrondo, J.L.R.; Goñi, F.M.; Alonso, A. The Binding of Aβ42 Peptide Monomers to Sphingomyelin/Cholesterol/Ganglioside Bilayers Assayed by Density Gradient Ultracentrifugation. Int. J. Mol. Sci. 2020, 21, 1674. [Google Scholar] [CrossRef] [PubMed]
- Ahyayauch, H.; Masserini, M.; Goñi, F.M.; Alonso, A. The interaction of Aβ42 peptide in monomer, oligomer or fibril forms with sphingomyelin/cholesterol/ganglioside bilayers. Int. J. Biol. Macromol. 2021, 168, 611–619. [Google Scholar] [CrossRef]
- Davis, C.H.; Berkowitz, M.L. Interaction between amyloid-beta (1–42) peptide and phospholipid bilayers: A molecular dynamics study. Biophys. J. 2009, 96, 785–797. [Google Scholar] [CrossRef]
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-β Oligomer Hypothesis: Beginning of the Third Decade. J. Alzheimers Dis. 2018, 64, S567–S610. [Google Scholar] [CrossRef]
- Maget-Dana, R. The monolayer technique: A potent tool for studying the interfacial properties of antimicrobial and membrane-lytic peptides and their interactions with lipid membranes. Biochim. Biophys. Acta 1999, 1462, 109–140. [Google Scholar] [CrossRef] [PubMed]
- Ambroggio, E.E.; Kim, D.H.; Separovic, F.; Barrow, C.J.; Barnham, K.J.; Bagatolli, L.A.; Fidelio, G.D. Surface behavior and lipid interaction of Alzheimer beta-amyloid peptide 1–42: A membrane-disrupting peptide. Biophys. J. 2005, 88, 2706–2713. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D. Lateral pressure in membranes. Biochim. Biophys. Acta 1996, 1286, 183–223. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, A.; McConnell, H.M. Critical points in charged membranes containing cholesterol. Proc. Natl. Acad. Sci. USA 2002, 99, 13391–13396. [Google Scholar] [CrossRef] [PubMed]
- Heerklotz, H.; Seelig, J. Titration calorimetry of surfactant-membrane partitioning and membrane solubilization. Biochim. Biophys. Acta 2000, 1508, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, G.; Prabhu, R.; Freire, E. Direct measurement of the energetics of association between myelin basic protein and phosphatidylserine vesicles. Biochemistry 1986, 25, 2265–2270. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Magraner, L.; Cortajarena, A.L.; Goñi, F.M.; Ostolaza, H. Membrane insertion of Escherichia coli alpha-hemolysin is independent from membrane lysis. J. Biol. Chem. 2006, 281, 5461–5467. [Google Scholar] [CrossRef]
- Yin, Y.I.; Bassit, B.; Zhu, L.; Yang, X.; Wang, C.; Li, Y.M.J. {gamma}-Secretase substrate concentration modulates the Abeta42/Abeta40 ratio: Implications for Alzheimer disease. Biol. Chem. 2007, 282, 23639–23644. [Google Scholar] [CrossRef]
- Goñi, F.M.; Alonso, A.; Bagatolli, L.A.; Brown, R.E.; Marsh, D.; Prieto, M.; Thewalt, J.L. Phase diagrams of lipid mixtures relevant to the study of membrane rafts. Biochim. Biophys. Acta 2008, 1781, 665–684. [Google Scholar] [CrossRef]
- Camino, J.D.; Gracia, P.; Chen, S.W.; Sot, J.; de la Arada, I.; Sebastián, V.; Arrondo, J.L.R.; Goñi, F.M.; Dobson, C.M.; Cremades, N. The extent of protein hydration dictates the preference for heterogeneous or homogeneous nucleation generating either parallel or antiparallel β-sheet α-synuclein aggregates. Chem. Sci. 2020, 11, 11902–11914. [Google Scholar] [CrossRef]
- Goñi, F.M. “Rafts”: A nickname for putative transient nanodomains. Chem. Phys. Lipids 2019, 218, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, S.; Grzybek, M.; Orłowski, A.; Róg, T.; Cramariuc, O.; Levental, I.; Eggeling, C.; Sezgin, E.; Vattulainen, I. Phase Partitioning of GM1 and Its Bodipy-Labeled Analog Determine Their Different Binding to Cholera Toxin. Front. Physiol. 2017, 8, 252. [Google Scholar] [CrossRef]
- Tsai, W.C.; Feigenson, G.W. Lowering line tension with high cholesterol content induces a transition from macroscopic to nanoscopic phase domains in model biomembranes. Biochim. Biophys. Acta Biomembr. 2019, 1861, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Ehehalt, R.; Keller, P.; Haass, C.; Thiele, C.; Simons, K. Amyloidogenic processing of the Alzheimer beta-amyloid precursor protein depends on lipid rafts. J. Cell Biol. 2003, 160, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Park, I.H.; Hwang, E.M.; Hong, H.S.; Boo, J.H.; Oh, S.S.; Lee, J.; Jung, M.W.; Bang, O.Y.; Kim, S.U.; Mook-Jung, I. Lovastatin enhances Abeta production and senile plaque deposition in female Tg2576 mice. Neurobiol. Aging. 2003, 24, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Abad-Rodriguez, J.; Ledesma, M.D.; Craessaerts, K.; Perga, S.; Medina, M.; Delacourte, A.; Dingwall, C.; De Strooper, B.; Dotti, C.G. Neuronal membrane cholesterol loss enhances amyloid peptide generation. J. Cell Biol. 2004, 167, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Siniscalco, D.; Francius, G.; Tarek, M.; Bali, S.K.; Laprévote, O.; Malaplate, C.; Oster, T.; Pauron, L.; Quilès, F. Molecular Insights for Alzheimer’s Disease: An Unexplored Storyline on the Nanoscale Impact of Nascent Aβ1–42 toward the Lipid Membrane. ACS Appl. Mater. Interfaces 2023, 15, 17507–17517. [Google Scholar] [CrossRef]
- Ipsen, J.H.; Karlström, G.; Mouritsen, O.G.; Wennerström, H.; Zuckermann, M.J. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim. Biophys. Acta 1987, 905, 162–172. [Google Scholar] [CrossRef]
- Chauhan, A.; Ray, I.; Chauhan, V.P. Interaction of amyloid beta-protein with anionic phospholipids: Possible involvement of Lys28 and C-terminus aliphatic amino acids. Neurochem. Res. 2000, 25, 423–429. [Google Scholar] [CrossRef]
- Simons, K.; Ehehalt, R. Cholesterol, lipid rafts, and disease. J. Clin. Investig. 2002, 110, 597–603. [Google Scholar] [CrossRef]
- Robinson, J.; Sarangi, N.K.; Keyes, T.E. Role of phosphatidylserine in amyloid-beta oligomerization at asymmetric phospholipid bilayers. Phys. Chem. Chem. Phys. 2023, 25, 7648–7661. [Google Scholar] [CrossRef] [PubMed]
- Saha, J.; Ford, B.J.; Wang, X.; Boyd, S.; Morgan, S.E.; Rangachari, V. Sugar distributions on gangliosides guide the formation and stability of amyloid-β oligomers. Biophys. Chem. 2023, 300, 107073. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.B.; Rodríguez, P.E.A.; Fidelio, G.D. Gangliosides smelt nanostructured amyloid Aβ(1-40) fibrils in a membrane lipid environment. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183749. [Google Scholar] [CrossRef] [PubMed]
- Haass, C.; Selkoe, D.J. Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer’s amyloid beta-peptide. Nat. Rev. Mol. Cell Biol. 2007, 8, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Levental, I.; Lyman, E. Regulation of membrane protein structure and function by their lipid nano-environment. Nat. Rev. Mol. Cell Biol. 2023, 24, 107–122. [Google Scholar] [CrossRef] [PubMed]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- de la Cueva, M.; Antequera, D.; Ordoñez-Gutierrez, L.; Wandosell, F.; Camins, A.; Carro, E.; Bartolome, F. Amyloid-β impairs mitochondrial dynamics and autophagy in Alzheimer’s disease experimental models. Sci. Rep. 2022, 12, 10092. [Google Scholar] [CrossRef]
- Hoshino, F.; Sakane, F. Docosahexaenoic acid-containing phosphatidic acid interacts with clathrin coat assembly protein AP180 and regulates its interaction with clathrin. Biochem. Biophys. Res. Commun. 2022, 587, 69–77. [Google Scholar] [CrossRef]
- Burrinha, T.; Martinsson, I.; Gomes, R.; Terrasso, A.P.; Gouras, G.K.; Almeida, C.G. Upregulation of APP endocytosis by neuronal aging drives amyloid-dependent synapse loss. J. Cell Sci. 2021, 134, jcs255752. [Google Scholar] [CrossRef]
- Chigorno, V.; Sonnino, S.; Ghidoni, R.; Tettamanti, G. Densitometric quantification of brain gangliosides separated by two-dimensional thin layer chromatography. Neurochem. Int. 1982, 4, 397–404. [Google Scholar] [CrossRef]
- Gobbi, M.; Re, F.; Canovi, M.; Beeg, M.; Gregori, M.; Sesana, S.; Sonnino, S.; Brogioli, D.; Musicanti, C.; Gasco, P.; et al. Lipid-based nanoparticles with high binding affinity for amyloid-beta1-42 peptide. Biomaterials 2010, 31, 6519–6529. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.R. Techniques to study amyloid fibril formation in vitro. Methods 2004, 34, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, K.N.; Manelli, A.M.; Stine, W.B., Jr.; Baker, L.K.; Krafft, G.A.; LaDu, M.J. Oligomeric and fibrillar species of amyloid-beta peptides differentially affect neuronal viability. J. Biol. Chem. 2002, 277, 32046–32053. [Google Scholar] [CrossRef] [PubMed]
- Gregori, M.; Cassina, V.; Brogioli, D.; Salerno, D.; De Kimpe, L.; Scheper, W.; Masserini, M.; Mantegazza, F. Stability of Aβ (1–42) peptide fibrils as consequence of environmental modifications. Eur. Biophys. J. 2010, 39, 1613–1623. [Google Scholar] [CrossRef]
- Arnulphi, C.; Sot, J.; García-Pacios, M.; Arrondo, J.L.; Alonso, A.; Goñi, F.M. Triton X-100 partitioning into sphingomyelin bilayers at subsolubilizing detergent concentrations: Effect of lipid phase and a comparison with dipalmitoylphosphatidylcholine. Biophys. J. 2007, 93, 3504–3514. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Heerklotz, H.; Jahnke, N.; Blume, A. Thermodynamics of lipid membrane solubilization by sodium dodecyl sulfate. Biophys. J. 2006, 90, 4509–4521. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Magraner, L.; Viguera, A.R.; García-Pacios, M.; Garcillán, M.P.; Arrondo, J.L.; de la Cruz, F.; Goñi, F.M.; Ostolaza, H. The calcium-binding C-terminal domain of Escherichia coli alpha-hemolysin is a major determinant in the surface-active properties of the protein. J. Biol. Chem. 2007, 282, 11827–11835. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monomer | SM/Ch/DMPA (47.5/47.5/5) a | SM/Ch/DMPA (40/40/20) a | SM/Ch/CL (47.5/47.5/5) a | SM/Ch/GM1 (47.5/47.5/5) b | SM/Ch/T. Gang. (47.5/47.5/5) b | SM/Ch (1/1) |
|---|---|---|---|---|---|---|
| Ka (M−1) (×104) | 3.09 ± 0.9 | 58.2 ± 8.0 | 16.0 ± 2 | 9.4 ± 0.3 | 28.0 ± 2 | - |
| Kd (μM) | 32 ± 1.1 | 1.71 ± 12.5 | 6.25 ± 1.2 | 10.6 ± 0.3 | 3.5 ± 0.2 | - |
| ∆H (kcal/mol) | −7.3 ± 0.05 | −2.8 ± 0.19 | −12.6 ± 1.4 | −15.6 ± 1.4 | −108.2 ± 12 | - |
| ∆S (cal/mol K) | −3.0 ± 0.1 | 8.1 ± 0.2 | −1.68 ± 0.4 | −27.4 ± 1.8 | −324 ± 8 | - |
| ∆G (kcal/mol) | −6.4 ± 0.1 | −5.3 ± 0.01 | −11.96 ± 0.6 | −7.1 ± 0.6 | −7.8 ± 0.6 | - |
| Oligomer | SM/Ch/DMPA (47.5/47.5/5) | SM/Ch/DMPA (40/40/20) | SM/Ch/CL (47.5/47.5/5) | SM/Ch/GM1 (47.5/47.5/5) a | SM/Ch/T. Gang. (47.5/47.5/5) a | SM/Ch (1/1) a |
|---|---|---|---|---|---|---|
| Ka (M−1) (×104) | 1.22 ± 0.23 | 16.4 ± 1.3 | 51 ± 1 | 11 ± 0.1 | 21.6 ± 0.8 | 36.7 ± 0.7 |
| Kd (µM) | 81.9 ± 4.3 | 6.1 ± 0.77 | 2.0 ± 9.09 | 9.1 ± 0.91 | 4.6 ± 0.1 | 2.72 ± 0.01 |
| ∆H (kcal/mol) | −1.19 ± 0.15 | −4.27 ± 0.53 | −4.92 ± 0.43 | −0.83 ± 0.02 | −2.43 ± 0.05 | −2.11 ± 0.01 |
| ∆S(cal/mol K) | 14.9 ± 1.5 | 10.1 ± 1.2 | 1.11 ± 0.52 | 11.2 ± 0.5 | 16.6 ± 0.4 | 18.6 ± 0.1 |
| ∆G (kcal/mol) | −5.80 ± 0.34 | −7.37 ± 0.25 | −5.25 ± 0.34 | −4.25 ± 0.07 | −7.57 ± 0.09 | −7.88 ± 0.04 |
| Fibril | SM/Ch/DMPA (47.5/47.5/5) | SM/Ch/DMPA (40/40/20) | SM/Ch/CL (47.5/47.5/5) | SM/Ch/GM1 (47.5/47.5/5) a | SM/Ch/T. Gang. (47.5/47.5/5) a | SM/Ch (1/1) a |
|---|---|---|---|---|---|---|
| Ka (M−1)(×104) | 18.5 ± 2.4 | 27.6 ± 8.2 | 28.6 ± 0.8 | 18.0 ± 0.7 | 13.0 ± 0.3 | 21.0 ± 0.4 |
| Kd (µM) | 5.4 ± 0.18 | 3.6 ± 0.12 | 3.5 ± 0.3 | 5.6 ± 0.2 | 7.6 ± 0.3 | 4.8 ± 0.7 |
| ∆H (kcal/mol) | −6.14 ± 0.3 | −8.41 ± 0.405 | −4.31 ± 0.07 | −1.78 ± 0.05 | −29.6 ± 0.4 | −0.87 ± 0.05 |
| ∆S (cal/mol K) | 4.31 ± 0.08 | −2.22 ± 0.05 | 11.1 ± 0.2 | 18.3 ± 0.7 | −71.9 ± 1.8 | 21.6 ± 0.2 |
| ∆G (kcal/mol) | −7.47 ± 0.02 | −7.72 ± 0.03 | −7.75 ± 0.06 | −7.45 ± 0.08 | −7.27 ± 0.05 | −7.56 ± 0.04 |
| Lipid Composition | ∆π (mN/m) at πi = 16 mN/m | Maximal Insertionpressures | |
|---|---|---|---|
| SM/Ch a | Monomers | 4.5 ± 0.3 | 32 ± 0.5 |
| Oligomers | 6.0 ± 0.2 | 26 ± 0.0 | |
| Fibrils | 1.94 ± 0.2 | 21 ± 0.4 | |
| SM/Ch/DMPA (5%) | Monomers | 11.0 ± 0.6 | 32 ± 0.0 |
| Oligomers | 6.5 ± 0.7 | 28 ± 0.2 | |
| Fibrils | 5.0 ± 0.4 | 26 ± 0.3 | |
| SM/Ch/DMPA (20%) | Monomers | 9.2 ± 0.5 | 30 ± 0.1 |
| Oligomers | 5.5 ± 0.1 | 27 ± 0.2 | |
| Fibrils | 5.3 ± 0.2 | 26 ± 0.4 | |
| SM/Ch/CL (5%) | Monomers | 12.0 ± 1 | 32 ± 0.0 |
| Oligomers | 6.4 ± 0.2 | 27 ± 0.1 | |
| Fibrils | 4.2 ± 0.1 | 26 ± 0.2 | |
| SM/Ch/GM1 (5%) a | Monomers | 17.1 ± 0.4 | 34 ± 0.2 |
| Oligomers | 12.4 ± 0.5 | 32 ± 0.0 | |
| Fibrils | 3.5 ± 0.5 | 24 ± 0.5 | |
| SM/Ch/total ganglioside (5%) a | Monomers | 11.7 ± 0.5 | 34 ± 0.0 |
| Oligomers | 8.5 ± 0.6 | 31 ± 0.3 | |
| Fibrils | 2.7 ± 0.2 | 24 ± 0.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahyayauch, H.; Masserini, M.E.; Goñi, F.M.; Alonso, A. The Influence of Lipid Electric Charge on the Binding of Aβ(1–42) Amyloid Peptide to Bilayers in the Liquid-Ordered State. Biomolecules 2024, 14, 298. https://doi.org/10.3390/biom14030298
Ahyayauch H, Masserini ME, Goñi FM, Alonso A. The Influence of Lipid Electric Charge on the Binding of Aβ(1–42) Amyloid Peptide to Bilayers in the Liquid-Ordered State. Biomolecules. 2024; 14(3):298. https://doi.org/10.3390/biom14030298
Chicago/Turabian StyleAhyayauch, Hasna, Massimo E. Masserini, Félix M. Goñi, and Alicia Alonso. 2024. "The Influence of Lipid Electric Charge on the Binding of Aβ(1–42) Amyloid Peptide to Bilayers in the Liquid-Ordered State" Biomolecules 14, no. 3: 298. https://doi.org/10.3390/biom14030298